Abstract
The enzyme β-galactosidase catalyzes the hydrolysis of lactose into glucose and galactose, although for its effective application it is necessary to establish techniques for purification, concentration, or polishing, such as membrane separation processes, in particular ultrafiltration. The present study aimed to investigate ultrafiltration and diafiltration applied as initial steps for concentration and salt removal, respectively, in the β-galactosidase purification processes. Additionally, the influence levels of the pH (6.5, 7.7, or 7.5), membrane molecular weight cut-off (30, 50, 60, or 100 kDa), operating pressure (1.5, 2.0, or 2.5 kgf/cm2), and ionic strength of the ultrafiltration using NaCL or KCl (0.01–0.1 M) were evaluated considering the enzyme recovery, purification, retention, and concentration factors in relation to the proteins, volume, activity, and protein flux and yield of the processes. The ultrafiltration of the crude enzyme extract at pH 7.5 and 1.5 kgf/cm2 with a 50 kDa polyethersulfone membrane resulted in a volume concentration of the β-galactosidase extract up to 7.1-fold greater, a purification factor 1.2-fold greater, and an enzyme recovery rate of 108.9% by eliminating metabolites during the purification process. In addition, the lowest flux variation range (16.0 to 13.1 L/m2·h) was observed under these same conditions, thereby representing a decrease of 18.0%. An increase in the operating pressure and the addition of salts results in reduced enzyme recovery (up to 38% of the process yield (734.1 to 453.7 U/h) and up to 40% of the enzyme recovery rate (108.9 to 60.6%) during the ultrafiltration using NaCl, respectively). The operation in the diafiltration mode allowed salt removal after the purification of β-galactosidase (enzymatic recovery rates above 93.4%) via precipitation and ion-exchange chromatography elution and as part of an aqueous two-phase system using 6 diafiltration cycles, thereby revealing its application potential.
1. Introduction
The enzyme β-galactosidase (E.C 3.2.1.23) catalyzes the hydrolysis of the disaccharide lactose into the monosaccharides glucose and galactose. The enzyme may act in solutions and also in different matrices rich in lactose, being applied in different stages of the processes as a fundamental way to reduce the lactose content of dairy products intended for people with lactose intolerance [1,2]. Around 70% of the adult population worldwide is lactose-intolerant, reaching 95–100% in Asians, for example. This makes it necessary for the food industry to apply technologies to reduce the lactose contents in products intended for these lactose-intolerant people in order to avoid symptoms such as diarrhea, flatulence, and abdominal cramps that appear with the consumption of lactose [3,4]. Moreover, hydrolysis with β-galactosidase contributes to the formation of galactooligosaccharides (GOS) [5,6,7], which are beneficial to humans [8] and improve the technological and sensory characteristics of foods [9,10,11]. However, for effective hydrolysis using enzymes, recovery techniques that do not compromise the enzymatic activity, and consequently the applications, viability, and economic potential, need to be applied [1].
Ultrafiltration (UF) is a technique based on the macromolecule size-exclusion mechanism and can be applied to both the concentration and polishing of biological products [12]. The concentration and separation of the biological material using membranes does not require additives or phase changes and can be performed isothermally at low temperatures and pressures, with lower energy consumption compared with the thermal processes. Furthermore, these conditions ensure minimal denaturation of bioproducts that are highly unstable and are less expensive than other methods [12,13,14]. In ultrafiltration systems, the operating conditions strongly affect the concentration process, particularly the protein size of the bioproducts [15], which should be deeply studied.
Some of the important applications of ultrafiltration techniques include fractionation and the recovery of numerous proteins [16,17,18], polysaccharides [19,20], enzymes [1,21], biosurfactants [22], oils [23,24], and bioactive peptides [25,26]. Most of the studies on β-galactosidases reported the use of only ultrafiltration when modeling the hydrolysis mechanism [27] and galactooligosaccharide synthesis [28,29], and for the removal of impurities such as salts [30] and polymers in an aqueous purification system [31]. The application of these techniques often neglects the characteristic peculiarities of the bioproduct or does not consider the interactions between molecules, which in turn compromise the efficiency of the process and hinder the scale-up procedure, resulting in the infeasibility of the process [1].
Ultrafiltration can be used as an alternative to other concentration techniques; therefore, it is considered an important process for applications in the food and pharmaceutics industries. This technique can also be applied for the removal of salts and polyethylene glycol used in the purification process when utilizated in the diafiltration mode. In this case, ultrafiltration is achieved after precipitation or separation in an aqueous two-phase system of β-galactosidase from Kluyveromyces marxianus used in the production of GOS [5] or other products of industrial interest [32].
Thus, we evaluated the influence of the pH, membrane molecular weight cut-off (MWCO), operating pressure, and ionic strength of the ultrafiltration process of β-galactosidase from K. marxianus CCT 7082. Furthermore, the ultrafiltration process was applied in the diafiltration mode as an alternative for salt removal from the β-galactosidase extract after the purification process. To the best of our knowledge, no previous study has been performed in this regard.
2. Materials and Methods
2.1. Materials
The malt extract, yeast extract, peptone, glucose, agar, ammonium sulfate ((NH4)2SO4), potassium phosphate monobasic (KH2PO4), bovine serum albumin (BSA), and magnesium sulfate heptahydrate (MgSO4·7H2O) were purchased from Merck (Darmstadt, Germany). The lactose, potassium chloride (KCl), and sodium chloride (NaCl) were purchased from Synth (São Paulo, Brazil). Membranes with molecular weight cut-offs (MWCO) of 30 kDa (polyethersulfone—PBTK Millipore® 15005), 50 kDa (polyethersulfone—PBQK Millipore® 15005), 60 kDa (polysulfone—Osmonics® YMEWSP3001), and 100 kDa (polyethersulfone—PBHK Millipore® 15005) were purchased from Merck-Millipore (Darmstadt, Germany). The o-nitrophenyl-galactopyranoside used as the substrate was purchased from Sigma Aldrich (Darmstadt, Germany). All other chemicals used were of analytical grade and were used as received, without any further purification.
2.2. Microorganism, Inoculum, and Submerged Cultivation
The microorganism K. marxianus CCT 7082 was maintained at 4 °C in yeast malt (YM) agar slants containing malt extract (3 g/L), yeast extract (3 g/L), peptone (5 g/L), glucose (10 g/L), and agar (20 g/L) [33]. The inoculum was prepared in 500 mL Erlenmeyer flasks with 150 mL of culture medium containing yeast extract (1 g/L), (NH4)2SO4 (1.2 g·L−1), KH2PO4 (5 g/L), MgSO4·7H2O (0.4 g/L), and lactose (10 g/L) in 0.2 mol/L of potassium phosphate at pH 5.5. The culture conditions were 30 °C and 180 rpm for 24 h [34].
The inoculum was added to the cultivation medium in a proportion of 10 % (v/v). The medium contained yeast extract (17 g/L), (NH4)2SO4 (8.8 g/L), KH2PO4 (5 g/L), MgSO4.7H2O (0.4 g/L), and lactose (28.2 g/L) in 0.2 M of potassium phosphate buffer at pH 6.0. The microorganism was cultivated at 30 °C and 180 rpm for 96 h [33].
2.3. Enzyme Extraction
The enzyme was extracted by adding 1.1 g of glass beads (0.6–0.8 mm diameter) for each mL of cells resuspended (40 mg dry cell/mL) in a 50 mmol/L potassium phosphate buffer at pH 6.6 [35]. The suspension was stirred in a vortex mixer for 40 min; thereafter, the cell extract was clarified via centrifugation at 4700× g and 4 °C for 10 min [36].
2.4. β-Galactosidase Concentration Using Ultrafiltration Step
2.4.1. Assessment of Menbrane Molecular Weight Cut-Off and Operating pH and Pressure
The ultrafiltration runs were carried out in a stirred dead-end cell system (designed and produced by our research group and not commercially available) as illustrated in Figure 1. The system was kept under refrigeration at 4 °C to minimize losses due to denaturation. Membranes with molecular weight cut-offs of 30, 50, 60, and 100 kDa were tested. The operating pressure and pH values varied from 1.5 to 2.5 kgf/cm2 and from 6.5 to 7.5, respectively. All assays were performed in triplicate. The enzymatic activity and total protein content values in the feed, retentate, and permeate were measured after each test.
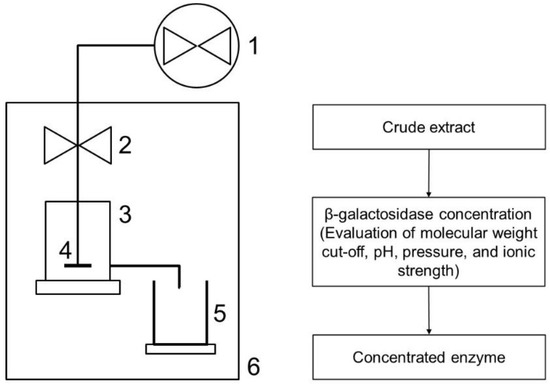
Figure 1.
Scheme of the UF system used for β-galactosidase concentration: (1) nitrogen cylinder with valve; (2) pressure control; (3) ultrafiltration cell; (4) stirrer; (5) collector; (6) refrigeration.
2.4.2. Ionic Strength Influence
The increases in ionic strength in the enzyme concentration and prepurification processes were evaluated by adding potassium chloride (KCl) and sodium chloride (NaCl) to the initial enzyme extract. The runs were performed in triplicate with the following concentrations: 0.01, 0.05, and 0.1 mol/L. The system was operated at 1.5 kgf/cm2, pH 7.5, and 4 °C, using a UF membrane of 50 kDa.
2.4.3. Diafiltration
The diafiltration was carried out using the same ultrafiltration apparatus to remove the ammonium sulfate, sodium chloride, and potassium phosphate salts homogenized with the enzyme extract obtained from published and recommended protocols for precipitation and ion-exchange chromatography elution and as part of the composition of the aqueous two-phase system (ATPS), respectively [1,37,38]. The enzymatic extracts were obtained after reproducing the protocols and the purified enzymatic extract, which was rich in salt and used in the diafiltration process. The complete removal of the salts was performed using six diafiltration cycles with two volumes of diafiltration solution per cycle (50 mmol/L potassium phosphate buffer) at a suitable pH for each case.
2.5. Ultrafiltration Performance
The performance in the enzyme recovery and concentration processes was evaluated using the recovery (Equation (1)), purification (Equation (2)), retention and concentration factors (retained proteins (Equation (3)), volume concentration factor (Equation (4)), activity (Equation (5)) and protein concentration factors (Equation (6)), flux (Equation (7)), and yield of the processes (Equation (8)). The uncertainties of the measured values were, on average, below 5%:
where Asf and Asi are the respective specific activities (U/mg) of the final and initial (crude extract) fractions. The specific activity is defined as the ratio between the activity (U/mg) and the total protein concentration (mg/mg) in the same sample:
where Af and Ai are the respective enzymatic activities (U/mL) of the final and initial (crude extract) fractions; Vf and Vi are the volumes (mL) of the final and initial (crude extract) fractions, respectively:
where Pcp and Pcf are the protein concentrations (mg/mL) in the permeate and feed, respectively:
where Vf and Vr are the volume (mL) fed into the ultrafiltration system and the volume remaining after permeation:
where Af and Ar are the activity levels(U/mL) at the beginning of the process and the end of the process:
where Pf and Pr are the protein concentrations (mg/mL) at the beginning of the process and the end of the process:
where Vp is the permeate volume (L), A is the membrane area (m2), and t is the time (h):
where At is the total activity (U) obtained in the final process and t is the total time of the process (h).
2.6. Analytical Methods
Enzyme Activity and Total Protein
The enzyme activity was determined using o-nitrophenyl-galactopyranoside as the substrate (ONPG, Aldrich), according to the method proposed by Inchaurrondo, et al. [39]. One unit of enzyme activity (U) is defined as the amount of enzyme necessary to yield 1 μmol of o-nitrophenol per minute at 37 °C and pH 6.6. The total protein content was assayed using the method proposed by Bradford [40], using bovine serum albumin (BSA) as the standard.
2.7. Statistical Analysis
All analyses were performed in triplicate. The results are presented as means ± standard deviations from the mean. The results were analyzed using a unilateral analysis of variance (ANOVA), and Tukey’s test (p < 0.05) was used to determine differences between means using Statistica 7.0 for Windows (Statsoft Inc., Tulsa, OK, USA).
3. Results and Discussion
3.1. β-Galactosidase Recovery and Concentration Using Ultrafiltration
The use of enzymatic extracts at various pH values and membranes with different molecular weight cut-offs (MWCO) showed no significant changes (p < 0.05) in the parameters evaluated during the β-galactosidase ultrafiltration process (Table 1). All runs, except for the 30 and 50 kDa membranes at pH 6.5, presented purification factors >1. These low purification factors might be due to the similar sizes of the contaminating proteins and the target enzyme. Another hypothesis for the low purification factor found in the present work is the retention caused by the dynamic membrane formed by the proteins retained on the membrane surface layer (gel layer) [41], which contributes to an increase in filtration resistance, rapid flow reduction, and the shortening of the operational cycles, affecting the process viability [42].

Table 1.
Effects of membrane molecular weight cut-offs on β-galactosidase concentrations at pH 6.5, 7.0, and 7.5.
The runs using the 50 kDa membrane at pH 7.5 exhibited the highest enzyme recovery rate (108.9%). This behavior can be explained by the presence of metabolites during the purification process, which can inhibit the enzyme activity. When the contaminants are removed from the medium, the enzyme may be activated [43,44,45].
All membranes tested, regardless of the pH, were able to fully retain the enzyme, while the enzymatic activity was not verified in the permeate flux. The high recovery rates of enzymatic activity (in relation to the total activity at the beginning of the process) ranged from 79.5 to 108.9%. The volume, activity, and protein concentration factor values were up to 8.1, 7.1, and 6.4-fold greater, respectively.
These results demonstrated the effectiveness of the ultrafiltration steps for the recovery and concentration of β-galactosidase. Furthermore, the ultrafiltration reduced the working volume, which is a crucial factor in the purification processes [1].
Moreover, the permeate flux was evaluated during the ultrafiltration process, since a low flux can lead to a lengthy and expensive process, making it one of the factors limiting deployment [12]. Figure 2A–C shows the permeate flux variation during ultrafiltration at pH 6.5, 7.0, and 7.5, respectively.
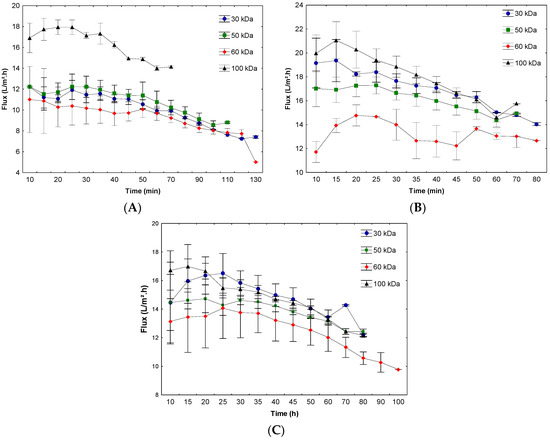
Figure 2.
Permeate flux during ultrafiltration at pH 6.5 (A), 7.0 (B), and 7.5 (C).
In the present work, a decrease in flux during permeation was observed in all conditions, which is considered a typical behavior of concentration polarization phenomena and fouling on the membrane surface. When the solution is permeated, the protein concentration at the membrane surface increases, thereby elevating the polarized layer. Depending on the situation, a gel layer may be formed, which in turn leads to a further decrease in the permeate flux [15].
At pH 7.5, low variation in the initial permeate flux as a function of the molar mass cut-off was observed, with the values ranging from 16.0 to 13.1 L/m2·h, representing a decrease of 18.0%. Throughout the process, the highest variation observed was from 16.7 to 12.1 L/m2.h, representing a reduction of 27.5%, with a maximum operation time of 100 min.
The lowest initial permeate flux, except for the 100 kDa membrane, occurred at pH 6.5 (range of 12.2 to 11.1 L/m2·h). The largest decrease in permeate flux was observed in the 60 kDa membrane (11.0 to 5.0 L/m2·h), representing a decrease of 54.5% over the process and a maximum operating time of 130 min. The low permeate flux at this pH may have been due to protein agglomeration and the production of a deposit on the membrane surface, thereby reducing the membrane permeability. Furthermore, the protein adsorption on the membrane surface may result in a second membrane that alters the function of the original membrane [46,47,48].
Although the 60 kDa membrane presented similar results to the other membranes regarding the purification factor, this membrane should be analyzed and used with caution due to the process presenting a longer operating time and a lower final permeate flux when compared to the other membranes, affecting the yield and viability of the process. This reduction in the yield of the process can lead to significant increases in the concentration and purification costs of this enzyme and can even make it unfeasible at the industrial scale. This effect may have occurred due to greater fouling of the membrane, as a larger pore diameter favors the permeation of compounds similar in size to the membrane pores, leading to a greater occurrence of blockage of the pores [12]. In addition, the differences in membrane material can also affect the separation performance during ultrafiltration, since the chemical composition can affect the functionality, charge, and interactions with the material, among other factors [49].
At pH 7.0, the average flux values ranged from 21.2 to 12.0 L/m2·h, with a reduction of 43.4% at a maximum operating time of 80 min. Although the application of this pH led to the highest initial flow, a high flux reduction rate was observed during the ultrafiltration process. The permeate flux variation is a phenomenon inherent in the process and is observed in several processes using ultrafiltration. Cassini, et al. [50] studied three ultrafiltration membranes used for recovering soy protein isolates. These authors reported reductions in the permeate flux by 30%, 69%, and 30% for the 5, 20, and 50 kDa membranes, respectively. Similarly, Chollangi and Hossain [51] reported fluxes of 10 and 125 L/m2·h for the 5 and 10 kDa membranes, respectively, using a pressure of 3.0 kgf/cm2 in the ultrafiltration of proteins and lactose from dairy effluents. These authors observed a decrease in the permeate flux, indicating that the protein increased fouling on the membrane and thereby affected the permeate flux.
The evaluation of all process factors—including the flow and its variations throughout the process, the enzymatic recovery and purification values, as well as the concentration and retention parameters for the protein and enzymatic activity—is necessary to define the conditions for carrying out ultrafiltration [1,21].
Based on the previous results, the effect of the operating pressure was assessed using the 50 kDa membrane at pH 7.5, which presented higher purification factor and enzyme recovery values and a slight decrease in the permeate flux during β-galactosidase ultrafiltration. Figure 3 presents these results.
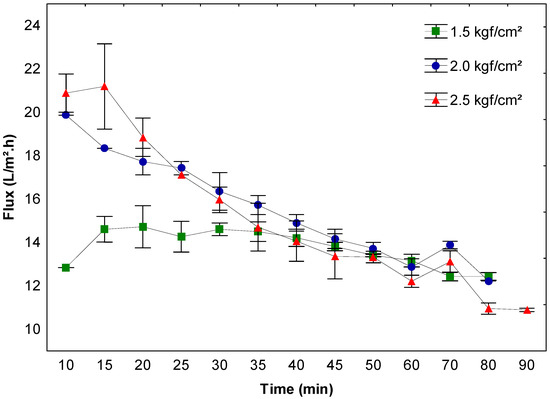
Figure 3.
Permeate flux and effect on the enzyme concentration using a 50 kDa membrane at pH 7.5 and 1.5, 2.0, and 2.5 kgf/cm2 operating pressures.
After comparing the initial flux with the final flux in the process at different pressures (Figure 3), decreases of approximately 3.5%, 38.4%, and 47.6% were observed, respectively, indicating that the increase in pressure causes a significant decrease in permeate flux during the process. In the runs operated at low pressures, the concentration was gradual, leading to a slight drop in the permeate flux, a consequent reduction in the operating time, and an increase in the process yield (734.1 U/h) when compared to those at higher pressures (484.0 and 453.7 U/h for pressures of 2.0 and 2.5 kgf/cm2, respectively). Additionally, a comparison between pressures of 2.0 and 2.5 kgf/cm2 also showed the effect of higher pressures on the membrane concentration process, where an inversion of permeation efficiency was observed over time, especially at 25 min. Despite initially presenting a higher flux, the pressure of 2.5 kgf/cm2 resulted in the faster concentration of the proteins (see the description of the effects below), altering the permeation capacity of the membrane, which made the process less efficient over time when compared to the process using a pressure of 2.0 kgf/cm2, for example.
After 35 min, no difference was observed between the pressures tested, and from this point onwards the processes seem equivalent. However, it is important to consider the advantages that reduce the operating costs and make the process viable, such as the constant filtration rate and low flow variation, the permeation of smaller particles present in the material, the membrane deformation, as well as consumption of gas used in the membrane permeation process [12,52,53].
The reduced recovery and purification factors of the enzyme in the systems operating with higher pressures may be associated with greater flux reductions caused by the membrane deformation (related to the compaction that occurs depending on the time of use, the constant conditions of high pressure, and the material used, which influences the permeability and increases the filtration resistance), concentration polarization, and fouling due to the applied pressure, leading to the inefficiency of the process; however, the occurrence of these phenomena is inevitable, can harm the performance of the membrane, and influence the process viability [54,55].
The decrease in resistance with increasing pressure is generally not observed in permeation experiments. In general, the opposite contradictory trend is observed; that is, the higher pressure results in the increased formation of the second layer on the membrane, thereby reducing the permeability [56,57].
Although working at reduced pressures with lower flux may seem inevitable, the results indicate that under less polarized conditions, the fouling is lower and the permeate flux is less affected during the process.
Sodium and potassium chloride were added to the enzymatic extract of β-galactosidase, as these salts can change the ionic strength of the solution [58], act as protective agents of the enzyme [59], and modify the permeability during the separation of some compounds [58,60,61]. However, the addition of NaCl and KCl in the β-galactosidase extract revealed no positive effect compared to the control test (without added salt) (Table 2) and reduced the activity, purity, concentration, and enzyme recovery. The retention of the activity and protein levels did not change when compared to the control assay.
Table 2.
Effects of ionic strength on the enzyme concentration and purification using a 50 kDa membrane at pH 7.5 and 1.5 kgf/cm2 of operating pressure.
The addition of salt to the extract did not cause migration of the enzyme or increased migration of contaminants to permeate proteins (data not shown). Although the protective effect of KCl was not observed, the denaturation of the enzyme was lower when compared to the NaCl recovery. The same behavior was observed by Braga, Lemes and Kalil [37] in β-galactosidase purification via ion-exchange chromatography. These authors found that the purification factor and enzyme recovery were higher when the KCl in the same concentration used in the present work (1 M) was used in the purification when compared to NaCl and ammonium sulfate.
The purification factor and enzyme recovery during the ultrafiltration decreased when the sodium and potassium chloride concentrations were increased from 0.01 M to 0.1 M. The prolonged contact of the enzyme with the salts seemed to have caused enzymatic denaturation. In addition, several changes were observed in the structure of the molecules and the electrostatic interactions between proteins, thereby resulting in protein agglomeration and consequent deposition and adsorption in the membrane surface to a larger extent [48,62].
3.2. Diafiltration
The removal of salts used in the purification processes is essential for maintaining the enzyme catalytic power, since it is known that large salt concentrations in certain cases can affect and even denature the target enzyme. This evidence reveals the importance of this process [63,64].
Thus, the ultrafiltration process was applied in diafiltration mode as an alternative to remove salts and other remaining contaminants at the end of the previously established purification procedures (aqueous two-phase system, precipitation, and ion-exchange chromatography) [1]. The fractions containing the purified enzyme derived from various purification procedures were diafiltered by adding a solute-free solvent (potassium phosphate buffer 0.05 M) and the filtration was stopped when the target salts were not detected in the enzyme extract. The number of cycles (6 cycles), as previously established for ammonium sulfate, was considered equivalent for other salts [65]. Moreover, our research group previously defined that at least five cycles were required for the total removal of the remaining PEG from the aqueous two-phase system in purification steps [21].
The combination of ultrafiltration and diafiltration processes was effective, particularly for concentrating and desalting the enzyme extract; furthermore, the total recovery of the sample was possible. The β-galactosidase recovery rate via diafiltration, regardless of the salt used, was always greater than 90% (Table 3), thereby indicating that this process is feasible for removing the salts present in the enzyme extract without significantly affecting the enzyme recovery.
Table 3.
Purification factor and β-galactosidase recovery in the diafiltration process after different techniques.
Another advantage of the diafiltration mode is associated with the possibility of reducing the concentration polarization and membrane fouling effects, in certain cases [66]. Thus, the choice of operating parameters is crucial in achieving good recovery of the enzyme. The diafiltration mode did not provide high purification factors; however, the process was effective for the complete removal of the salts present in the extract.
4. Conclusions
The ultrafiltration and concentration of β-galactosidase from Kluyveromyces marxianus CCT 7082 were efficiently applied, despite not promoting high purification factors (=1.0-fold). In this case, the use of ultrafiltration at pH 7.5 and 1.5 kgf/cm2 with a 50 kDa membrane resulted in the maximum concentration of beta-galactosidase extract (7.1-fold) and 100% enzyme recovery. The increase in operating pressure caused losses in the enzyme recovery of up to 38% of the process yield due to the greater flux reduction caused by the membrane deformation, concentration polarization, and fouling due to the applied pressure, leading to inefficiency in the process. The use of salts in the feed solution (KCl and NaCl) caused decreases in enzyme purification and recovery of up to 40% during the process using NaCl, probably due to protein denaturation. The diafiltration mode was efficient for salt removal from the purified enzymatic extract using different techniques, including an aqueous biphasic system, ion exchange chromatography, and ammonium sulfate precipitation, thereby indicating its potential application (recovery >93.4% and purification factor of 1.0-fold) compared to other more expensive and laborious techniques.
Author Contributions
Conceptualization and methodology, A.C.L., F.d.O.M., A.d.S.F., M.B.E., M.D.L. and S.J.K.; investigation, A.C.L., F.d.O.M. and A.d.S.F.; writing—original draft preparation, A.C.L.; writing—review and editing, A.C.L., M.B.E., M.D.L. and S.J.K.; visualization and supervision, M.D.L. and S.J.K.; project administration, S.J.K.; funding acquisition, S.J.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
This work was supported by FAPERGS, CAPES, CNPq, and IF Goiano (process no. 23218.001212.2021-05).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lemes, A.C.; Machado, J.R.; Brites, M.L.; Di-Luccio, M.; Kalil, S.J. Design Strategies for Integrated β-Galactosidase Purification Processes. Chem. Eng. Technol. 2014, 37, 1–9. [Google Scholar]
- Dekker, P.J.T.; Koenders, D.; Bruins, M.J. Lactose-Free Dairy Products: Market Developments, Production, Nutrition and Health Benefits. Nutrients 2019, 11, 551. [Google Scholar] [CrossRef] [PubMed]
- Kocabaş, H.; Ergin, F.; Aktar, T.; Küçükçetin, A. Effect of lactose hydrolysis and salt content on the physicochemical, microbiological, and sensory properties of ayran. Int. Dairy J. 2022, 129, 105360. [Google Scholar] [CrossRef]
- Ugidos-Rodríguez, S.; Matallana-González, M.C.; Sánchez-Mata, M.C. Lactose malabsorption and intolerance: A review. Food Funct. 2018, 9, 4056–4068. [Google Scholar] [CrossRef] [PubMed]
- Braga, A.R.C.; Manera, A.P.; Ores, J.D.; Sala, L.; Maugeri, F.; Kalil, S.J. Kinetics and Thermal Properties of Crude and Purified β-Galactosidase with Potential for the Production of Galactooligosaccharides. Food Technol. Biotechnol. 2013, 51, 45–52. [Google Scholar]
- Yu, L.; O’Sullivan, D.J. Production of galactooligosaccharides using a hyperthermophilic β-galactosidase in permeabilized whole cells of Lactococcus lactis. J. Dairy Sci. 2014, 97, 694–703. [Google Scholar] [CrossRef]
- Huang, J.; Zhu, S.; Zhao, L.; Chen, L.; Du, M.; Zhang, C.; Yang, S.-T. A novel β-galactosidase from Klebsiella oxytoca ZJUH1705 for efficient production of galacto-oligosaccharides from lactose. Appl. Microbiol. Biotechnol. 2020, 104, 6161–6172. [Google Scholar] [CrossRef]
- Tomal, A.A.B.; Farinazzo, F.S.; Bachega, A.; Bosso, A.; Silva, J.B.; Suguimoto, H. Galacto Oligosaccharides and Human Health Implications. Nutr. Food Sci. Int. J. 2019, 9, 1–4. [Google Scholar]
- Asraf, S.S.; Gunasekaran, P. Current trends of β-galactosidase research and application. Appl Microbiol Biot 2010, 1, 880–890. [Google Scholar]
- Schmidt, C.; Mende, S.; Jaros, D.; Rohm, H. Fermented milk products: Effects of lactose hydrolysis and fermentation conditions on the rheological properties. Dairy Sci. Technol. 2016, 96, 199–211. [Google Scholar] [CrossRef]
- Lima, P.C.; Gazoni, I.; de Carvalho, A.M.G.; Bresolin, D.; Cavalheiro, D.; de Oliveira, D.; Rigo, E. β-galactosidase from Kluyveromyces lactis in genipin-activated chitosan: An investigation on immobilization, stability, and application in diluted UHT milk. Food Chem. 2021, 349, 129050. [Google Scholar] [CrossRef] [PubMed]
- Lemes, A.C.; Braga, A.R.C.; Gautério, G.V.; Fernandes, K.F.; Egea, M.B. Application of Membrane Technology for Production of Bioactive Peptides. In Bioactive Peptides: Production, Bioavailability, Health Potential and Regulatory Issues; Onuh, J.O., Selvamuthukumaran, M., Pathak, Y.V., Eds.; CRC Press: Boca Raton, FL, USA, 2021; Volume 1, pp. 253–279. [Google Scholar]
- Cui, Z. Protein separation using ultrafiltration: An example of multi-scale complex systems. China Particuology 2005, 3, 343–348. [Google Scholar] [CrossRef]
- Saxena, A.; Tripathi, B.P.; Kumar, M.; Shahi, V.K. Membrane-based techniques for the separation and purification of proteins: An overview. Adv. Colloid Interface Sci. 2009, 145, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Cui, Z.F. Protein purification by ultrafiltration with pre-treated membrane. J. Membr. Sci. 2000, 167, 47–53. [Google Scholar] [CrossRef]
- Lu, R.R.; Xu, S.Y.; Wang, Z.; Yang, R.J. Isolation of lactoferrin from bovine colostrum by ultrafiltration coupled with strong cation exchange chromatography on a production scale. J. Membr. Sci. 2007, 297, 152–161. [Google Scholar] [CrossRef]
- Van Audenhaege, M.; Garnier-Lambrouin, F.; Piot, M.; Gésan-Guiziou, G. Unexpected displacement of the equilibrium between the apo and the holo form during ultrafiltration of the metalloprotein α-lactalbumin. J. Membr. Sci. 2012, 401–402, 195–203. [Google Scholar] [CrossRef]
- Iltchenco, S.; Preci, D.; Bonifacino, C.; Fraguas, E.F.; Steffens, C.; Panizzolo, L.A.; Colet, R.; Fernandes, I.A.; Abirached, C.; Valduga, E.; et al. Whey protein concentration by ultrafiltration and study of functional properties. Ciência Rural 2018, 48, 778–780. [Google Scholar] [CrossRef]
- Patsioura, A.; Galanakis, C.M.; Gekas, V. Ultrafiltration optimization for the recovery of β-glucan from oat mill waste. J. Membr. Sci. 2011, 373, 53–63. [Google Scholar] [CrossRef]
- Xie, J.H.; Shen, M.Y.; Nie, S.P.; Zhao, Q.; Li, C.; Xie, M.Y. Separation of water-soluble polysaccharides from Cyclocarya paliurus by ultrafiltration process. Carbohydr. Polym. 2014, 101, 479–483. [Google Scholar] [CrossRef]
- Sala, L.; Gautério, G.V.; Younan, F.F.; Brandelli, A.; Moraes, C.C.; Kalil, S.J. Integration of ultrafiltration into an aqueous two-phase system in the keratinase purification. Process Biochem. 2014, 49, 2016–2024. [Google Scholar] [CrossRef]
- Long, X.; Meng, Q.; Sha, R.; Huang, Q.; Zhang, G. Two-step ultrafiltration of rhamnolipids using PSU-g-PEG membrane. J. Membr. Sci. 2012, 409–410, 105–112. [Google Scholar] [CrossRef]
- Tres, M.V.; Racoski, J.C.; Di Luccio, M.; Oliveira, J.V.; Treichel, H.; de Oliveira, D.; Mazutti, M.A. Separation of soybean oil/n-hexane and soybean oil/n-butane mixtures using ceramic membranes. Food Res Int 2014, 63 Part A, 33–41. [Google Scholar] [CrossRef]
- Biron, D.d.S.; Zeni, M.; Bergmann, C.P.; Santos, V.d. Analysis of Composite Membranes in the Separation of Emulsions Sunflower oil/water. Mater. Res. 2017, 20, 843–852. [Google Scholar] [CrossRef]
- Firdaous, L.; Dhulster, P.; Amiot, J.; Gaudreau, A.; Lecouturier, D.; Kapel, R.; Lutin, F.; Vézina, L.-P.; Bazinet, L. Concentration and selective separation of bioactive peptides from an alfalfa white protein hydrolysate by electrodialysis with ultrafiltration membranes. J. Membr. Sci. 2009, 329, 60–67. [Google Scholar] [CrossRef]
- Lemes, A.C.; Sala, L.; Ores, J.C.; Braga, A.R.; Egea, M.B.; Fernandes, K.F. A Review of the Latest Advances in Encrypted Bioactive Peptides from Protein-Rich Waste. Int. J. Mol. Sci. 2016, 17, 950. [Google Scholar] [CrossRef] [PubMed]
- Hatzinikolaou, D.G.; Katsifas, E.; Mamma, D.; Karagouni, A.D.; Christakopoulos, P.; Kekos, D. Modeling of the simultaneous hydrolysis–ultrafiltration of whey permeate by a thermostable β-galactosidase from Aspergillus niger. Biochem. Eng. J. 2005, 24, 161–172. [Google Scholar] [CrossRef]
- Chockchaisawasdee, S.; Athanasopoulos, V.I.; Niranjan, K.; Rastall, R.A. Synthesis of galacto-oligosaccharide from lactose using beta-galactosidase from Kluyveromyces lactis: Studies on batch and continuous UF membrane-fitted bioreactors. Biotechnol. Bioeng. 2005, 89, 434–443. [Google Scholar] [CrossRef]
- Palai, T.; Mitra, S.; Bhattacharya, P.K. Kinetics and design relation for enzymatic conversion of lactose into galacto-oligosaccharides using commercial grade beta-galactosidase. J. Biosci. Bioeng. 2012, 114, 418–423. [Google Scholar] [CrossRef]
- Hemavathi, A.B.; Raghavarao, K.S.M.S. Differential partitioning of β-galactosidase and β-glucosidase using aqueous two phase extraction. Process Biochem. 2011, 46, 649–655. [Google Scholar] [CrossRef]
- Veide, A.; Lindbäck, T.; Enfors, S.-O. Recovery of β-galactosidase from a poly (ethylene glycol) solution by diafiltration. Enzym. Microb Tech. 1989, 11, 744–751. [Google Scholar] [CrossRef]
- Lemes, A.C.; Pavón, Y.; Lazzaroni, S.; Rozycki, S.; Brandelli, A.; Kalil, S.J. A new milk-clotting enzyme produced by Bacillus sp. P45 applied in cream cheese development. LWT Food Sci. Technol. 2016, 66, 217–224. [Google Scholar] [CrossRef]
- Manera, A.P.; Ores, J.C.; Ribeiro, V.A.; Burkert, C.A.V.; Kalil, S.J. Optimization of the culture medium for the production of beta-galactosidase from Kluyveromyces marxianus CCT 7082. Food Technol. Biotech. 2008, 46, 66–72. [Google Scholar]
- Pinheiro, R.; Belo, I.; Mota, M. Growth and beta-galactosidase activity in cultures of Kluyveromyces marxianus under increased air pressure. Lett. Appl. Microbiol. 2003, 37, 438–442. [Google Scholar] [CrossRef]
- Medeiros, F.O.; Veiga Burkert, C.A.; Juliano Kalil, S. Purification of β-Galactosidase by Ion Exchange Chromatography: Elution Optimization Using an Experimental Design. Chem. Eng. Technol. 2012, 35, 911–918. [Google Scholar] [CrossRef]
- Medeiros, F.O.; Alves, F.G.; Lisboa, C.R.; Martins, D.S.; Burkert, C.A.V.; Kalil, S.J. Ondas ultrassônicas e pérolas de vidro: Um novo método de extração de β-galactosidase para uso em laboratório. Química Nova 2008, 31, 336–339. [Google Scholar] [CrossRef]
- Braga, A.R.C.; Lemes, A.C.; Kalil, S.J. Single Chromatographic Step for β-Galactosidase Purification: Influence of Salt and Elution Parameters. Sep. Sci. Technol. 2014, 49, 1817–1824. [Google Scholar] [CrossRef]
- Heidtmann, R.B.; Duarte, S.H.; Pereira, L.P.; Braga, A.R.C.; Kalil, S.J. Kinetics and thermodynamic characterization of β-galactosidase from Kluyveromyces marxianus CCT 7082 fractionated with ammonium sulphate. Braz. J. Food Technol. 2012, 15, 41–49. [Google Scholar] [CrossRef]
- Inchaurrondo, V.A.; Flores, M.V.; Voget, C.E. Growth and β-galactosidase synthesis in aerobic chemostat cultures of Kluyveromyces lactis. J. Ind. Microbiol. Biotech. 1998, 20, 291–298. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Ghosh, R.; Cui, Z.F. Purification of lysozyme using ultrafiltration. Biotechnol. Bioeng. 2000, 68, 191–203. [Google Scholar] [CrossRef]
- Ni, L.; Shi, Q.; Wu, M.; Ma, J.; Wang, Y. Fouling behavior and mechanism of hydrophilic modified membrane in anammox membrane bioreactor: Role of gel layer. J. Membr. Sci. 2021, 620, 118988. [Google Scholar] [CrossRef]
- Cortés, G.; Trujillo-Roldán, M.A.; Ramírez, O.T.; Galindo, E. Production of β-galactosidase by Kluyveromyces marxianus under oscillating dissolved oxygen tension. Process Biochem. 2005, 40, 773–778. [Google Scholar] [CrossRef]
- Pan, I.H.; Yao, H.J.; Li, Y.K. Effective extraction and purification of β-xylosidase from Trichoderma koningii fermentation culture by aqueous two-phase partitioning. Enzym. Microb. Tech. 2001, 28, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Porto, T.S.; Medeiros e Silva, G.M.; Porto, C.S.; Cavalcanti, M.T.H.; Neto, B.B.; Lima-Filho, J.L.; Converti, A.; Porto, A.L.F.; Pessoa, A., Jr. Liquid–liquid extraction of proteases from fermented broth by PEG/citrate aqueous two-phase system. Chem. Eng. Process. Process Intensif. 2008, 47, 716–721. [Google Scholar] [CrossRef]
- Saha, N.K.; Balakrishnan, M.; Ulbricht, M. Polymeric membrane fouling in sugarcane juice ultrafiltration: Role of juice polysaccharides. Desalination 2006, 189, 59–70. [Google Scholar] [CrossRef]
- Bullón, J.; Belleville, M.P.; Rios, G.M. Preparation of gelatin formed-in-place membranes: Effect of working conditions and substrates. J. Membr. Sci. 2000, 168, 159–165. [Google Scholar] [CrossRef]
- Palecek, S.P.; Mochizuki, S.; Zydney, A.L. Effect of ionic environment on BSA filtration and the properties of BSA deposits. Desalination 1993, 90, 147–159. [Google Scholar] [CrossRef]
- Kumar, R.; Ismail, A.F. Fouling control on microfiltration/ultrafiltration membranes: Effects of morphology, hydrophilicity, and charge. J. Appl. Polym. Sci. 2015, 132, 1–20. [Google Scholar] [CrossRef]
- Cassini, A.S.; Tessaro, I.C.; Marczak, L.D.F.; Pertile, C. Ultrafiltration of wastewater from isolated soy protein production: A comparison of three UF membranes. J. Clean. Prod. 2010, 18, 260–265. [Google Scholar] [CrossRef]
- Chollangi, A.; Hossain, M.M. Separation of proteins and lactose from dairy wastewater. Chem. Eng. Process. Process Intensif. 2007, 46, 398–404. [Google Scholar] [CrossRef]
- Shahid, M.; Faure, C.; Ottoboni, S.; Lue, L.; Price, C. Employing Constant Rate Filtration To Assess Active Pharmaceutical Ingredient Washing Efficiency. Org. Process Res. Dev. 2022, 26, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.S.; Bilad, M.R.; Shamsuddin, N.; Suhaimi, H.; Ismail, N.M.; Jaafar, J.; Ismail, A.F. Confounding Effect of Wetting, Compaction, and Fouling in an Ultra-Low-Pressure Membrane Filtration: A Review. Polymers 2022, 14, 2073. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Leow, H.F. Microfiltration of activated sludge wastewater—The effect of system operation parameters. Sep. Purif. Technol. 2002, 29, 189–198. [Google Scholar] [CrossRef]
- Aghajani, M.; Maruf, S.H.; Wang, M.; Yoshimura, J.; Pichorim, G.; Greenberg, A.; Ding, Y. Relationship between permeation and deformation for porous membranes. J. Membr. Sci. 2017, 526, 293–300. [Google Scholar] [CrossRef]
- Van de Ven, W.J.C.; Sant, K.v.t.; Pünt, I.G.M.; Zwijnenburg, A.; Kemperman, A.J.B.; van der Meer, W.G.J.; Wessling, M. Hollow fiber dead-end ultrafiltration: Influence of ionic environment on filtration of alginates. J. Membr. Sci. 2008, 308, 218–229. [Google Scholar] [CrossRef]
- Leberknight, J.; Wielenga, B.; Lee-Jewett, A.; Menkhaus, T.J. Recovery of high value protein from a corn ethanol process by ultrafiltration and an exploration of the associated membrane fouling. J. Membr. Sci. 2011, 366, 405–412. [Google Scholar] [CrossRef]
- Becht, N.O.; Malik, D.J.; Tarleton, E.S. Evaluation and comparison of protein ultrafiltration test results: Dead-end stirred cell compared with a cross-flow system. Sep. Purif. Technol. 2008, 62, 228–239. [Google Scholar] [CrossRef]
- Athès, V.; Combes, D. Influence of Additives on High Pressure Stability of β-Galactosidase from Kluyveromyces Lactis and Invertase from Saccharomyces Cerevisiae. Enzym. Microb. Tech. 1998, 22, 532–537. [Google Scholar] [CrossRef]
- Lin, S.H.; Hung, C.L.; Juang, R.S. Effect of operating parameters on the separation of proteins in aqueous solutions by dead-end ultrafiltration. Desalination 2008, 234, 116–125. [Google Scholar] [CrossRef]
- Teng, M.-Y.; Lin, S.-H.; Wu, C.-Y.; Juang, R.-S. Factors affecting selective rejection of proteins within a binary mixture during cross-flow ultrafiltration. J. Membr. Sci. 2006, 281, 103–110. [Google Scholar] [CrossRef]
- Kelly, S.T.; Zydney, A.L. Protein fouling during microfiltration: Comparative behavior of different model proteins. Biotechnol. Bioeng. 1997, 55, 91–100. [Google Scholar] [CrossRef]
- Lebendiker, M.; Danieli, T. Production of prone-to-aggregate proteins. FEBS Lett. 2014, 588, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Hamada, H.; Arakawa, T.; Shiraki, K. Effect of additives on protein aggregation. Curr. Pharm. Biotechnol. 2009, 10, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Lemes, A.C. Purification of β-galactosidase: Process design. In School of Chemistry and Food; Federal University of Rio Grande: Rio Grande, Brazil, 2015; 95p, Available online: https://repositorio.furg.br (accessed on 26 December 2022).
- Duarte, R.M.B.O.; Santos, E.B.H.; Duarte, A.C. Comparison between diafiltration and concentration operation modes for the determination of permeation coefficients of humic substances through ultrafiltration membranes. Anal. Chim. Acta 2001, 442, 155–164. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).