
Optimization of an Innovative Hydrothermal Processing on Prebiotic Properties of Eucheuma denticulatum, a Tropical Red Seaweed


Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Seaweed Sample Preparation
2.3. Hydrolysis and Extraction of Seaweed Oligosaccharide Fraction
2.4. Prebiotic Activity Assay and Prebiotic Activity Score
enteric (Δ log CFU/mL prebiotic/Δ log CFU/mL glucose)
2.5. Quantification of HMF
2.6. Selection of Three Levels of Independent Variables (X1, X2, X3)
2.7. Experimental Design and Validation of Models
2.8. FT-IR Spectra Acquisition
2.9. Statistical Analysis
3. Results
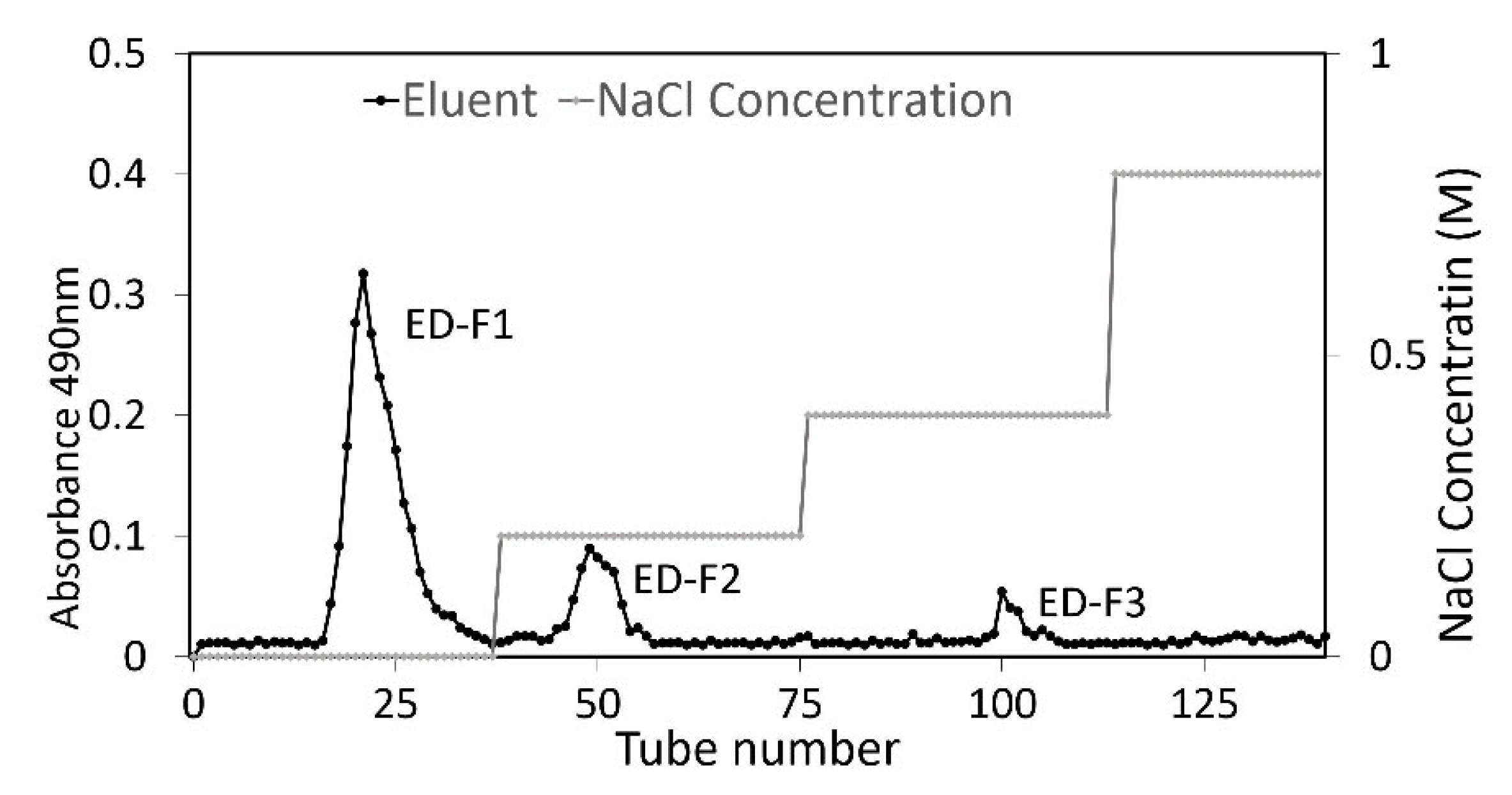
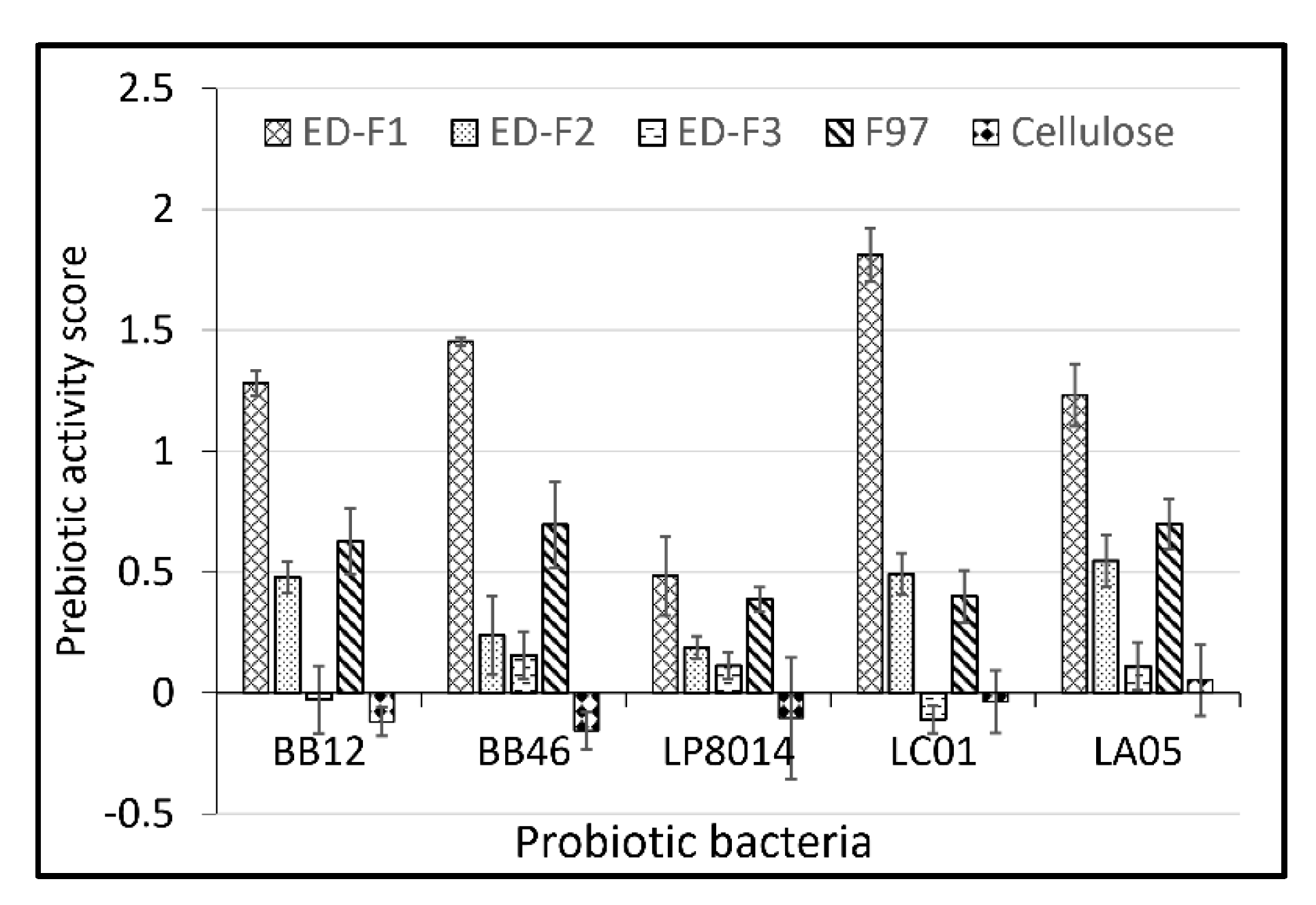
3.1. Purification and Prebiotic Activity Score of Red Seaweed Oligosaccharide Fractions
3.2. Selection of Factor Levels
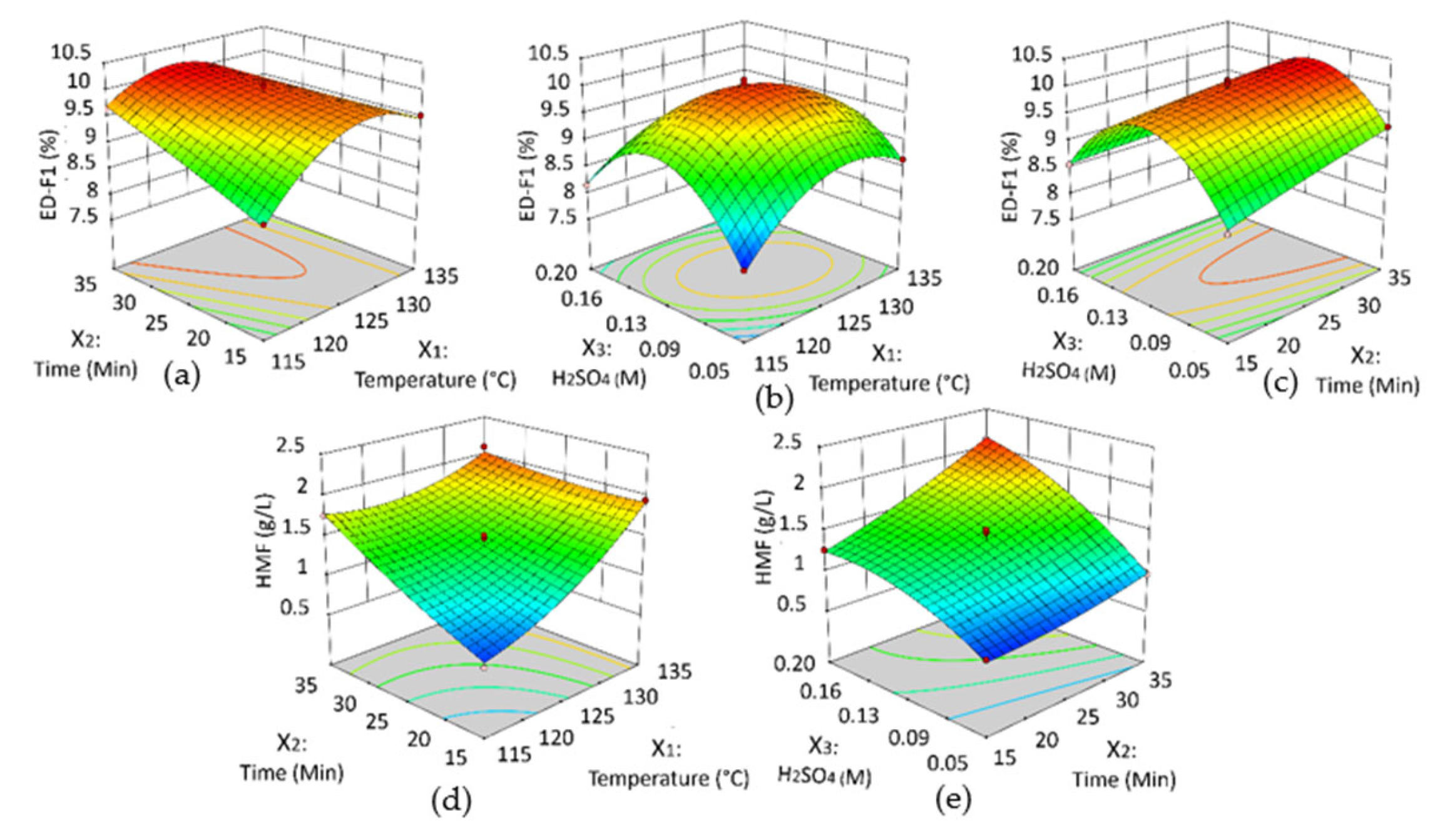
3.3. RSM Model and Analysis of Variance
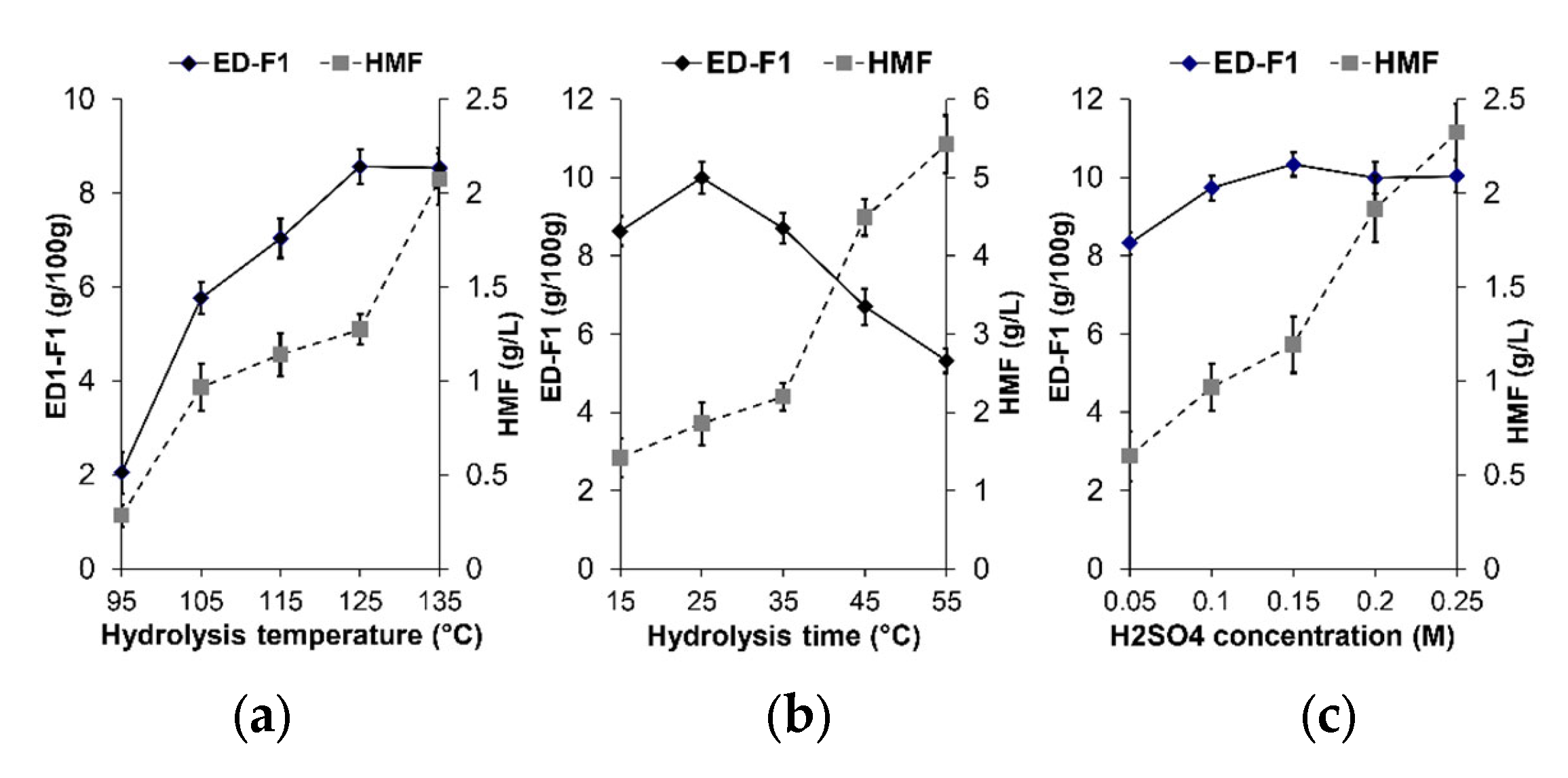
3.4. Effects of Hydrolysis Parameters on the Yield of Oligosaccharides Fraction ED-F1 and HMF By-Products
3.5. Validation of the Model and Optimal Extraction Condition of ED-F1 Oligosaccharide Fraction
3.6. Characterization of ED-F1
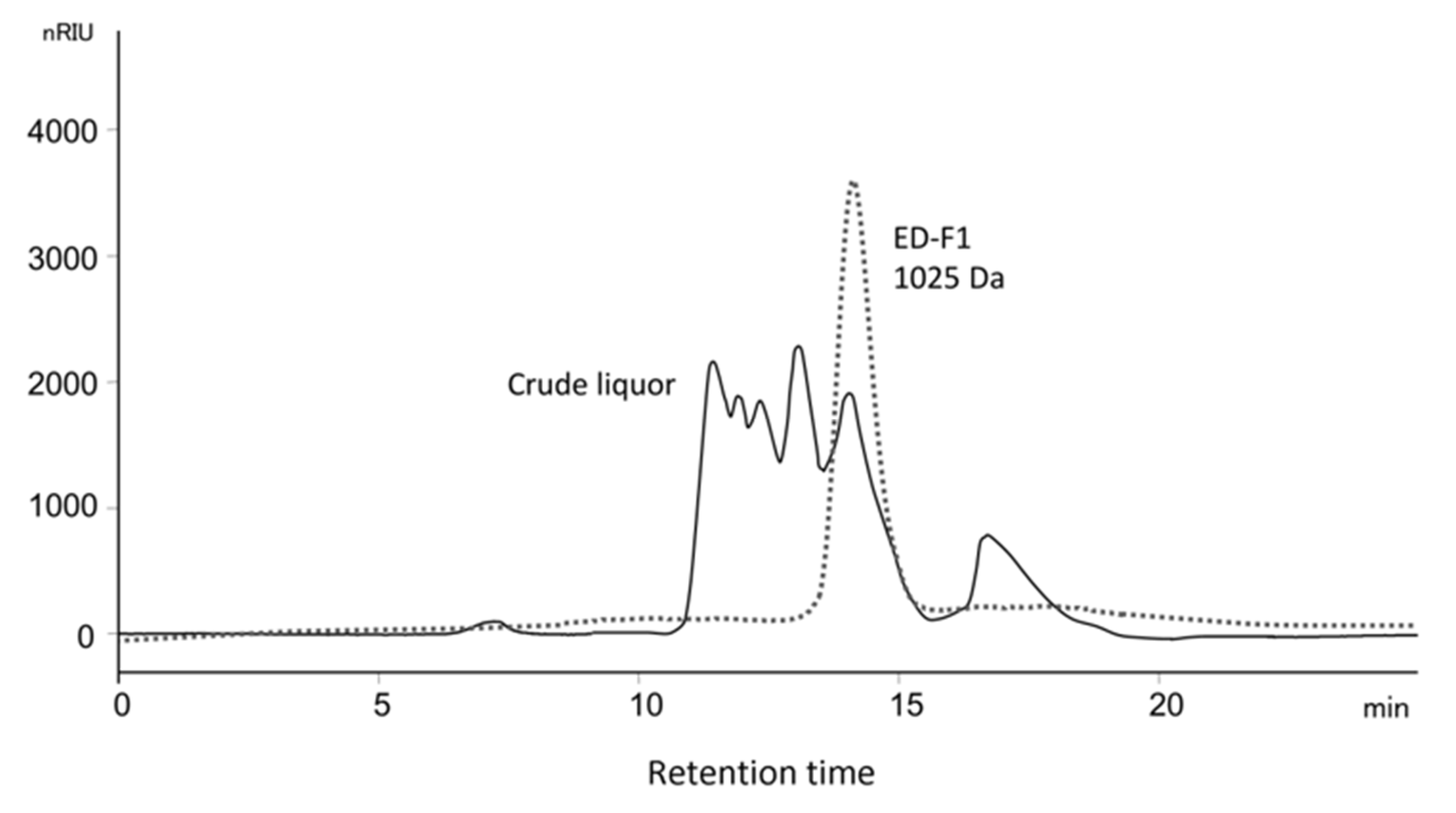
3.6.1. Size-Exclusion Chromatography
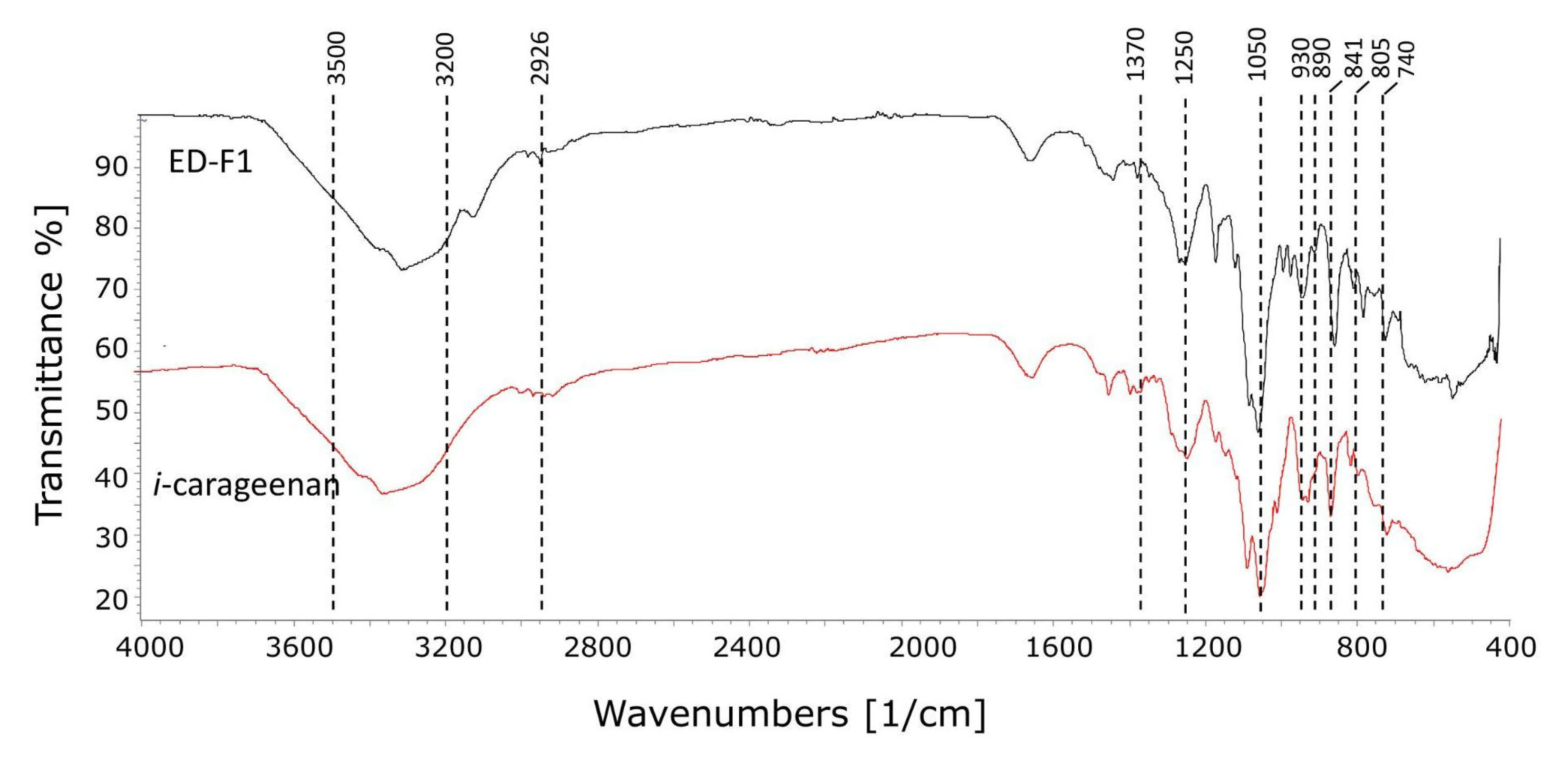
3.6.2. Fourier Transform Infrared Spectra Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy; Available online: https://www.fao.org/3/cc0461en/cc0461en.pdf (accessed on 4 July 2022).
- Chopin, T.; Tacon, A.G.J. Importance of seaweeds and extractive species in global aquaculture production. Rev. Fish. Sci. Aquac. 2021, 29, 139–148. [Google Scholar] [CrossRef]
- Gómez-Guzmán, M.; Rodríguez-Nogales, A.; Algieri, F.; Gálvez, J. Potential role of seaweed polyphenols in cardiovascular-associated disorders. Mar. Drugs 2018, 16, 250. [Google Scholar] [CrossRef]
- Shimoda, H. Seaweed fucoxanthin supplementation improves obesity parameters in mildlyobese Japanese subjects. J. Funct. Food Health Dis. 2017, 7, 246–262. [Google Scholar] [CrossRef]
- Lopes-Costa, E.; Abreu, M.; Gargiulo, D.; Rocha, E.; Ramos, A.A. Anticancer effects of seaweed compounds fucoxanthin and phloroglucinol, alone and in combination with 5-fluorouracil in colon cells. J. Toxicol. Environ. Health 2017, 80, 776–787. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Zhang, X.; Liu, J.; Song, J.; Yu, P.; Chen, P.; Lio, Z.; Wu, M.; Tong, H. Physicochemical characterization of Sargassum fusiforme fucoidan fractions and their antagonistic effect against P-selectin-mediated cell adhesion. Int. J. Biol. Macromol. 2019, 15, 656–662. [Google Scholar] [CrossRef] [PubMed]
- Charoensiddhi, S.; Abraham, R.E.; Su, P.; Zhang, W. Chapter four-seaweed and seaweed-derived metabolites as prebiotics. In Advances in Food and Nutrition Research; Toldrá, F., Ed.; Academic Press: Cambridge, MA, USA, 2020; pp. 97–156. [Google Scholar] [CrossRef]
- Cherry, P.; Yadav, S.; Strain, C.R.; Allsopp, P.J.; McSorley, E.M.; Ross, P.; Stanton, C. Prebiotics from seaweeds: An ocean of opportunity? Mar. Drugs 2019, 17, 327. [Google Scholar] [CrossRef] [PubMed]
- Porse, H.; Rudolph, B. The seaweed hydrocolloid industry: 2016 updates, requirements, and outlook. J. Appl. Phycol. 2017, 29, 2187–2200. [Google Scholar] [CrossRef]
- Zhu, B.; Ni, F.; Xiong, Q.; Yao, Z. Marine oligosaccharides originated from seaweeds: Source, preparation, structure, physiological activity and applications. Crit. Rev. Food Sci. Nutr. 2020, 61, 60–74. [Google Scholar] [CrossRef]
- Seong, H.; Bae, J.-H.; Seo, J.S.; Kim, S.-A.; Kim, T.-J.; Han, N.S. Comparative analysis of prebiotic effects of seaweed polysaccharides laminaran, porphyran, and ulvan using in vitro human fecal fermentation. J. Funct. Foods 2019, 57, 408–416. [Google Scholar] [CrossRef]
- Antonio, M.S.; Andrea, S.; Nunziacarla, S.; Valbona, A.; Marilena, S.; Gioele, C. Gracilaria gracilis, source of agar: A short review. Curr. Org. Chem. 2017, 21, 380–386. [Google Scholar]
- Bajury, D.M.; Rawi, M.H.; Sazali, I.H.; Abdullah, A.; Sarbini, S.R. Prebiotic evaluation of red seaweed (Kappaphycus alvarezii) using in vitro colon model. Int. J. Food Sci. Nutr. 2017, 68, 821–828. [Google Scholar] [CrossRef]
- Chen, X.; Sun, Y.; Hu, L.; Liu, S.; Yu, H.; Xing, R.; Li, P. In vitro prebiotic effects of seaweed polysaccharides. J. Oceanol. Limnol. 2018, 36, 926–932. [Google Scholar] [CrossRef]
- Yang, C.; Lai, S.; Chen, Y.; Liu, D.; Liu, B.; Ai, C.; Wan, X.; Gao, L.; Chen, X.; Zhao, C. Anti-diabetic effect of oligosaccharides from seaweed Sargassum confusum via JNK-IRS1/PI3K signalling pathways and regulation of gut microbiota. Food Chem. Toxicol. 2019, 131, 110562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Aweya, J.J.; Huang, Z.-X.; Kang, Z.-Y.; Bai, Z.-H.; Li, K.-H.; He, X.-T.; Liu, Y.; Chen, X.-Q.; Cheong, K.-L. In vitro fermentation of Gracilaria lemaneiformis sulfated polysaccharides and its agaro-oligosaccharides by human fecal inocula and its impact on microbiota. Carbohydr. Polym. 2020, 234, 115894. [Google Scholar] [CrossRef] [PubMed]
- Trincone, A. Short bioactive marine oligosaccharides: Diving into recent literature. Curr. Biotechnol. 2015, 4, 212–222. [Google Scholar] [CrossRef]
- Klinchongkon, K.; Khuwijitjaru, P.; Wiboonsirikul, J.; Adachi, S. Extraction of oligosaccharides from passion fruit peel by subcritical water treatment. J. Food Process Eng. 2017, 40, e12269. [Google Scholar] [CrossRef]
- You, Y.; Zhang, X.; Li, P.; Lei, F.; Jiang, J. Co-production of xylooligosaccharides and activated carbons from Camellia oleifera shell treated by the catalysis and activation of zinc chloride. Bioresour. Technol. 2020, 306, 123131. [Google Scholar] [CrossRef]
- Chen, Z.; Shen, N.; Wu, X.; Jia, J.; Wu, Y.; Chiba, H.; Hui, S. Extraction and quantitation of phytosterols from edible brown seaweeds: Optimization, validation, and application. Foods 2023, 12, 244. [Google Scholar] [CrossRef]
- Hosni, S.; Gani, S.S.A.; Orsat, V.; Hassan, M.; Abdullah, S. Ultrasound-assisted extraction of antioxidants from Melastoma malabathricum Linn, modeling and optimization using Box–Behnken design. Molecules 2022, 28, 487. [Google Scholar] [CrossRef]
- Zou, X.-G.; Chi, Y.; Cao, Y.-Q.; Zheng, M.; Deng, Z.-Y.; Cai, M.; Yang, K.; Sun, P.-L. Preparation process optimization of peptides from Agaricus blazei Murrill, and comparison of their antioxidant and immune enhancing activities separated by ultrafiltration membrane technology. Foods 2023, 12, 251. [Google Scholar] [CrossRef]
- Meinita, M.D.; Hong, Y.K.; Jeong, G.T. Comparison of sulfuric and hydrochloric acids as catalysts in hydrolysis of Kappaphycus alvarezii (cottonii). Bioprocess Biosyst. Eng. 2011, 35, 123–128. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, Z.; Chen, Y.; Zhenteng Cui, Z.; Liangen Shi, L. Structural elucidation and in vitro antitumor activity of a novel oligosaccharide from Bombyx batryticatus. Carbohydr. Polym. 2014, 103, 434–441. [Google Scholar] [CrossRef] [PubMed]
- Masuko, T.; Minami, A.; Iwasaki, N.; Majima, T.; Nishimura, S.; Lee, Y.C. Carbohydrate analysis by a phenol–sulfuric acid method in microplate format. Anal. Biochem. 2005, 339, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Moniz, P.; Ho, A.L.; Duarte, L.C.; Kolida, S.; Rastall, R.A.; Pereira, H.; Carvalheiro, F. Assessment of the bifidogenic effect of substituted xylo-oligosaccharides obtained from corn straw. Carbohydr. Polym. 2016, 136, 466–473. [Google Scholar] [CrossRef]
- Huebner, J.; Wehling, R.L.; Hutkins, R.W. Functional activity of commercial prebiotics. Int. Dairy J. 2007, 17, 770–775. [Google Scholar] [CrossRef]
- De Andrade, J.K.; Komatsu, E.; Perreault, H.; Torres, Y.R.; Da Rosa, M.R.; Felsner, M.L. In house validation from direct determination of 5-hydroxymethyl-2-furfural (HMF) in Brazilian corn and cane syrups samples by HPLC–UV. Food Chem. 2016, 190, 481–486. [Google Scholar] [CrossRef] [PubMed]
- Altemimi, A.B.; Mohammed, M.J.; Yi-Chen, L.; Watson, D.G.; Lakhssassi, N.; Cacciola, F.; Ibrahim, S.A. Optimization of ultrasonicated kaempferol extraction from Ocimum basilicum using a box–Behnken design and its densitometric validation. Foods 2020, 9, 1379. [Google Scholar] [CrossRef]
- Sellimi, S.; Younes, I.; Ayed, H.B.; Maalej, H.; Montero, V.; Rinaudo, M.; Dahia, M.; Mechichi, T.; Hajji, M.; Nasri, M. Structural, physicochemical and antioxidant properties of sodium alginate isolated from a Tunisian brown seaweed. Int. J. Biol. Macromol. 2015, 72, 1358–1367. [Google Scholar] [CrossRef]
- Silchenko, A.S.; Rasin, A.B.; Kusaykin, M.I.; Kalinovsky, A.I.; Miansong, Z.; Changheng, L.; Malyarenko, O.; Zueva, A.O.; Zvyagintseva, T.N.; Ermakova, S.P. Structure, enzymatic transformation, anticancer activity of fucoidan and sulphated fucooligosaccharides from Sargassum horneri. Carbohydr. Polym. 2017, 175, 654–660. [Google Scholar] [CrossRef]
- Fournière, M.; Latire, T.; Lang, M.; Terme, N.; Bourgougnon, N.; Bedoux, G. Production of active poly- and oligosaccharidic fractions from Ulva sp. by combining enzyme-assisted extraction (eae) and depolymerization. Metabolites 2019, 9, 182. [Google Scholar] [CrossRef] [PubMed]
- Duong-Ly, K.C.; Gabelli, S.B. Salting out of proteins using ammonium sulfate precipitation. Methods Enzymol. 2014, 541, 85–94. [Google Scholar]
- Cosenza, V.A.; Navarro, D.A.; Stortz, C.A.; Ana, M.; Rojas, A.M. Rheology of partially and totally oxidized red seaweed galactans. Carbohydr. Polym. 2020, 230, 115653. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Hu, H.; Wang, L.; Liu, F.; Pan, S. Preparation and prebiotic potential of pectin oligosaccharides obtained from citrus peel pectin. Food Chem. 2018, 244, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Praveen, M.; Karthika Parvathy, K.R.; Jayabalan, R.; Balasubramanian, P. Dietary fiber from Indian edible seaweeds and its in-vitro prebiotic effect on the gut microbiota. Food Hydrocoll. 2019, 96, 343–353. [Google Scholar] [CrossRef]
- Fuhren, J.; Schwalbe, M.; Peralta-Marzal, L.; Rosch, C.; Schols, H.A.; Kleerebezem, W. Phenotypic and genetic characterization of differential galacto-oligosaccharide utilization in Lactobacillus plantarum. Sci. Rep. 2020, 10, 21657. [Google Scholar] [CrossRef]
- Singh, B.P.; Vij, S. α-Galactosidase activity and oligosaccharides reduction pattern of indigenous lactobacilli during fermentation of soy milk. Food Biosci. 2018, 22, 32–37. [Google Scholar] [CrossRef]
- Zúñiga, M.; Yebra, M.J.; Monedero, V. Complex oligosaccharide utilization pathways in lactobacillus. Curr. Issues Mol. Biol. 2021, 40, 49–80. [Google Scholar] [CrossRef]
- Yoo, H.-U.; Ko, M.-J.; Chung, M.-S. Hydrolysis of beta-glucan in oat flour during subcritical-water extraction. Food Chem. 2020, 308, 125670. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.; Mao, H.; Wang, P.; Zuo, Z.; Gao, L.; Shi, X.; Yin, R.; Gao, N.; Zhao, J. Characterization of the hydrolysis kinetics of fucosylated glycosaminoglycan in mild acid and structures of the resulting oligosaccharides. Mar. Drugs 2020, 18, 286. [Google Scholar] [CrossRef]
- Sophonputtanaphoca, S.; Pridam, C.; Chinnak, J.; Nathong, M.; Juntipwong, P. Production of non-digestible oligosaccharides as value-added by-products from rice straw. Agric. Nat. Resour. 2018, 52, 169–175. [Google Scholar] [CrossRef]
- Wang, T.; Li, C.; Song, M.; Fan, R. Xylo-oligosaccharides preparation through acid hydrolysis of hemicelluloses isolated from press-lye. Grain Oil Sci. Technol. 2019, 2, 73–77. [Google Scholar] [CrossRef]
- Lin, Q.; Li, H.; Ren, J.; Deng, A.; Li, W.; Liu, C.; Sun, R. Production of xylooligosaccharides by microwave-induced, organic acid-catalyzed hydrolysis of different xylan-type hemicelluloses: Optimization by response surface methodology. Carbohydr. Polym. 2017, 157, 214–225. [Google Scholar] [CrossRef] [PubMed]
- Rungruangsaphakun, J.; Keawsompong, S. Optimization of hydrolysis conditions for the mannooligosaccharides copra meal hydrolysate production. 3 Biotech 2018, 8, 169. [Google Scholar] [CrossRef]
- Khodaei, N.; Karboune, S. Optimization of enzymatic production of prebiotic galacto/galacto(arabino)-oligosaccharides and oligomers from potato rhamnogalacturonan I. Carbohydr. Polym. 2018, 181, 1153–1159. [Google Scholar] [CrossRef]
- Xu, Y.; Shen, M.; Chen, Y.; Lou, Y.; Luo, R.; Chen, J.; Zhang, Y.; Li, J.; Wang, W. Optimization of the polysaccharide hydrolysate from Auricularia auricula with antioxidant activity by response surface methodology. Int. J. Biol. Macromol. 2018, 113, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hu, C.; Zhang, H.; Chen, W.; Deng, Q.; Tang, H.; Huang, F. Optimation for preparation of oligosaccharides from flaxseed gum and evaluation of antioxidant and antitumor activities in vitro. Int. J. Biol. Macromol. 2020, 153, 1107–1116. [Google Scholar] [CrossRef] [PubMed]
- Danish, M.; Khanday, W.A.; Hashim, R.; Sulaiman, N.S.B.; Akhtar, M.N.; Nizami, M. Application of optimized large surface area date stone (Phoenix dactylifera) activated carbon for rhodamin B removal from aqueous solution: Box-Behnken design approach. Ecotoxicol. Environ. Saf. 2017, 139, 280–290. [Google Scholar] [CrossRef]
- Manmai, N.; Unpaprom, Y.; Ramaraj, R. Bioethanol production from sunflower stalk: Application of chemical and biological pretreatments by response surface methodology (RSM). Biomass Convers. Biorefin. 2021, 11, 1759–1773. [Google Scholar] [CrossRef]
- Savic-Gajic, I.M.; Savic, I.M.; Nikolic, V.D. Modelling and optimization of quercetin extraction and biological activity of quercetin-rich red onion skin extract from Southeastern Serbia. J. Food Nutr. Res. 2018, 57, 15–26. [Google Scholar]
- Fawzy, M.A.; Gomaa, M. Optimization of citric acid treatment for the sequential extraction of fucoidan and alginate from Sargassum latifolium and their potential antioxidant and Fe(III) chelation properties. J. Appl. Phycol. 2021, 33, 2523–2535. [Google Scholar] [CrossRef]
- Guo, Z.; Zhao, B.; Li, H.; Miao, S.; Zheng, B. Optimization of ultrasound-microwave synergistic extraction of prebiotic oligosaccharides from sweet potatoes (Ipomoea batatas L.). Innov. Food Sci. Emerg. Technol. 2019, 54, 51–63. [Google Scholar] [CrossRef]
- Prasad, S.; Malav, M.K.; Kumar, S.; Singh, A.; Pant, D.; Radhakrishnan, S. Enhancement of bio-ethanol production potential of wheat straw by reducing furfural and 5-hydroxymethylfurfural (HMF). Bioresour. Technol. Rep. 2018, 4, 50–56. [Google Scholar] [CrossRef]
- Beckendorff, A.; Lamp, A.; Kaltschmitt, M. Optimization of hydrolysis conditions for xylans and straw hydrolysates by HPLC analysis. Biomass Convers. Biorefin. 2021. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, X.; Zhang, R.; Xiao, L.; Yuan, T.; Shi, Q.; Sun, R. Evaluation of xylooligosaccharide production from residual hemicelluloses of dissolving pulp by acid and enzymatic hydrolysis. RSC Adv. 2018, 8, 35211–35217. [Google Scholar] [CrossRef] [PubMed]
- Ebikade, E.; Lym, J.; Wittreich, G.; Saha, B.; Vlachos, D.G. Kinetic studies of acid hydrolysis of food waste-derived saccharides. Ind. Eng. Chem. Res. 2018, 57, 17365–17374. [Google Scholar] [CrossRef]
- He, B.-L.; Zheng, Q.-W.; Guo, L.-Q.; Huang, J.-Y.; Yun, F.; Huang, S.-S.; Lin, J.-F. Structural characterization and immune-enhancing activity of a novel high-molecular-weight polysaccharide from Cordyceps militaris. Int. J. Biol. Macromol. 2020, 145, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Wei, X.; Fu, X.; Xioa, M.; Liu, Z.; Zhang, L.; Mou, H. Dietary galactosyl and mannosyl carbohydrates: In-vitro assessment of prebiotic effects. Food Chem. 2020, 329, 127179. [Google Scholar] [CrossRef]
- Han, Z.-L.; Yang, M.; Fu, X.-D.; Chen, M.; Su, Q.; Zhao, Y.-H.; Mou, H.-J. Evaluation of prebiotic potential of three marine algae oligosaccharides from enzymatic hydrolysis. Mar. Drugs 2019, 17, 173. [Google Scholar] [CrossRef] [PubMed]
- Kang, O.L.; Yong, P.F.; Ma’aruf, A.G.; Osman, H.; Nazaruddin, R. Physicochemical and antioxidant studies on oven-dried, freeze-dried and spray-dried agaro-oligosaccharide powders. Int. Food Res. J. 2014, 21, 2363–2367. [Google Scholar]
- Fernando, I.; Sanjeewa, K.; Samarakoon, K.W.; Lee, W.W.; Kim, H.S.; Kang, N.; Ranasinghe, P.; Lee, H.S.; Jeon, Y.J. A fucoidan fraction purified from Chnoospora minima; a potential inhibitor of LPS-induced inflammatory responses. Int. J. Biol. Macromol. 2017, 104, 1185–1193. [Google Scholar] [CrossRef] [PubMed]
- Duan, F.; Yu, Y.; Liu, Z.; Tian, L.; Mou, H. An effective method for the preparation of carrageenan oligosaccharides directly from Eucheuma cottonii using cellulase and recombinant κ-carrageenase. Algal Res. 2016, 1, 93–99. [Google Scholar] [CrossRef]
- Pereira, G.A.; Arruda, H.S.; Molina, G.; Pastore, G.M. Extraction optimization and profile analysis of oligosaccharides in banana pulp and peel. J. Food Process Preserv. 2017, 42, e13408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Independent Variables 1 | Responses 2 | ||||
|---|---|---|---|---|---|
| Std. Order | X1 Temperature (°C) | X2 Time (Min) | X3 H2SO4 (M) | ED-F1 Yield (g/100 g) | HMF (g/L) |
| 1 | 115 (−1) | 15 (−1) | 0.13(0) | 8.58 | 0.67 |
| 2 | 135 (+1) | 15 (−1) | 0.13(0) | 9.52 | 1.94 |
| 3 | 115 (−1) | 35 (+1) | 0.13(0) | 9.64 | 1.75 |
| 4 | 135 (+1) | 35 (+1) | 0.13(0) | 9.12 | 2.10 |
| 5 | 115 (−1) | 25 (0) | 0.05 (−1) | 7.78 | 0.64 |
| 6 | 135 (+1) | 25 (0) | 0.05 (−1) | 8.64 | 1.28 |
| 7 | 115 (−1) | 25 (0) | 0.2 (+1) | 8.15 | 1.50 |
| 8 | 135 (+1) | 25 (0) | 0.2 (+1) | 7.65 | 2.14 |
| 9 | 125 (0) | 15 (−1) | 0.05 (−1) | 8.41 | 0.73 |
| 10 | 125 (0) | 35 (+1) | 0.05 (−1) | 9.26 | 0.96 |
| 11 | 125 (0) | 15(−1) | 0.2 (+1) | 8.55 | 1.26 |
| 12 | 125 (0) | 35 (+1) | 0.2 (+1) | 8.77 | 2.10 |
| 13 | 125 (0) | 25 (0) | 0.13(0) | 9.86 | 1.36 |
| 14 | 125 (0) | 25 (0) | 0.13 (0) | 10.11 | 1.47 |
| 15 | 125 (0) | 25 (0) | 0.13 (0) | 9.85 | 1.35 |
| 16 | 125 (0) | 25 (0) | 0.13 (0) | 10.05 | 1.51 |
| 17 | 125 (0) | 25 (0) | 0.13 (0) | 9.87 | 1.32 |
| Regression Coefficients 1 | ||||
|---|---|---|---|---|
| Model Parameters | ED-F1 Yield | HMF | ||
| Full Quadratic Model | Reduced Quadratic Model | Full Quadratic Model | Reduced Quadratic Model | |
| Intercept | 9.97 | 9.98 | 5.88 | 5.88 |
| X1-Temperature (°C) | −0.098 * | −0.098 * | 0.72 *** | 0.72 *** |
| X2-Time (Min) | −0.216 *** | −0.216 *** | 0.39 *** | 0.39 *** |
| X3-H2SO4 (M) | −0.121 * | −0.121 * | 1.37 *** | 1.37 *** |
| X1X2 | −0.365 *** | −0.365 *** | 0.23 ** | 0.23 ** |
| X1X3 | −0.34 *** | −0.34 *** | −0.23 | - |
| X2X3 | −0.158 *** | −0.158 *** | −0.0063 * | −0.0063 * |
| X12 | −0.722 * | 0.722 * | 0.90 ** | 0.90 ** |
| X22 | −0.029 | - | −0.043 | - |
| X32 | −1.19 *** | −1.19 *** | −1.05 ** | −1.05 ** |
| Polynomial model F value p value | 98.47 0.0012 | 121.19 <0.0001 | 41.18 <0.0001 | 58.16 <0.0001 |
| Lack of fit F value p value | 0.804 0.553 | 0.6738 0.6443 | 0.472 0.718 | 0.464 0.789 |
| Standard deviation | 0.108 | 0.103 | 0.1 | 0.095 |
| R2 | 0.9922 | 0.9918 | 0.9815 | 0.9784 |
| Mean | 9.05 | 9.05 | 1.41 | 1.41 |
| Adjusted R2 | 0.9821 | 0.9836 | 0.9576 | 0.9616 |
| Coefficient of variation (CV)(%) | 1.19 | 1.14 | 7.10 | 6.77 |
| Predicted R2 | 0.9452 | 0.9607 | 0.901 | 0.926 |
| Adeq precision | 27.522 | 30.3894 | 20.466 | 24.02 |
| Optimum Conditions 1 | Responses 2 | Desirability = 0.844 | ||||
|---|---|---|---|---|---|---|
| Temperature (°C) (In range) | Time (Min) (In range) | H2SO4 (M) (In range) | Predicted | Actual | % Relative error | |
| 121 | 21 | 0.12 | ED-F1 Yield (g/100 g) (Maximize) | 10.81 | 11.15 ±0.03 | 3.15 |
| HMF (g/L) (Minimize) | 1.06 | 1.11 ±0.08 | 4.72 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Padam, B.S.; Siew, C.K.; Chye, F.Y. Optimization of an Innovative Hydrothermal Processing on Prebiotic Properties of Eucheuma denticulatum, a Tropical Red Seaweed. Appl. Sci. 2023, 13, 1517. https://doi.org/10.3390/app13031517
Padam BS, Siew CK, Chye FY. Optimization of an Innovative Hydrothermal Processing on Prebiotic Properties of Eucheuma denticulatum, a Tropical Red Seaweed. Applied Sciences. 2023; 13(3):1517. https://doi.org/10.3390/app13031517
Chicago/Turabian StylePadam, Birdie Scott, Chee Kiong Siew, and Fook Yee Chye. 2023. "Optimization of an Innovative Hydrothermal Processing on Prebiotic Properties of Eucheuma denticulatum, a Tropical Red Seaweed" Applied Sciences 13, no. 3: 1517. https://doi.org/10.3390/app13031517
APA StylePadam, B. S., Siew, C. K., & Chye, F. Y. (2023). Optimization of an Innovative Hydrothermal Processing on Prebiotic Properties of Eucheuma denticulatum, a Tropical Red Seaweed. Applied Sciences, 13(3), 1517. https://doi.org/10.3390/app13031517