

Cytoprotective Effect of Acid-Hydrolyzed Sesame Leaves against Oxidative Stress in HepG2 Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Sample Preparation
2.3. Quantitation of Verbascoside and Caffeic Acid Content
2.4. LC-Q-TOF MS Analysis
2.5. Cell Culture and Cytotoxicity
2.6. Determination of Intracellular Reactive Oxygen Species Generation
2.7. Quantitation of Glutathione and Lipid Peroxidation
2.8. Measurement of Antioxidant Enzyme Activities
2.9. Statistical Analyses
3. Results and Discussion
3.1. Effects of Acid-Hydrolyzed Sesame Leaves on Verbascoside Content
3.2. Identification of Phytochemicals in Acid-Hydrolyzed Sesame Leaves
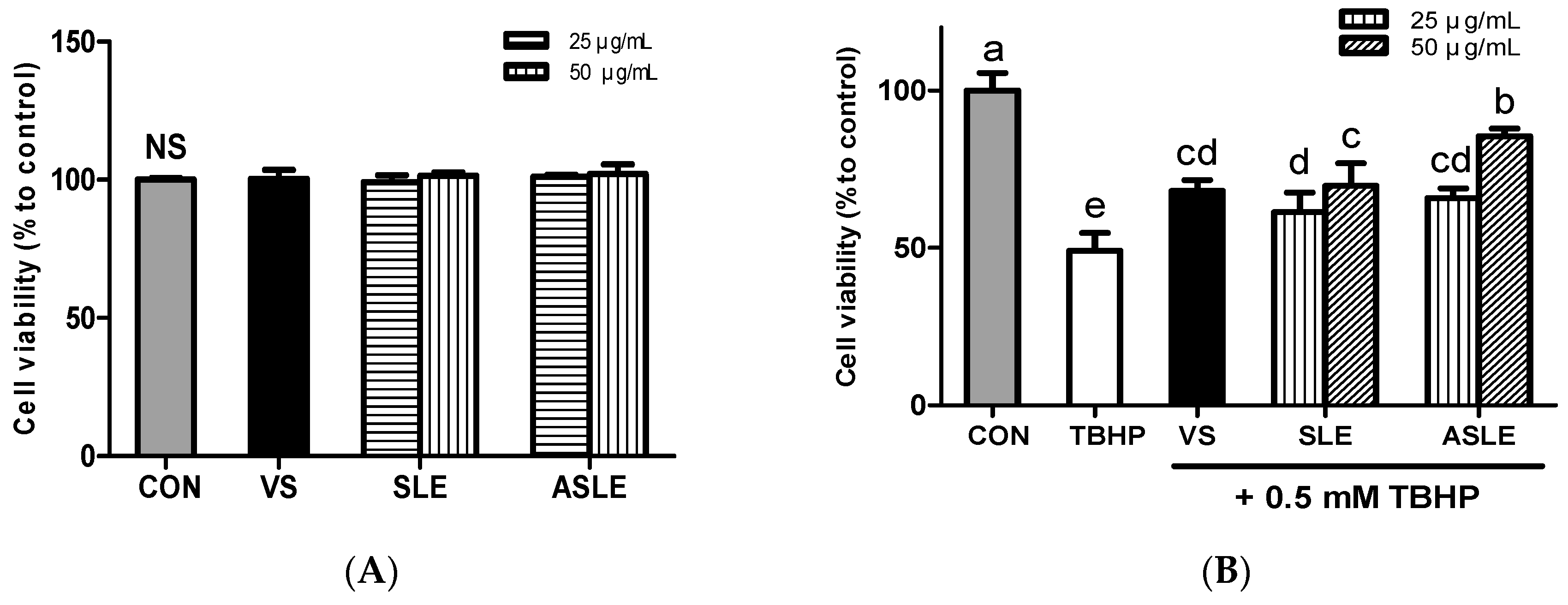
3.3. Effects of Acid-Hydrolyzed Sesame Leaves on Cell Viability and Cytoprotection in HepG2 Cells
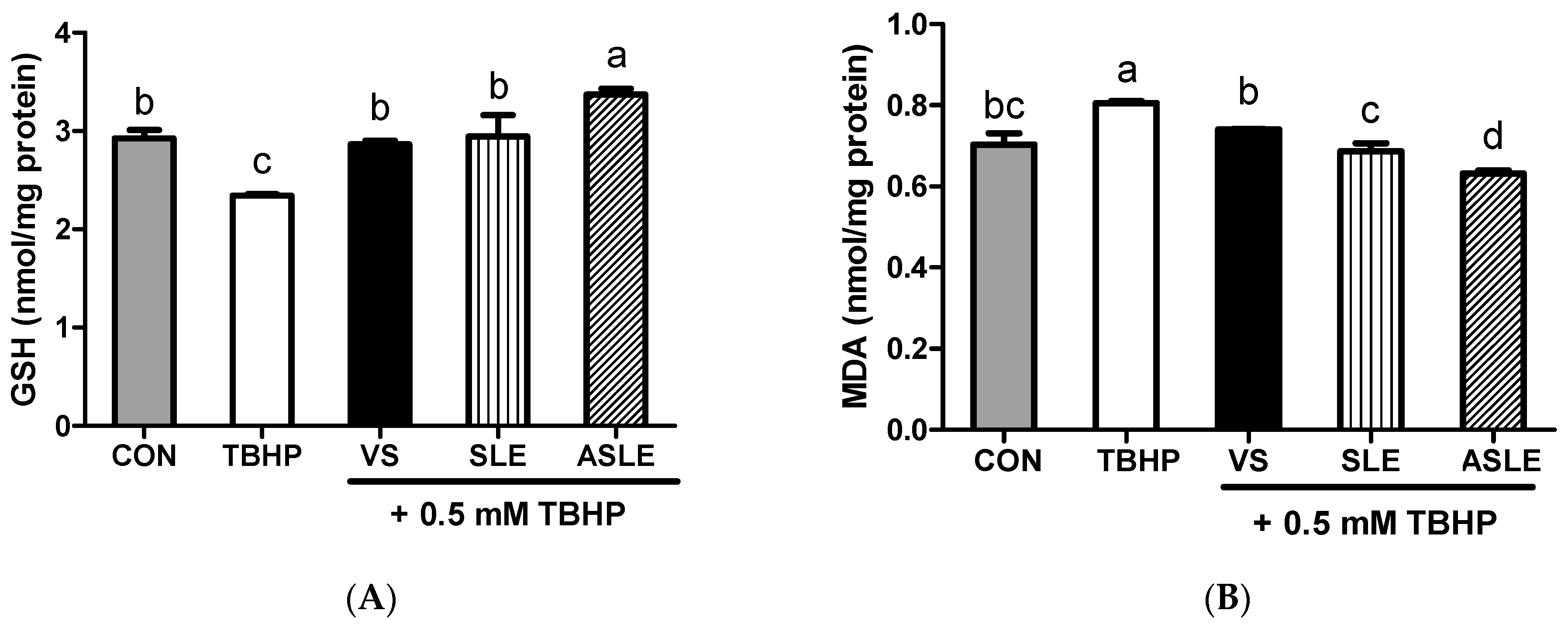
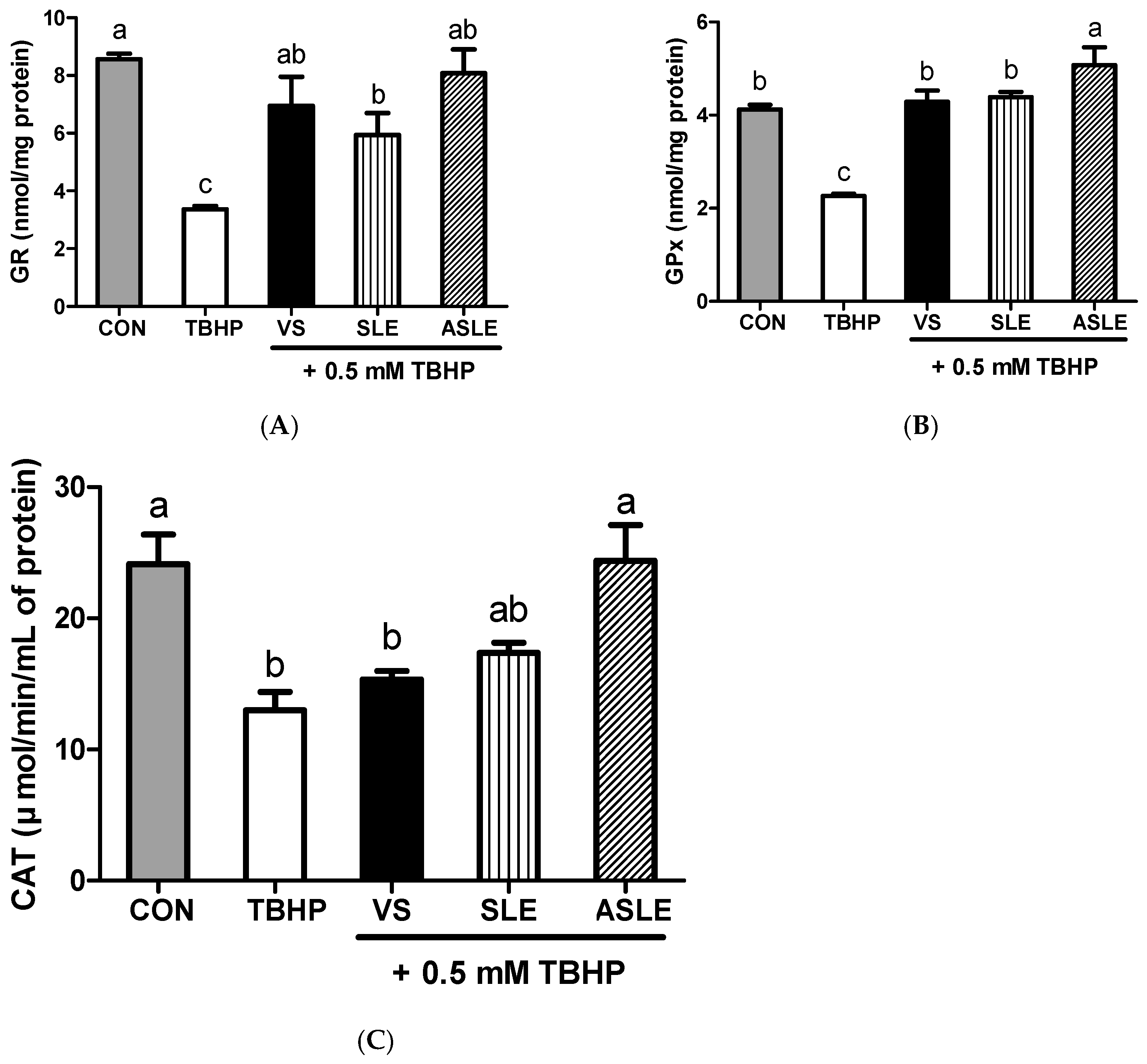
3.4. Intracellular Antioxidative Activities of Acid-Hydrolyzed Sesame Leaves
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ando, K.; Fujita, T. Metabolic syndrome and oxidative stress. Free Radic. Biol. Med. 2009, 47, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen under Control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef] [PubMed]
- Tausz, M.; Sircelj, H.; Grill, D. The glutathione system as a stress marker in plant ecophysiology: Is a stress-response concept valid? J. Exp. Bot. 2004, 55, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Cichoż-Lach, H.; Michalak, A. Oxidative stress as a crucial factor in liver diseases. World J. Gastroenterol. 2014, 20, 8082–8091. [Google Scholar] [CrossRef]
- Li, S.; Tan, H.Y.; Wang, N.; Zhang, S.J.; Lao, L.; Wong, C.W.; Feng, Y. The Role of Oxidative Stress and Antioxidants in Liver Diseases. Int. J. Mol. Sci. 2015, 16, 26087–26124. [Google Scholar] [CrossRef] [PubMed]
- Fuji, Y.; Uchida, A.; Fukahori, K.; Chino, M.; Ohtsuki, T.; Matsufuji, H. Chemical characterization and biological activity in young sesame leaves (Sesamum indicum L.) and changes in iridoid and polyphenol content at different growth stages. PLoS ONE 2018, 13, e0194449. [Google Scholar] [CrossRef]
- Hosseini, M.J.; Shahraki, J.; Tafreshian, S.; Salimi, A.; Kamalinejad, M.; Pourahmad, J. Protective effects of Sesamum indicum extract against oxidative stress induced by vanadium on isolated rat hepatocytes. Environ. Toxicol. 2016, 31, 979–985. [Google Scholar] [CrossRef]
- Fuji, Y.; Ohtsuki, T.; Matsufuji, H. Accumulation and Subcellular Localization of Acteoside in Sesame Plants (Sesamum indicum L.). ACS Omega 2018, 3, 17287–17294. [Google Scholar] [CrossRef]
- Alipieva, K.; Korkina, L.; Orhan, I.E.; Georgiev, M.I. Verbascoside—A review of its occurrence, (bio)synthesis and pharmacological significance. Biotechnol. Adv. 2014, 32, 1065–1076. [Google Scholar] [CrossRef]
- Chou, Y.S.; Ho, Y.L.; Ding, C.W.; Chang, Y.S. New antioxidant phenylethanol glycosides from Torenia concolor. J. Asian Nat. Prod. Res. 2009, 11, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Cardinali, A.; Pati, S.; Minervini, F.; D’Antuono, I.; Linsalata, V.; Lattanzio, V. Verbascoside, isoverbascoside, and their derivatives recovered from olive mill wastewater as possible food antioxidants. J. Agric. Food Chem. 2012, 60, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Nigro, O.; Tuzi, A.; Tartaro, T.; Giaquinto, A.; Vallini, I.; Pinotti, G. Biological effects of verbascoside and its anti-inflammatory activity on oral mucositis: A review of the literature. Anticancer Drugs 2020, 31, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Day, A.J.; Canada, F.J.; Diaz, J.C.; Kroon, P.A.; McLauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett. 2000, 468, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Lee, H.; Sung, J.; Kim, Y.; Jeong, H.S.; Lee, J. Conversion of Rutin to Quercetin by Acid Treatment in Relation to Biological Activities. Prev. Nutr. Food Sci. 2019, 24, 313–320. [Google Scholar] [CrossRef]
- Yang, J.; Lee, J.; Kim, Y. Effect of Deglycosylated Rutin by Acid Hydrolysis on Obesity and Hyperlipidemia in High-Fat Diet-Induced Obese Mice. Nutrients 2020, 12, 1539. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Feng, C.; Hu, L.; Zhao, X.; Tang, X.; Huang, Y.; Luo, J.; Xu, M.; Xie, W. Exploration of a ternary deep eutectic solvent for the efficient extraction of plantamajoside, acteoside, quercetin and kaempferol from Plantago asiatica L. Phytochem. Anal. 2022, 33, 94–104. [Google Scholar] [CrossRef]
- Kim, D.; Kim, Y.; Kim, Y. Effect of Purple Sweet Potato Using Different Cooking Methods on Cytoprotection against Ethanol-Induced Oxidative Damage through Nrf2 Activation in HepG2 Cells. Antioxidants 2023, 12, 1650. [Google Scholar] [CrossRef]
- Görgüç, A.; Özer, P.; Yılmaz, F.M. Microwave-assisted enzymatic extraction of plant protein with antioxidant compounds from the food waste sesame bran: Comparative optimization study and identification of metabolomics using LC/Q-TOF/MS. J. Food Process. Preserv. 2019, 44, 14304. [Google Scholar] [CrossRef]
- Olalere, O.A.; Abdurahman, H.N.; Gan, C.Y. Microwave-enhanced extraction and mass spectrometry fingerprints of polyphenolic constituents in Sesamum indicum leaves. Ind. Crops Prod. 2019, 131, 151–159. [Google Scholar] [CrossRef]
- Kim, Y.; Choi, Y.; Ham, H.; Jeong, H.S.; Lee, J. Protective effects of oligomeric and polymeric procyanidin fractions from defatted grape seeds on tert-butyl hydroperoxide-induced oxidative damage in HepG2 cells. Food Chem. 2013, 137, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.A.; Cerniglia, G.J.; Zaman, A. Microtiter plate assay for the measurement of glutathione and glutathione disulfide in large numbers of biological samples. Anal. Biochem. 1990, 190, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Fossati, P.; Prencipe, L.; Berti, G. Use of 3,5-dichloro-2-hydroxybenzenesulfonic acid/4-aminophenazone chromogenic system in direct enzymic assay of uric acid in serum and urine. Clin. Chem. 1980, 26, 227–231. [Google Scholar] [CrossRef] [PubMed]
- Smith, I.K.; Vierheller, T.L.; Thorne, C.A. Assay of glutathione reductase in crude tissue homogenates using 5,5′-dithiobis(2-nitrobenzoic acid). Anal. Biochem. 1988, 175, 408–413. [Google Scholar] [CrossRef] [PubMed]
- Flohe, L.; Gunzler, W.A. Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Panizzi, L.; Flamini, G.; Cioni, P.L.; Morelli, I. Composition and antimicrobial properties of essential oils of four Mediterranean Lamiaceae. J. Ethnopharmacol. 1993, 39, 167–170. [Google Scholar] [CrossRef]
- Tang, Y.; Zhang, B.; Li, X.; Chen, P.X.; Zhang, H.; Liu, R.; Tsao, R. Bound Phenolics of Quinoa Seeds Released by Acid, Alkaline, and Enzymatic Treatments and Their Antioxidant and α-Glucosidase and Pancreatic Lipase Inhibitory Effects. J. Agric. Food Chem. 2016, 64, 1712–1719. [Google Scholar] [CrossRef]
- Wang, L.; Lin, X.; Zhang, J.; Zhang, W.; Hu, A.; Li, W.; Li, C.; Liu, S. Extraction methods for the releasing of bound phenolics from Rubus idaeus L. leaves and seeds. Ind. Crops Prod. 2019, 135, 1–9. [Google Scholar] [CrossRef]
- Alam, M.; Ahmed, S.; Elasbali, A.M.; Adnan, M.; Alam, S.; Hassan, M.I.; Pasupuleti, V.R. Therapeutic Implications of Caffeic Acid in Cancer and Neurological Diseases. Front. Oncol. 2022, 12, 860508. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Aly, S.M.; Ali, H.; Babiker, A.Y.; Srikar, S.; Khan, A.A. Therapeutic effects of date fruits (Phoenix dactylifera) in the prevention of diseases via modulation of anti-inflammatory, anti-oxidant and anti-tumour activity. Int. J. Clin. Exp. Med. 2014, 7, 483–491. [Google Scholar] [PubMed]
- Tanaka, T.; Kojima, T.; Kawamori, T.; Wang, A.; Suzui, M.; Okamoto, K.; Mori, H. Inhibition of 4-nitroquinoline-1-oxide-induced rat tongue carcinogenesis by the naturally occurring plant phenolics caffeic, ellagic, chlorogenic and ferulic acids. Carcinogenesis 1993, 14, 1321–1325. [Google Scholar] [CrossRef] [PubMed]
- Sul, D.; Kim, H.S.; Lee, D.; Joo, S.S.; Hwang, K.W.; Park, S.Y. Protective effect of caffeic acid against beta-amyloid-induced neurotoxicity by the inhibition of calcium influx and tau phosphorylation. Life Sci. 2009, 84, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Wanasundara, P.K. Phenolic antioxidants. Crit. Rev. Food Sci. Nutr. 1992, 32, 67–103. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Shi, R.; Wang, X.; Shen, H.M. Luteolin, a flavonoid with potential for cancer prevention and therapy. Curr. Cancer Drug Targets 2008, 8, 634–646. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Tian, X.H.; Yi, Y.S.; Jiang, W.S.; Zhou, Y.J.; Cheng, W.J. Luteolin-induced protection of H(2)O(2)-induced apoptosis in PC12 cells and the associated pathway. Mol. Med. Rep. 2015, 12, 7699–7704. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.H.; Yao, X.L.; Zhang, Y.; Zhang, S.F.; Hu, J.C. Luteolin Could Improve Cognitive Dysfunction by Inhibiting Neuroinflammation. Neurochem. Res. 2018, 43, 806–820. [Google Scholar] [CrossRef]
- Hussain, J.; Ali, L.; Khan, A.L.; Rehman, N.U.; Jabeen, F.; Kim, J.S.; Al-Harrasi, A. Isolation and bioactivities of the flavonoids morin and morin-3-O-beta-D-glucopyranoside from Acridocarpus orientalis-A wild Arabian medicinal plant. Molecules 2014, 19, 17763–17772. [Google Scholar] [CrossRef]
- Chagas, M.; Behrens, M.D.; Moragas-Tellis, C.J.; Penedo, G.X.M.; Silva, A.R.; Goncalves-de-Albuquerque, C.F. Flavonols and Flavones as Potential anti-Inflammatory, Antioxidant, and Antibacterial Compounds. Oxidative Med. Cell Longev. 2022, 2022, 9966750. [Google Scholar] [CrossRef]
- Visioli, F.; Bellomo, G.; Montedoro, G.; Galli, C. Low density lipoprotein oxidation is inhibited in vitro by olive oil constituents. Atherosclerosis 1995, 117, 25–32. [Google Scholar] [CrossRef]
- Hamden, K.; Allouche, N.; Damak, M.; Elfeki, A. Hypoglycemic and antioxidant effects of phenolic extracts and purified hydroxytyrosol from olive mill waste in vitro and in rats. Chem. Biol. Interact. 2009, 180, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed]
- Jove, M.; Mota-Martorell, N.; Pradas, I.; Martin-Gari, M.; Ayala, V.; Pamplona, R. The Advanced Lipoxidation End-Product Malondialdehyde-Lysine in Aging and Longevity. Antioxidants 2020, 9, 1132. [Google Scholar] [CrossRef] [PubMed]
- Lu, S.C. Dysregulation of glutathione synthesis in liver disease. Liver Res. 2020, 4, 64–73. [Google Scholar] [CrossRef]
- Hong, S.; Kim, Y.; Sung, J.; Lee, H.; Heo, H.; Jeong, H.S.; Lee, J. Jujube (Ziziphus jujuba Mill.) Protects Hepatocytes against Alcohol-Induced Damage through Nrf2 Activation. Evid. Based Complement. Alternat. Med. 2020, 2020, 6684331. [Google Scholar] [CrossRef]
- Liao, W.; Ning, Z.; Chen, L.; Wei, Q.; Yuan, E.; Yang, J.; Ren, J. Intracellular antioxidant detoxifying effects of diosmetin on 2,2-azobis(2-amidinopropane) dihydrochloride (AAPH)-induced oxidative stress through inhibition of reactive oxygen species generation. J. Agric. Food Chem. 2014, 62, 8648–8654. [Google Scholar] [CrossRef]
- Ji, S.L.; Cao, K.K.; Zhao, X.X.; Kang, N.X.; Zhang, Y.; Xu, Q.M.; Yang, S.L.; Liu, Y.L.; Wang, C. Antioxidant activity of phenylethanoid glycosides on glutamate-induced neurotoxicity. Biosci. Biotechnol. Biochem. 2019, 83, 2016–2026. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | HCl Concentration (M) | Verbascoside g/100 g Sample Dry Weight (%) | Caffeic Acid g/100 g Sample Dry Weight (%) |
|---|---|---|---|
| Sesame leaves | 0 | 3.950 ± 0.015 c | 0.354 ± 0.000 b |
| 0.1 | 4.398 ± 0.010 a | 0.534 ± 0.002 a | |
| 0.5 | 4.262 ± 0.001 b | 0.307 ± 0.001 c |
| Group | Ion Mode | Adduct | RT (min) (1) | m/z Calculate (2) | m/z Observed (3) | Error (ppm) | Fragment Ions (m/z) | Formula | Identification | Found (4) |
|---|---|---|---|---|---|---|---|---|---|---|
| Iridoid | Neg | [M−H]− | 4.5205 | 419.1195 | 419.1193 | −0.477 | 401.10835, 257.0665, 239.0558 | C17H24O12 | Sesamoside | SLE, ASLE |
| Pos | [M+H]+ | 3.6555 | 423.1497 | 423.1499 | 0.466 | 211.05975, 193.0494, 165.0544, 139.03965, 137.05945 | C17H26O12 | Lamalbid | SLE, ASLE | |
| [M+H]+ | 3.131 | 407.1548 | 407.1555 | 1.749 | 209.08055, 191.06965, 177.0546, 149.05945, 121.0509 | C17H26O11 | Shanzhiside methyl ester | SLE, ASLE | ||
| Phenolic compound | Neg | [M−H]− | 13.965 | 315.0510 | 315.0511 | 0.235 | 300.0454, 272.0779, 255.0093, 243.0267 | C16H12O7 | 3-O-Methylquercetin | SLE, ASLE |
| [M−H]− | 12.118 | 623.1981 | 623.1992 | 1.694 | 624.2018, 461.0723, 179.0347, 161.0259 | C29H36O15 | Verbascoside | SLE, ASLE | ||
| [M−H]− | 6.393 | 353.0878 | 353.0880 | 0.549 | 355.0934, 354.0912, 353.088, 352.0755, 351.0719, 192.05965 | C16H18O9 | Chlorogenic acid | SLE, ASLE | ||
| [M−H]− | 5.111 | 487.1457 | 487.1456 | −0.236 | 341.08415, 179.03485 | C21H28O13 | Cistanoside F | SLE, ASLE | ||
| [M−H]− | 15.205 | 299.0561 | 299.0559 | −0.669 | 285.0344 | C16H12O6 | Diosmetin | SLE, ASLE | ||
| [M−H]− | 13.965 | 315.0510 | 315.0511 | 0.235 | 315.05115, 301.0112, 300.02985, 243.02575, 136.93345 | C16H12O7 | Eupafolin | SLE, ASLE | ||
| [M−H]− | 11.269 | 193.0506 | 193.0505 | −0.684 | 178.02265, 149.0454, 135.0255 | C10H10O4 | Ferulic acid | SLE, ASLE | ||
| [M−H]− | 12.974 | 153.0193 | 153.0194 | 0.444 | 154.0226, 153.0194 | C7H6O4 | Gentisic acid | SLE, ASLE | ||
| [M−H]− | 3.1725 | 153.0557 | 153.0557 | −0.444 | 135.0451, 123.0446 | C8H10O3 | Hydroxytyrosol | SLE, ASLE | ||
| [M−H]− | 14.847 | 285.0405 | 285.0401 | −1.270 | 199.03665, 175.03575, 151.04005, 133.01285 | C15H10O6 | Luteolin | SLE, ASLE | ||
| [M−H]− | 12.873 | 301.0354 | 301.0348 | −1.913 | 299.0186, 227.0135, 151.0406 | C15H10O7 | Morin | ASLE | ||
| [M−H]− | 11.868 | 153.0193 | 153.0193 | −0.209 | 109.0318, 108.01955 | C7H12O6 | Protocatechuic acid | SLE, ASLE | ||
| [M−H]− | 2.083 | 191.0561 | 191.0572 | 5.695 | 173.0458, 159.0296, 145.0516 | C7H6O4 | Quinic acid | SLE, ASLE | ||
| [M−H]− | 13.965 | 315.0510 | 315.0511 | 0.235 | 300.04535, 193.0508, 165.038, 121.0298 | C16H12O7 | Rhamnetin | SLE, ASLE | ||
| Pos | [M+H]+ | 15.176 | 301.0707 | 301.0704 | −0.880 | 258.0496, 121.0509 | C16H12O6 | 3-O-Methylkaempferol | ASLE | |
| [M+H]+ | 4.496 | 181.0495 | 181.0494 | −0.746 | 179.05085, 164.04275, 163.0392, 145.035, 135.04395, 117.0709 | C9H8O4 | Caffeic aicd | SLE, ASLE | ||
| [M+H]+ | 12.400 | 325.1434 | 325.1439 | 1.538 | 270.0787, 149.0238 | C20H20O4 | Corylifolinin | SLE, ASLE | ||
| [M+H]+ | 14.239 | 623.1912 | 623.1949 | 5.979 | 486.16945, 485.1654 | C36H30O10 | Dioonflavone | SLE, ASLE | ||
| [M+NH4]+ | 16.478 | 518.1657 | 518.1703 | 8.903 | 441.11505, 399.12525, 357.1337, 315.07535, 181.08605, 165.0651 | C25H24O11 | Epicatechin pentaacetate | SLE, ASLE | ||
| [M+H]+ | 4.513 | 195.0652 | 195.0655 | 1.615 | 177.05495, 145.03525, 117.055, 89.049, 63.01085 | C10H10O4 | Ferulic acid | SLE, ASLE | ||
| [M+H]+ | 16.062 | 271.0601 | 271.0601 | 0.000 | 243.0619, 223.0592, 215.0777, 169.04615, 153.03285, 141.07155 | C15H10O5 | Genistein | SLE, ASLE | ||
| [M+H]+ | 1.758 | 303.0863 | 303.0844 | −6.318 | 305.08415, 304.0873, 177.05545, 153.0363 | C16H14O6 | Hesperetin | SLE, ASLE | ||
| [M+H]+ | 1.242 | 317.0656 | 317.0617 | −12.300 | 302.0328, 274.0419, 219.0219 | C16H12O7 | Isorhamnetin | SLE, ASLE | ||
| [M+H]+ | 3.468 | 287.0550 | 287.0517 | −11.578 | 135.0434, 68.9935 | C15H10O6 | Luteolin | SLE, ASLE | ||
| [M+H]+ | 4.146 | 146.0600 | 146.0596 | −3.012 | 128.0484, 118.0646, 91.05435, 86.0076 | C9H7NO | Oxyquinoline | SLE, ASLE | ||
| [M+H]+ | 6.959 | 139.0390 | 139.0400 | 7.401 | 121.0507, 105.04225, 95.04865, 77.0389 | C7H6O3 | P-Salicylic acid | SLE, ASLE | ||
| [M+H]+ | 13.965 | 317.0656 | 317.0655 | −0.249 | 319.07135, 318.06905 | C16H12O7 | Rhamnetin | SLE, ASLE | ||
| [M+H]+ | 3.514 | 225.0758 | 225.0757 | −0.222 | 208.068, 207.06485, 192.0361, 175.03855, 147.0435, 119.0488 | C11H12O5 | Sinapic acid | SLE, ASLE | ||
| [M+H]+ | 15.446 | 331.0812 | 331.0815 | 0.819 | 333.0893, 332.08375 | C17H14O7 | Tricin | SLE, ASLE |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, H.; Kim, Y.; Kim, Y. Cytoprotective Effect of Acid-Hydrolyzed Sesame Leaves against Oxidative Stress in HepG2 Cells. Appl. Sci. 2023, 13, 12833. https://doi.org/10.3390/app132312833
Shin H, Kim Y, Kim Y. Cytoprotective Effect of Acid-Hydrolyzed Sesame Leaves against Oxidative Stress in HepG2 Cells. Applied Sciences. 2023; 13(23):12833. https://doi.org/10.3390/app132312833
Chicago/Turabian StyleShin, Heeyoon, Yoonjeong Kim, and Younghwa Kim. 2023. "Cytoprotective Effect of Acid-Hydrolyzed Sesame Leaves against Oxidative Stress in HepG2 Cells" Applied Sciences 13, no. 23: 12833. https://doi.org/10.3390/app132312833
APA StyleShin, H., Kim, Y., & Kim, Y. (2023). Cytoprotective Effect of Acid-Hydrolyzed Sesame Leaves against Oxidative Stress in HepG2 Cells. Applied Sciences, 13(23), 12833. https://doi.org/10.3390/app132312833