1. Introduction
Quantitative research on the balance of global greenhouse gases and their control mechanisms has become a focus among global environmental science research programs. Notable programs that are dedicated to addressing this issue include the International Geosphere-Biosphere Program, the World Climate Research Program, the International Human Dimensions Program on Global Environmental Change, and Global Change and Terrestrial Ecosystems. Wetland ecosystems play a crucial role in the global carbon cycle. Among the five major terrestrial ecosystems closely linked to the global carbon cycle—forests, grasslands, croplands, wetlands, and inland waters—wetlands constitute the largest component of the carbon reservoir of the terrestrial biosphere [
1]. Consequently, they play a vital role in the global carbon cycle [
2,
3]. The total global area of wetlands is approximately 5.7 × 10
6 km
2. This accounts for approximately 6% of the Earth’s land area, which is significantly lower than the area covered by forests; however, due to their high productivity and redox potential, wetlands have become crucial sites for biogeochemical processes in the biosphere [
4]. Owing to prolonged or intermittent waterlogging, the anaerobic conditions in wetlands inhibit the decomposition of detritus, leading to the accumulation of organic matter in the soil. As a result, wetlands are carbon sinks that mitigate the rise in atmospheric CO
2 concentrations. According to estimates by the Intergovernmental Panel on Climate Change, global terrestrial ecosystems store approximately 2.48 × 10
6 TgC (1 Tg = 1 × 10
12 g), of which peatlands store around 0.5 × 10
6 TgC. Peatlands alone store twice the carbon content of all forests combined, constituting 20% of the total carbon stock in global terrestrial ecosystems [
5]. This indicates that wetland ecosystems have significant potential to mitigate global warming. Although they have substantial soil carbon reservoirs, they are also significant sources of methane emissions, accounting for more than 20% of the global annual methane emissions each year [
6]. In the context of global warming, as soil moisture decreases and soil oxidation capacity intensifies, the anaerobic conditions in wetlands are mitigated. This leads to a significant acceleration of the decomposition rates of detritus and peat, resulting in an increase in CO
2 emissions, which can create a positive feedback mechanism that exacerbates global warming [
7].
Reeds are widely distributed in various types of salt marshes because of their adaptability and high reproductive capacity. They are extensively cultivated in various countries because of their significant economic and ecological value. Numerous studies [
8,
9,
10,
11] indicate that peatlands and reed wetlands serve as crucial carbon sinks. However, the carbon sink function of wetlands is influenced by various factors, including hydrology, temperature, and atmospheric CO
2 concentrations as well as nutrients like nitrogen (N) and phosphorus (P). Hence, studying variations in carbon fluxes in reed wetlands is important for understanding the sources and sinks of greenhouse gases.
The Momoge salt marshes are located in Jilin Province, China, and are characterized by arid and semi-arid lands with fragile ecosystems. Natural hydrological changes and human disturbances contribute to significant spatiotemporal variability in greenhouse gas emissions. Reed marshes, as one of the widely distributed wetland types in the region, play a crucial role in wetland carbon balances. Currently, due to the limited direct observations of CO2 exchange between reed marshes and the atmosphere, quantitatively analyzing the correlations between CO2 fluxes and environmental factors remains difficult. This limits our understanding of various physical and chemical processes that influence carbon accumulation and cycling in reed marshes. Advancements in eddy covariance technology have made accurate measurements of carbon exchange in wetland ecosystems possible. This study is based on four years of data from eddy correlation flux towers and automated meteorological stations within the study area. By investigating the CO2 fluxes in the Momoge salt marsh ecosystem, this study analyzes the daily, seasonal, and interannual variation in the net ecosystem exchange (NEE). Additionally, by exploring the correlations between net CO2 exchanges and major environmental factors at both daily and monthly scales, the results provide new insights into the local climate characteristics and land–atmosphere interactions in the Momoge wetlands. These findings can provide localized and accurate data for climate change modeling, global ecosystem modeling, and developing strategies for reducing greenhouse gas emissions.
3. Results
3.1. Daily Dynamics of Net CO2 Exchange in the Salt Marsh Ecosystem
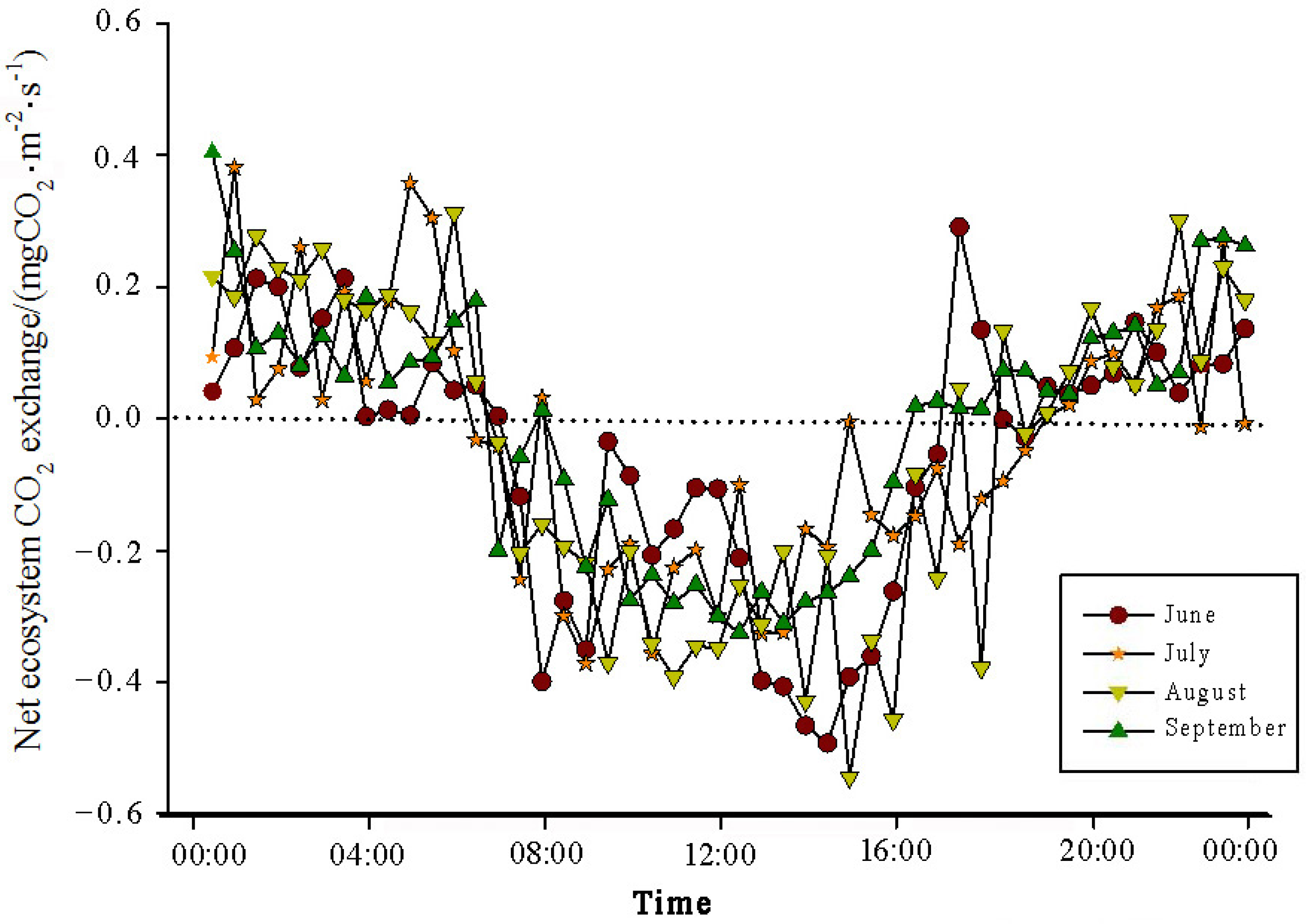
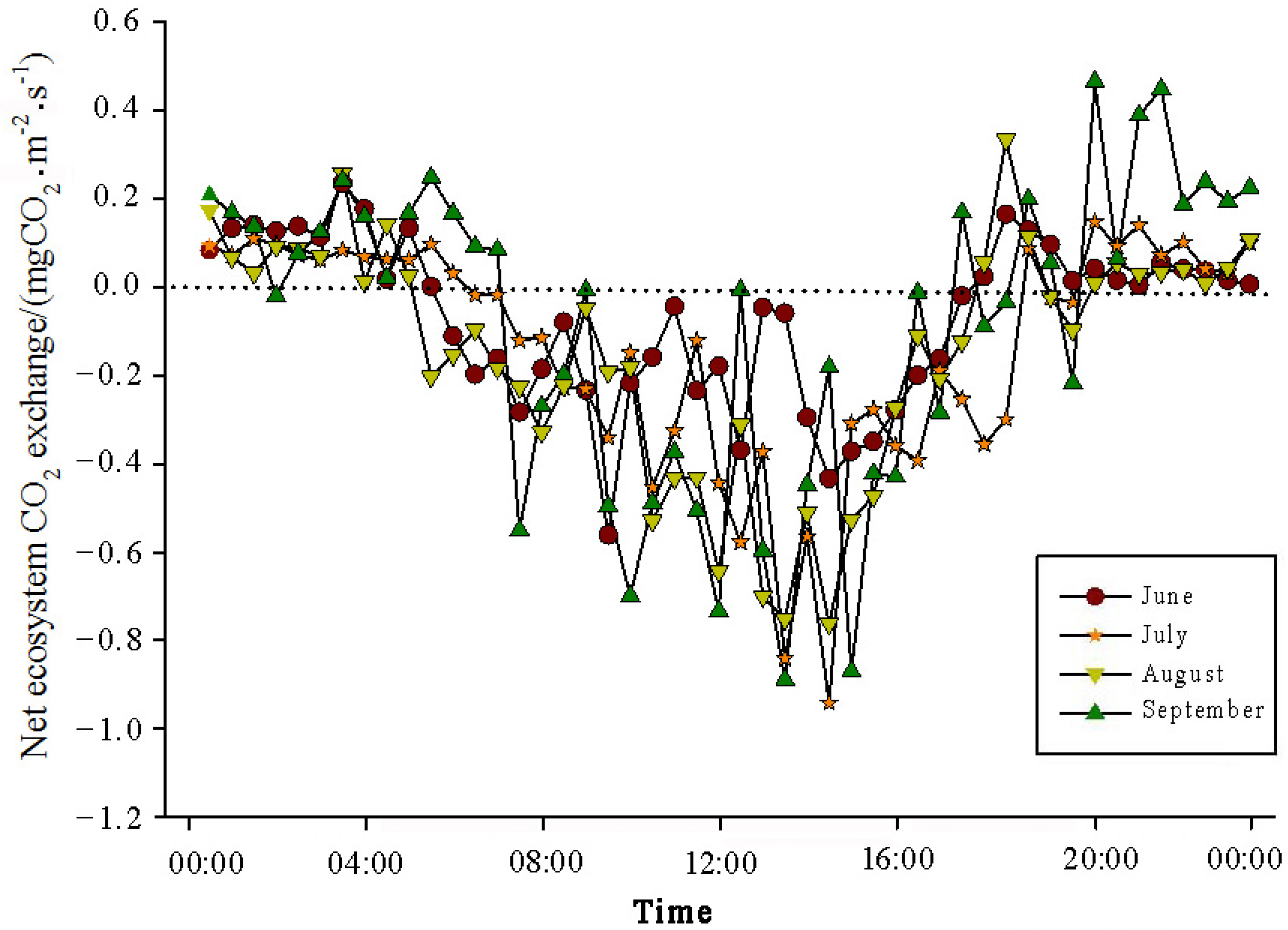
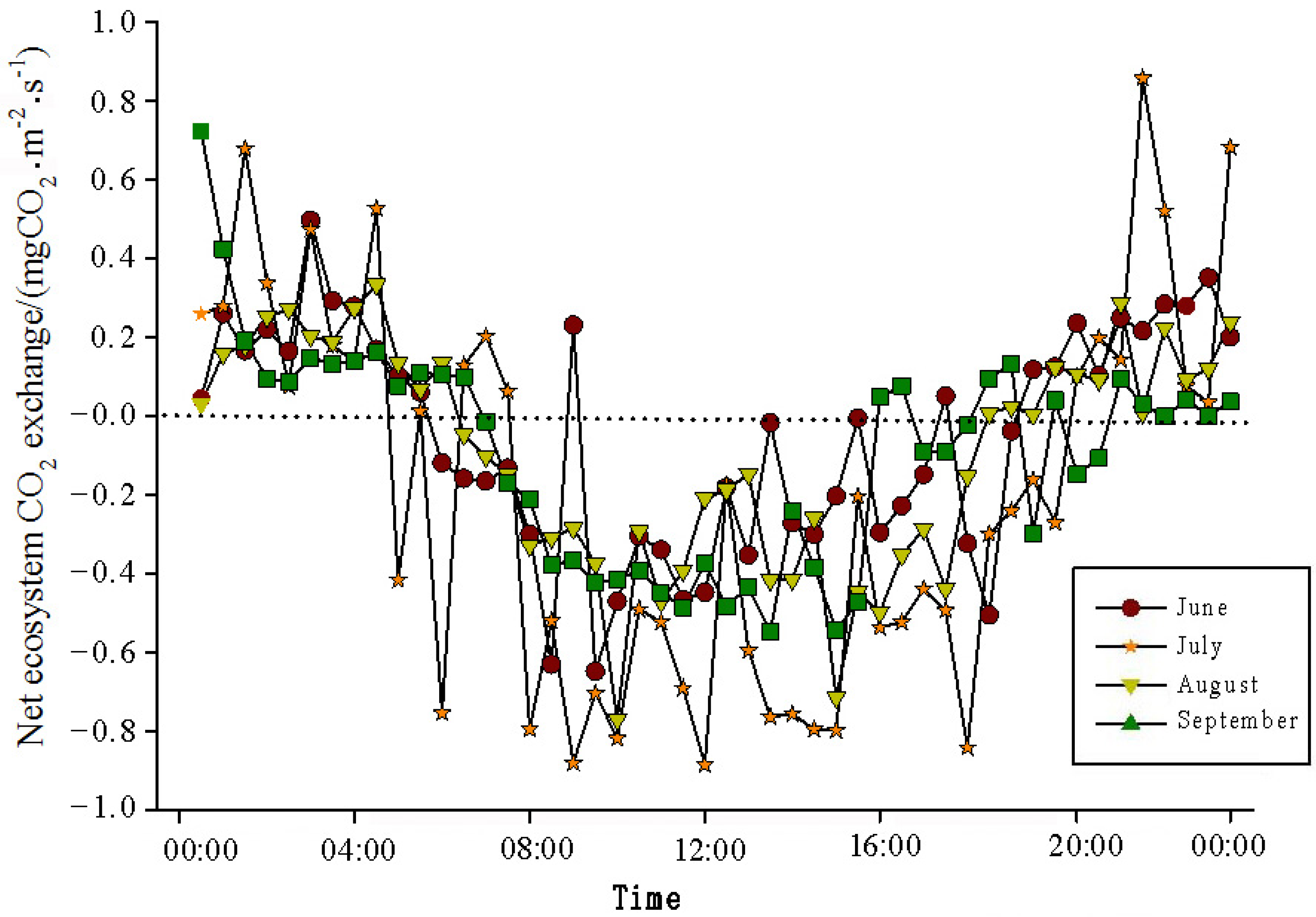
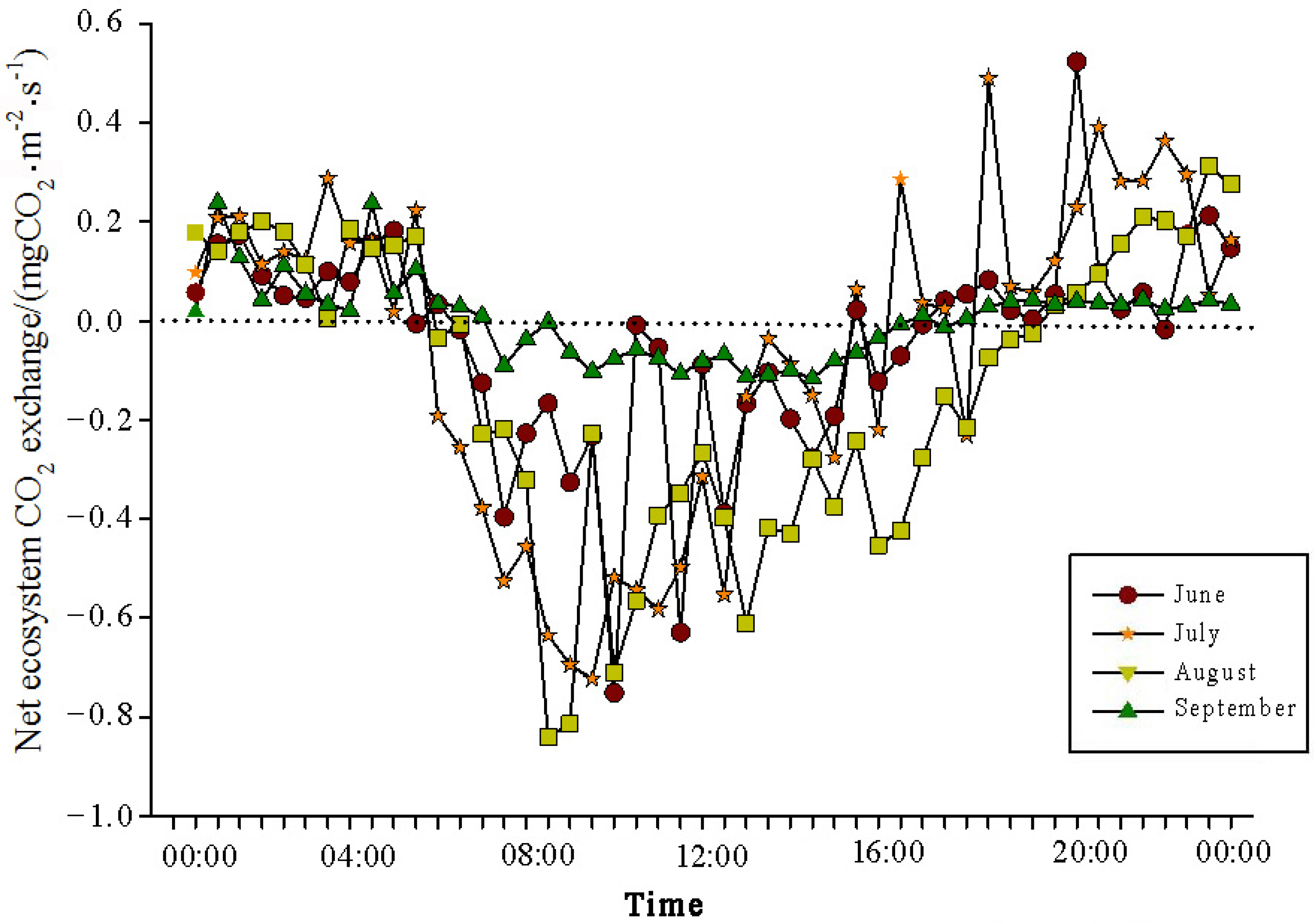
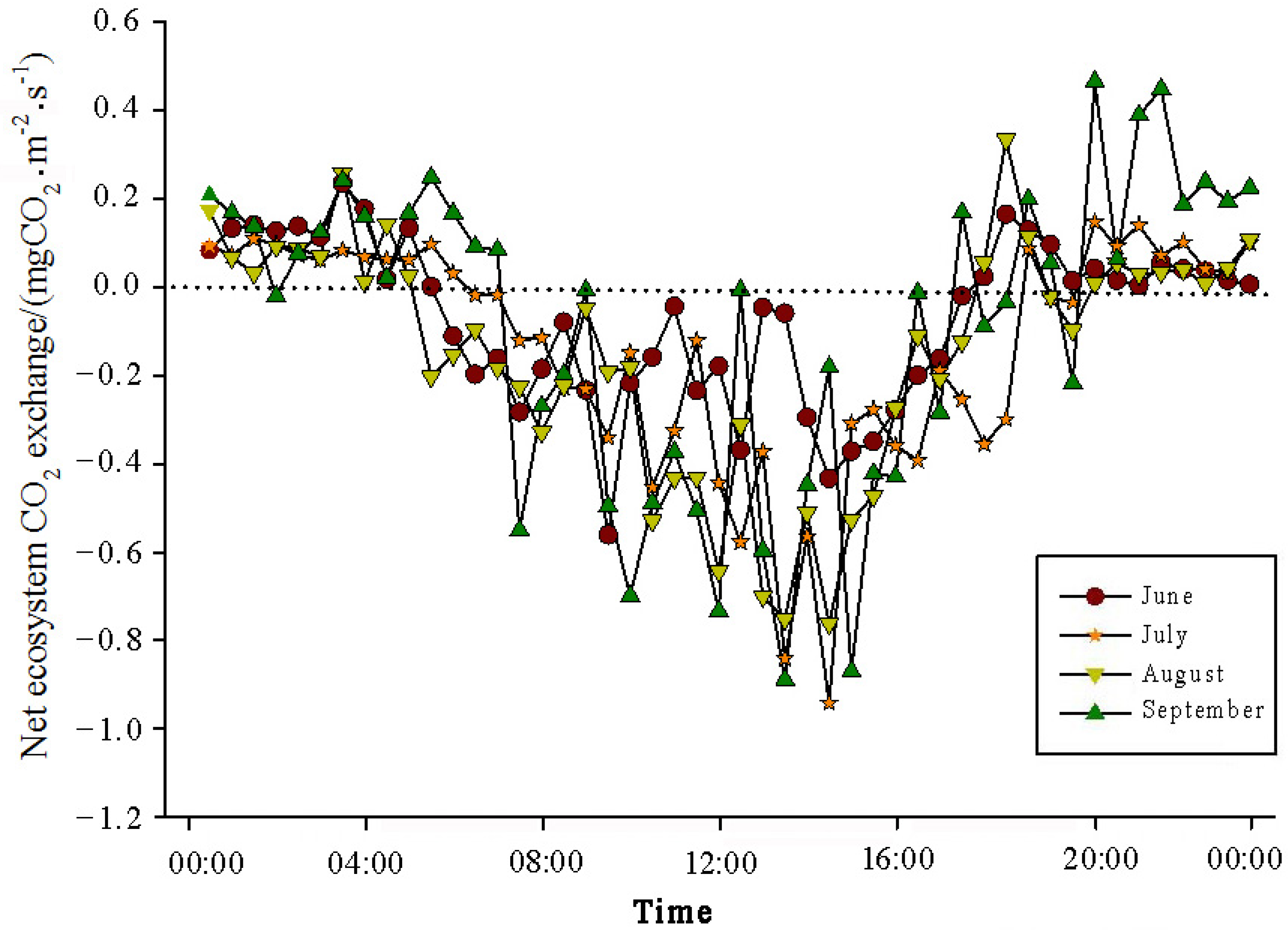
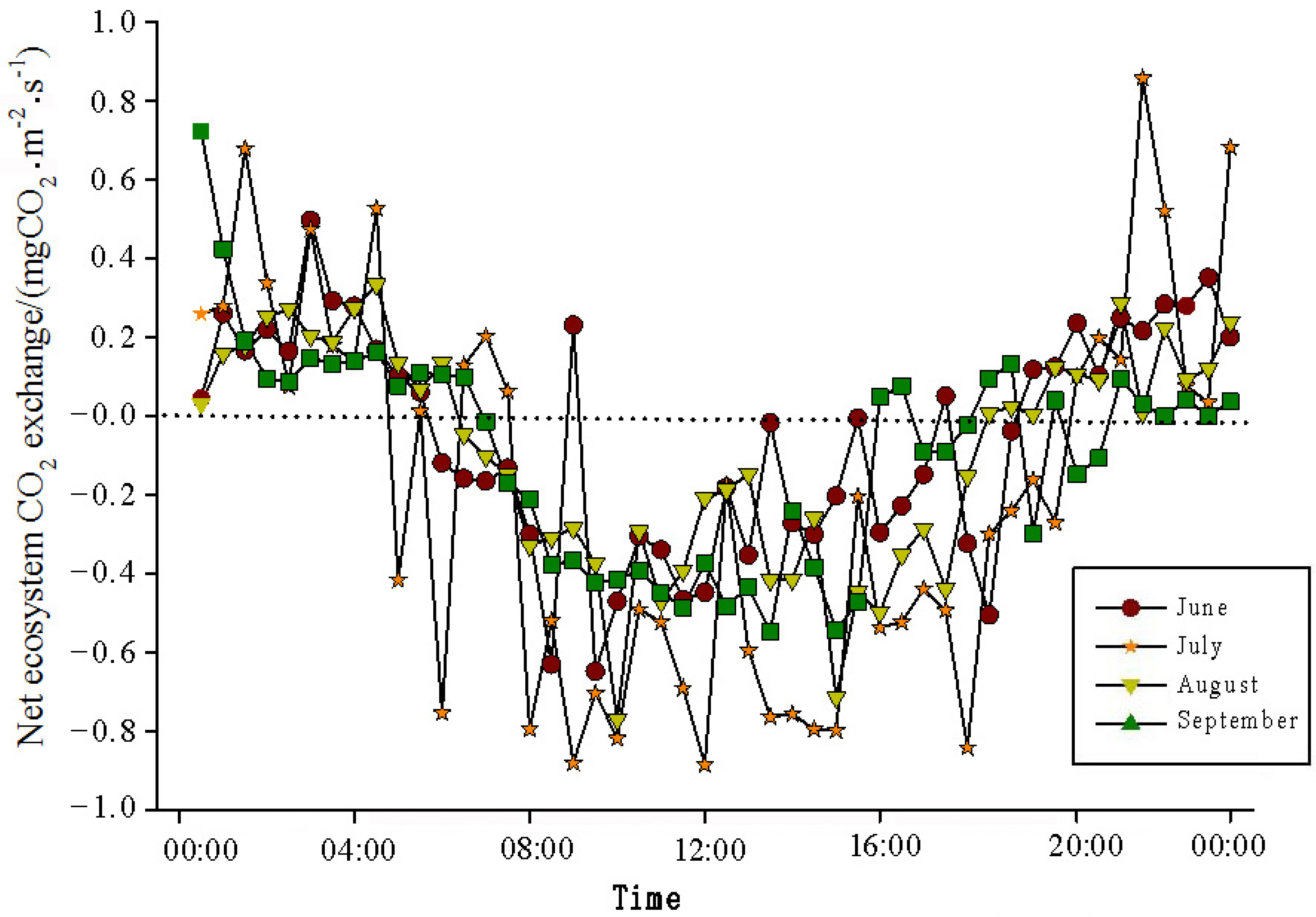
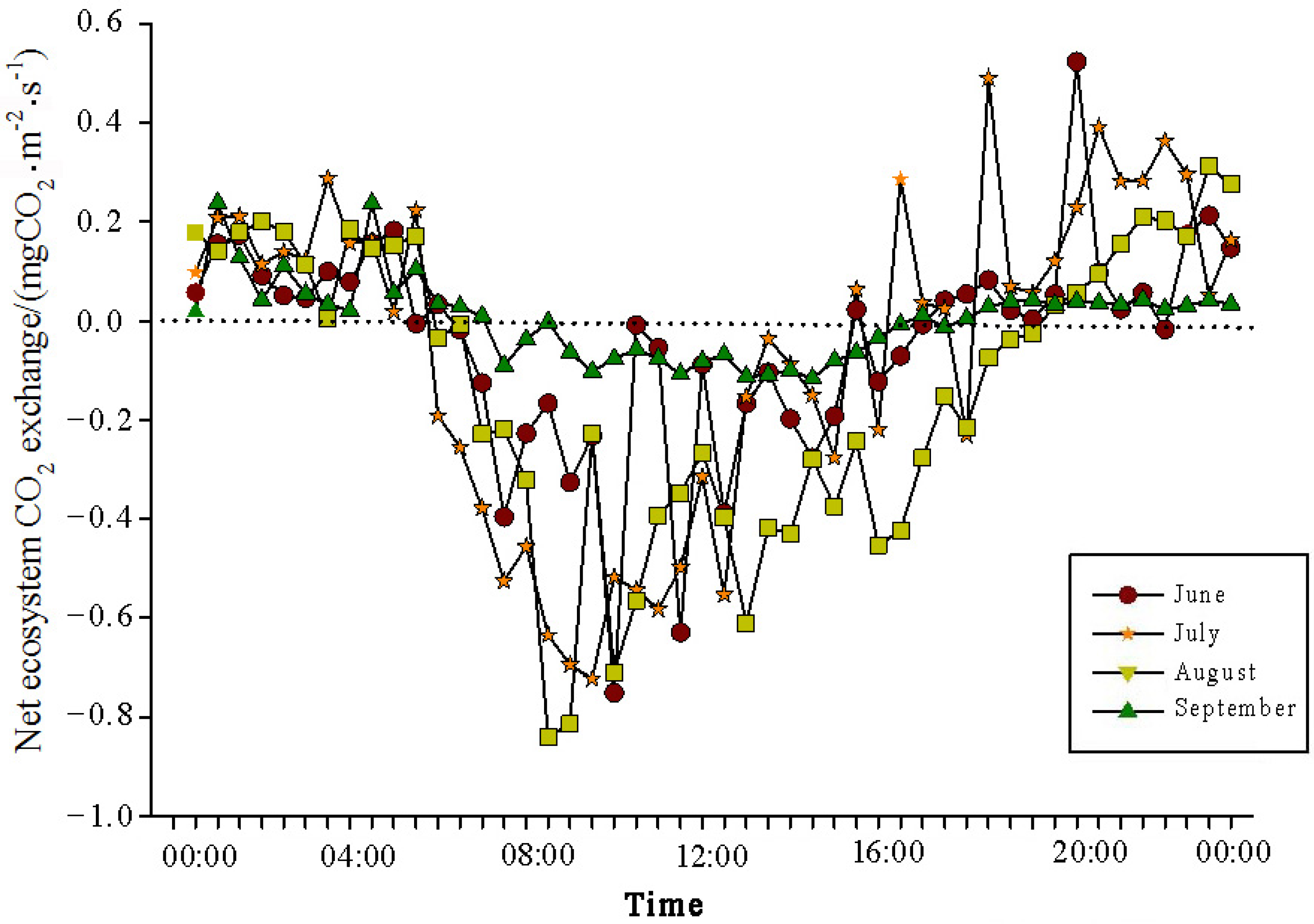
The analysis of the daily variations in CO
2 flux in the Momoge salt marsh ecosystem over four years during the growing season (June–September) showed two trends (
Figure 3,
Figure 4,
Figure 5 and
Figure 6). Throughout the day, the CO
2 flux in the Momoge salt marsh initially declined and subsequently increased. Prior to 6:00 AM, due to the respiration of wetland vegetation, the NEE was positive. After sunrise, around 7:00 AM, reeds initiated photosynthesis, which was stronger than the sum of the autotrophic respiration and heterotrophic respiration of reeds and soils, resulting in carbon uptake. This uptake intensity increased with enhanced solar radiation. From 8:00 AM to 11:00 AM, the carbon uptake rate reached its peak. In the first trend, during the midday period from 11:00 AM to 1:00 PM, carbon uptake diminished due to light saturation, a phenomenon often referred to as midday depression. Around 4:00 PM in the afternoon, the ecosystem’s carbon sequestration capacity started to increase again, reaching the second peak of CO
2 uptake (
Figure 3 and
Figure 5). Subsequently, the carbon uptake gradually decreased. Around 5:00 PM, the net carbon exchange shifted from negative to positive values, with NEE approaching zero. During this time, the vegetation mainly engaged in respiration, releasing CO
2. The ecosystem began to emit CO
2 to the atmosphere, and photosynthesis essentially ceased. In the second trend, the ecosystem only had one uptake peak (
Figure 4) from 12:00 PM to 1:00 PM. After reaching the absorption peak around 12:00 PM, the absorption was suppressed in the afternoon due to light saturation. The amount of absorbed CO
2 began to decrease, and the rate of carbon sequestration in the ecosystem declined gradually. From approximately 7:00 PM to 8:00 PM, the ecosystem became a CO
2 source.
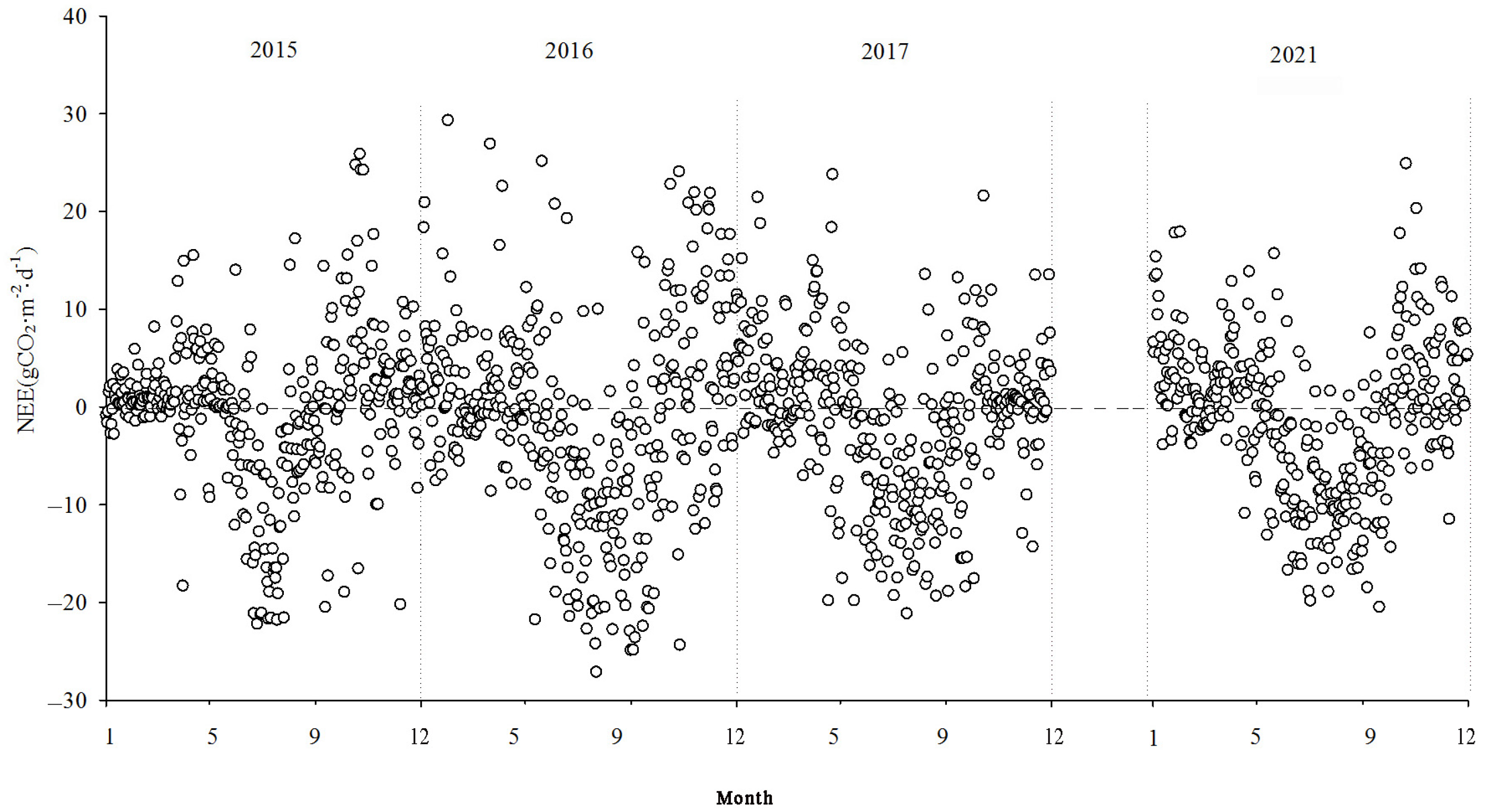
3.2. Seasonal and Interannual Dynamics of Net CO2 Exchange
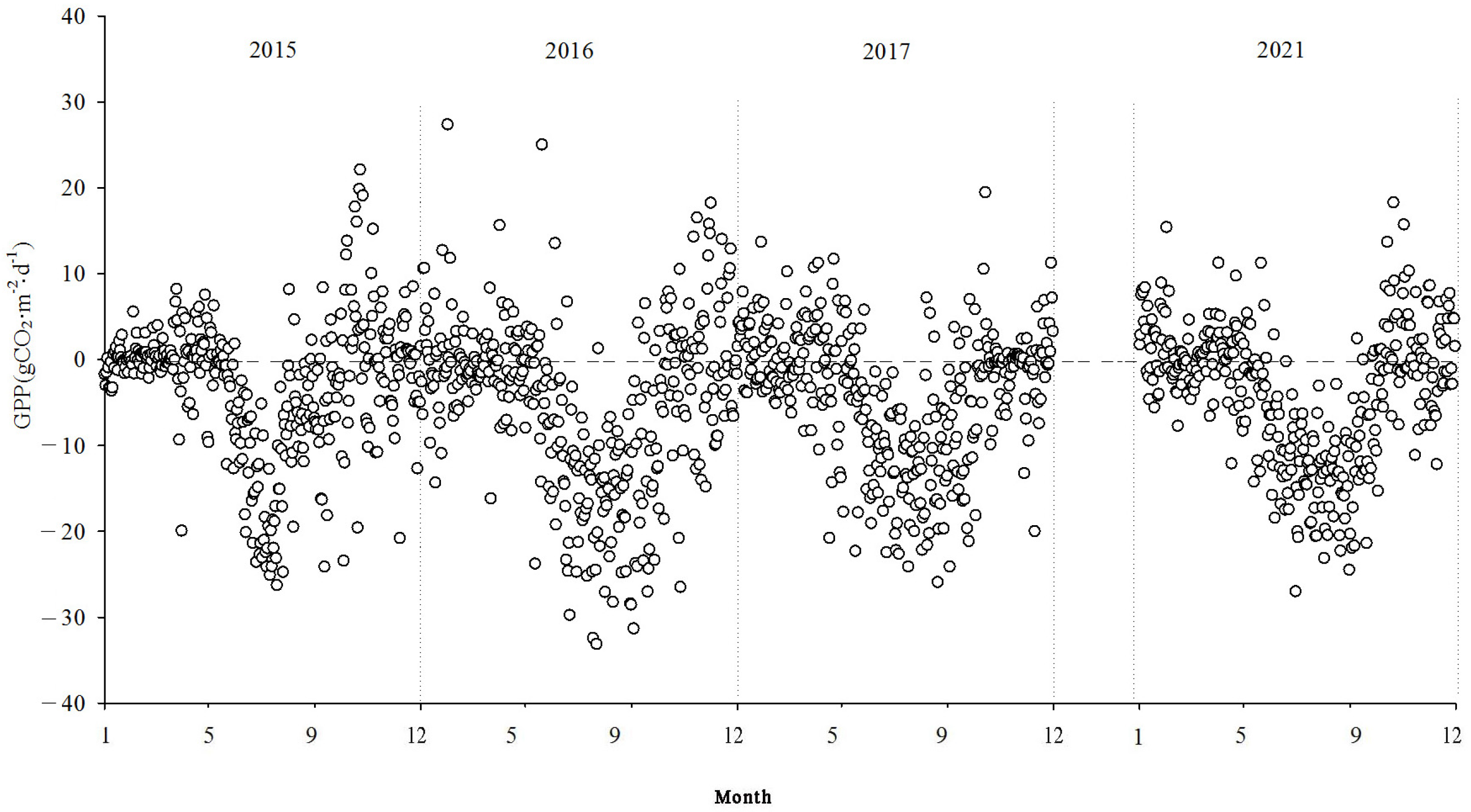
Figure 7,
Figure 8 and
Figure 9 show the interannual dynamics of different components of CO
2 fluxes in the Momoge salt marsh from 2015 to 2017 and in 2021, where the NEE, GPP, and ecosystem respiration (Reco) are daily cumulative values (g CO
2·m
−2·d
−1). The GPP and Reco were estimated using the NEE.
During the study period, the NEE values illustrated the variations in carbon source/sink dynamics within the study area. Each year, the variation in carbon flux in the study area approximately followed a “V” shape. During the non-growing seasons, the carbon flux tended to be positive, indicating CO2 emissions into the atmosphere. The Momoge salt marsh ecosystem acted as a CO2 source. During the growing season, the carbon flux was predominantly negative, indicating the absorption of CO2 from the atmosphere. The Momoge salt marsh functioned as a carbon sink during this period. Throughout the observation period, the average daily cumulative NEE value was approximately −1.03 g CO2·m−2·d−1. The numbers of days with negative carbon flux were 163, 202, 193, and 186 in 2015, 2016, 2017, and 2021, respectively. The proportion of negative carbon flux days was 51% for the entire observation period. During the high vegetation growth period in summer, the carbon flux values could be relatively high, even reaching positive values. This situation was typically observed during rainy or cloudy days between June and September. Due to the low PAR under such conditions, the NEE values generally tended to be small. With weaker vegetation photosynthesis and elevated temperatures, ecosystem respiration intensified, resulting in the wetland ecosystem acting as a carbon source.
At the interannual scale, the monthly mean daily dynamics of NEE exhibited a characteristic “V”-shaped curve, showing significant variations across seasons. During the non-growing season, the overall fluctuations were relatively small, whereas during the growing season, the overall fluctuations were more pronounced.
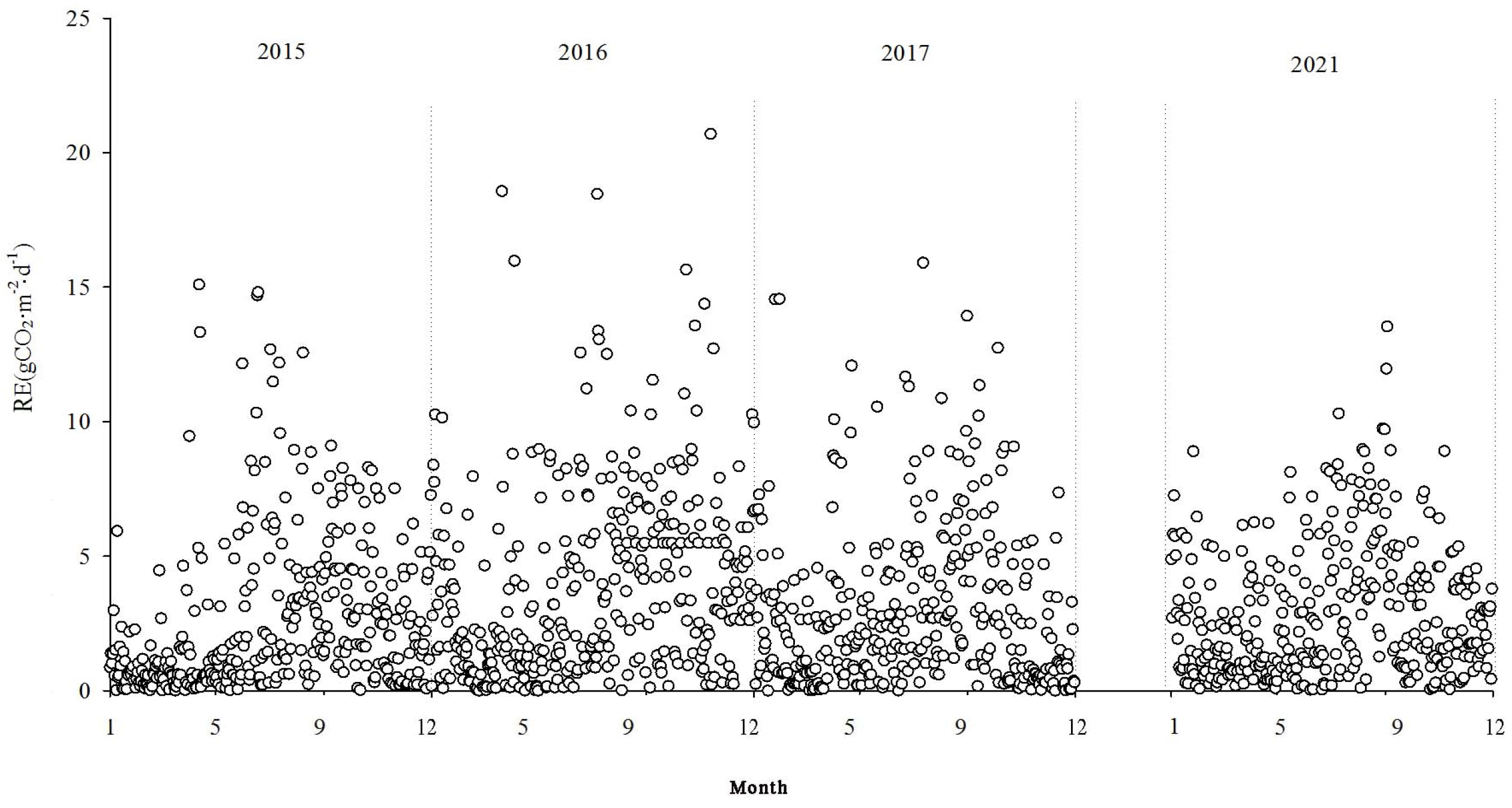
Reco encompasses both soil and plant respiration. During the non-growing season, the Reco values were relatively low but still positive, indicating the emission of CO2 into the atmosphere. As temperatures rose, Reco gradually increased, reaching its peak during the summer months. During the study period, Reco values varied within the range of 0.002 g CO2·m−2·d−1–20.702 g CO2·m−2·d−1, with a mean value of 3.08 g CO2·m−2·d−1.
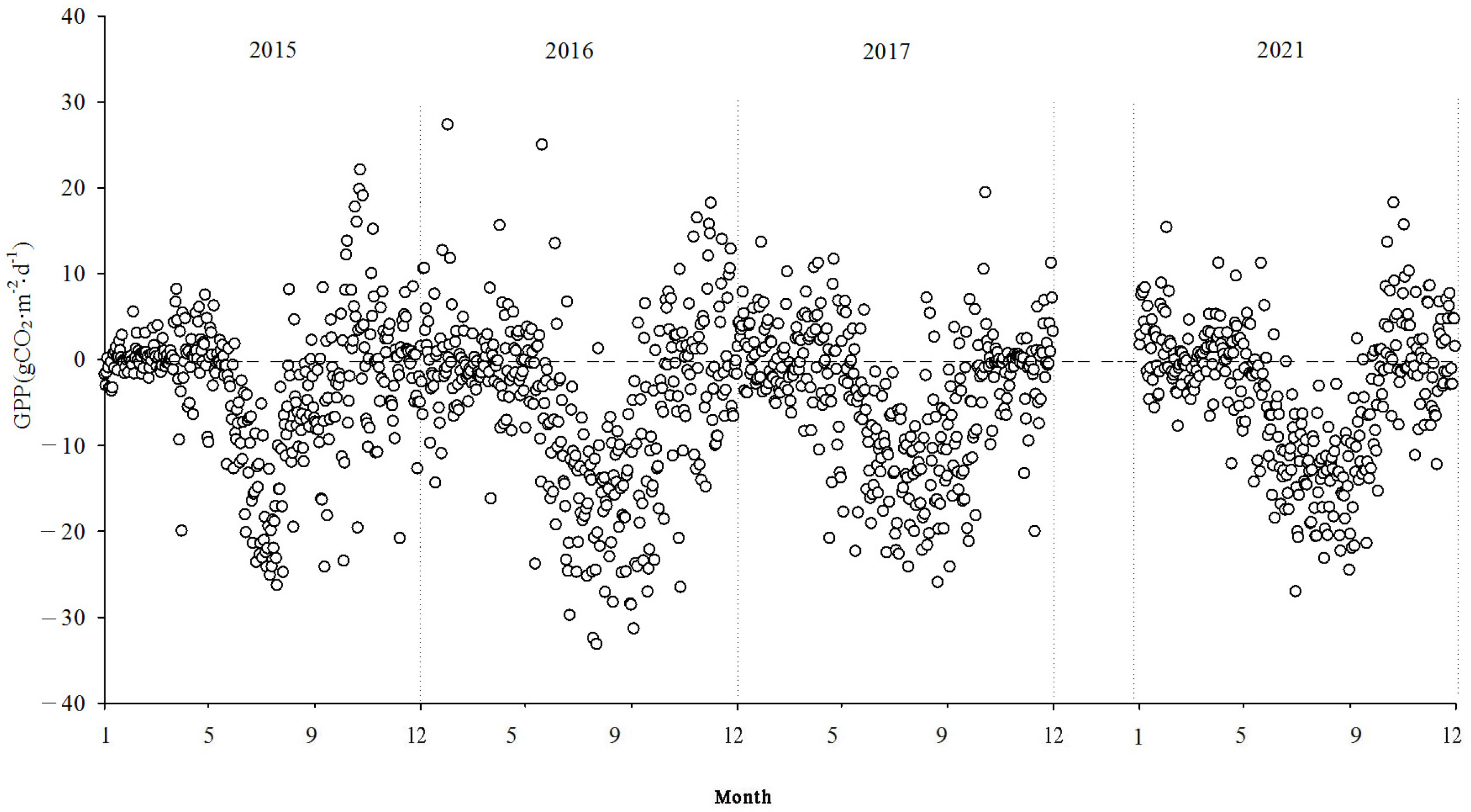
The total GPP of the ecosystem exhibited a yearly “V”-shaped pattern. During the non-growing season, the GPP values were relatively low. In the early growing season, with the emergence of reeds and the increase in leaf area, both the GPP and Reco started to rise. As the vegetation’s physiological activities intensified and photosynthesis dominated respiration, the wetland ecosystem acted as a carbon sink. During the main growing period (June to September), the GPP and Reco grew gradually.
3.3. Ecosystem CO2 Balances
The dynamics of ecosystem CO
2 exchange were closely linked to climate conditions and vegetation, displaying significant seasonal and interannual variations. The ecosystem acted as a carbon sink during the growing season (May–October) and as a carbon source during the non-growing season (December–April) (
Figure 10).
The four distinct seasons of the climatic conditions and vegetation in the Momoge salt marsh led to noticeable variations in the CO
2 flux of the wetland ecosystem. This could be observed in the average daily data for each month. By calculating the daily average CO
2 flux for each month from 2015 to 2017 and in 2021, a monthly average CO
2 flux graph was generated. As shown in
Figure 8, the monthly average CO
2 flux was positive from January to April and from October to December. This indicates that the ecosystem emitted CO
2 to the atmosphere, acting as a carbon source. In May, as reeds started to sprout, photosynthesis intensified and the amount of CO
2 absorbed by reeds gradually increased. From June to September, the wetland entered its growing season, and the monthly average carbon flux values were negative. July marked the peak, where the ecosystem absorbed CO
2 from the atmosphere. During this period, the reed wetlands exhibited significant carbon sink activity during the day. Subsequently, as the reeds entered the senescence phase, the amount of absorbed CO
2 decreased, and the system shifted back to emitting carbon. Overall, within the study period, the wetland ecosystem exhibited its strongest carbon sequestration capacity in July, with a minimum monthly cumulative carbon flux of up to −359.98 g CO
2·m
−2 (July 2021). On the other hand, the month of January had the highest carbon emissions to the atmosphere, with a maximum monthly cumulative carbon flux of 185.89 g CO
2·m
−2 (October 2015). The CO
2 flux differed significantly among the different months in each year, with the CO
2 flux values from June to September notably lower than those in the other months (F = 29.79,
p < 0.001).
The NEE monitoring over the four years in the Momoge salt marsh revealed that it functions as a carbon sink overall. The carbon sequestration intensities for each year were as follows: 206.94 g CO2·m−2·yr−1, 500.28 g CO2·m−2·yr−1, 436.88 g CO2·m−2·yr−1, and 358.61 g CO2·m−2·yr−1 in 2015, 2016, 2017, and 2021, respectively. The cumulative GPP values were −1152.62 g CO2·m−2·yr−1, −1928.62 g CO2·m−2·yr−1, −1516.79 g CO2·m−2·yr−1, and −1409.74 g CO2·m−2·yr−1. The cumulative Reco values were 945.68 g CO2·m−2·yr−1, 1428.34 g CO2·m−2·yr−1, 1079.91 g CO2·m−2·yr−1, and 1051.13 g CO2·m−2·yr−1, respectively.
3.4. Effects of Environmental Factors on CO2 Exchange in the Wetland Ecosystem
3.4.1. Daily Scale
Pearson correlations were used to analyze the correlations between the factors and the dependent variable. A matrix of the correlation coefficients between the independent variables is shown in
Table 1. NEE represents the daily average CO
2 exchange, Tsoil represents the temperature at a soil depth of 5 cm at the daily scale, Tair represents the daily average air temperature, PAR represents the daily average photosynthetically active radiation, PPT represents the daily precipitation, and WL represents the water level.
From the table, the Tsoil, PAR, PPT, and WL all significantly influenced the net CO2 exchange. Specifically, the Tair, Tsoil, and PAR exhibited negative correlations with net CO2 exchange. As the temperature and PAR increased, the net CO2 exchange of the ecosystem decreased, indicating a trend towards an enhanced carbon absorption capacity. The PPT was negatively correlated with the net CO2 exchange. On rainy days, lower solar radiation resulted in reduced CO2 absorption. The WL showed a positive correlation with the net CO2 exchange. As the WL rose, the proportion of plants exposed to the surface decreased, leading to a reduction in the wetland’s carbon absorption capacity.
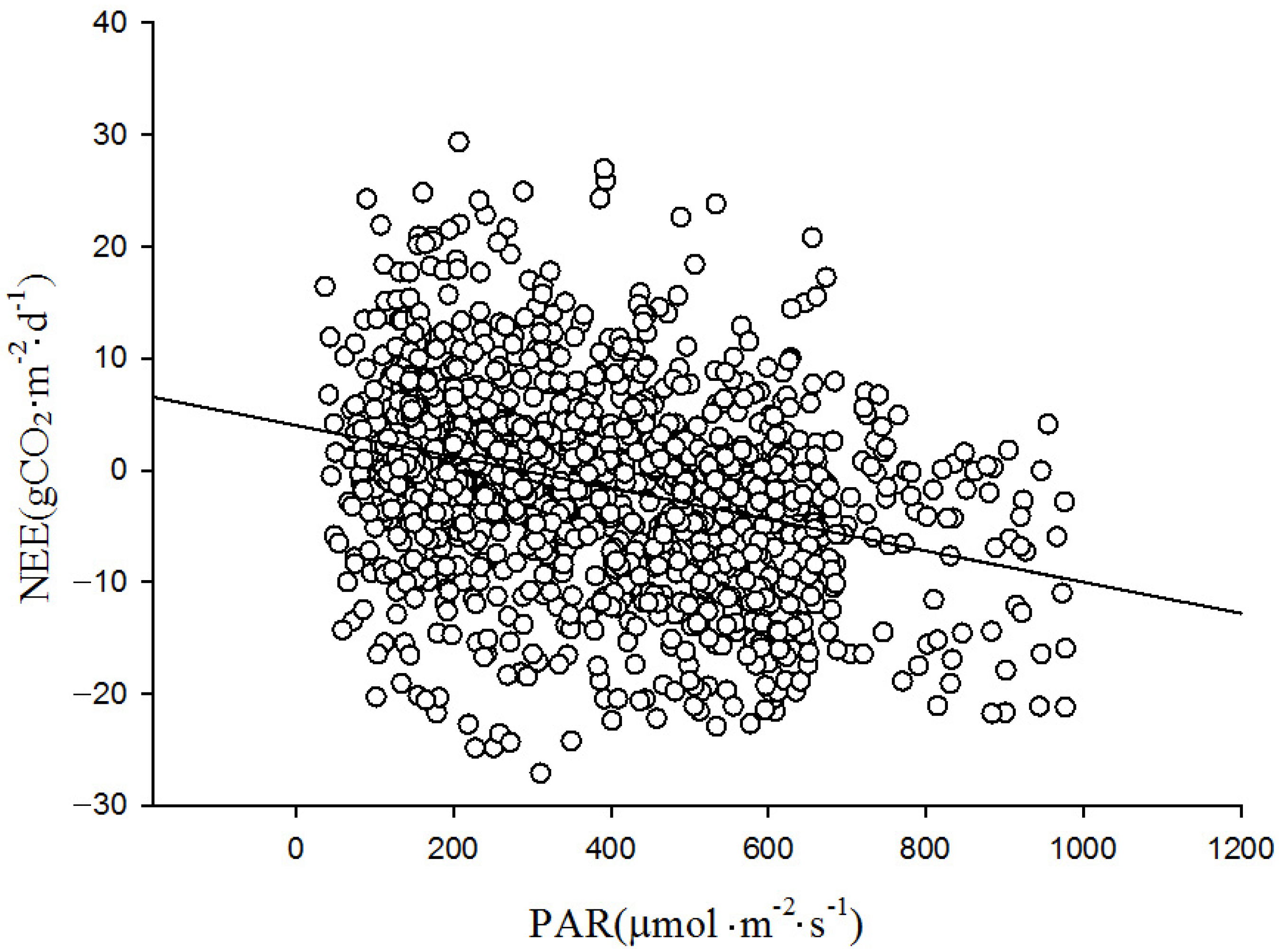
The role of PAR on the net exchange of CO
2 in wetlands was mainly reflected in the daytime of the growing season. In
Figure 11, the net CO
2 exchange evidently increased rapidly with the rise in PAR. However, as the PAR intensity surpassed a certain threshold, the rate of the increase in the net CO
2 exchange slowed, eventually reaching a steady-state value. The influence of PAR was highly significant during different growth stages of the reeds. The phenomenon of net CO
2 absorption was closely related to the growth activities of the reeds. During the sprouting to flowering period from 4 May to 18 August, the wetland exhibited a strong photosynthetic assimilation capacity, with the maximum carbon absorption reaching up to 1.9 mg·m
−2·s
−1. During the early and late stages of reed growth, the carbon absorption capacity was relatively weak, with the maximum carbon absorption reaching only 0.94 mg·m
−2·s
−1.
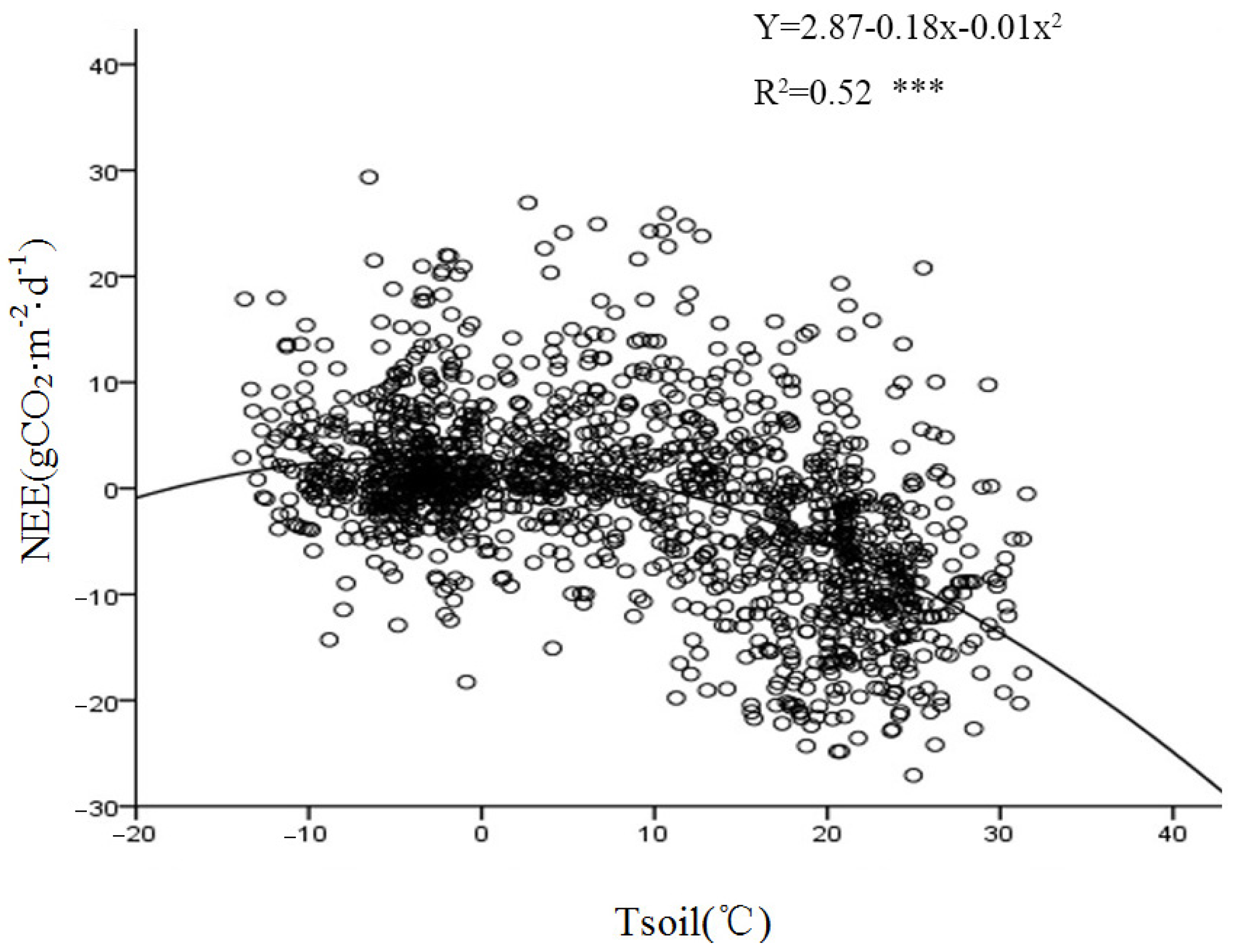
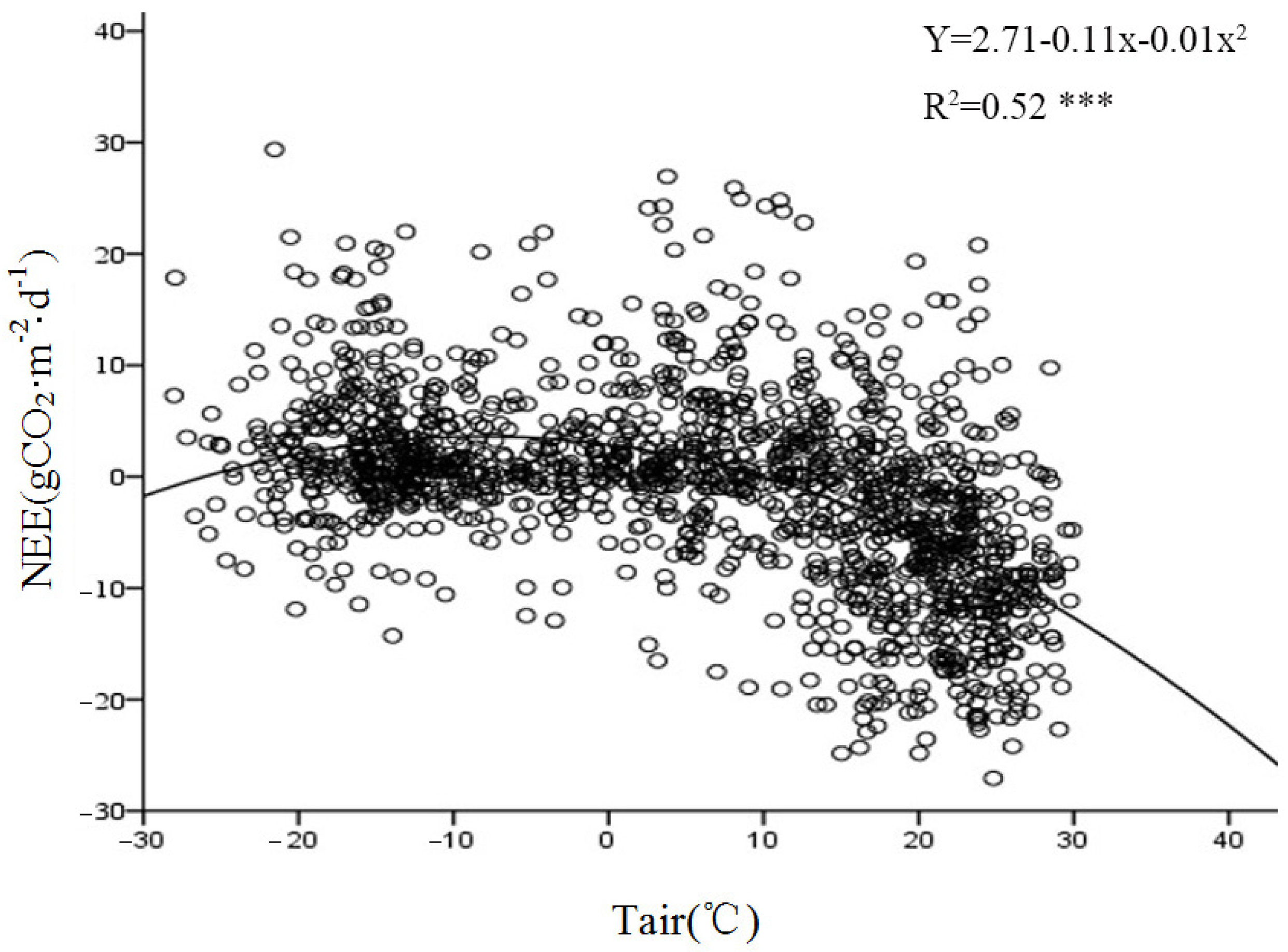
The temperature during the growing season had a significant impact on the photosynthetic physiology of the vegetation and was the most important meteorological factor affecting the respiration intensities of various components within the ecosystem.
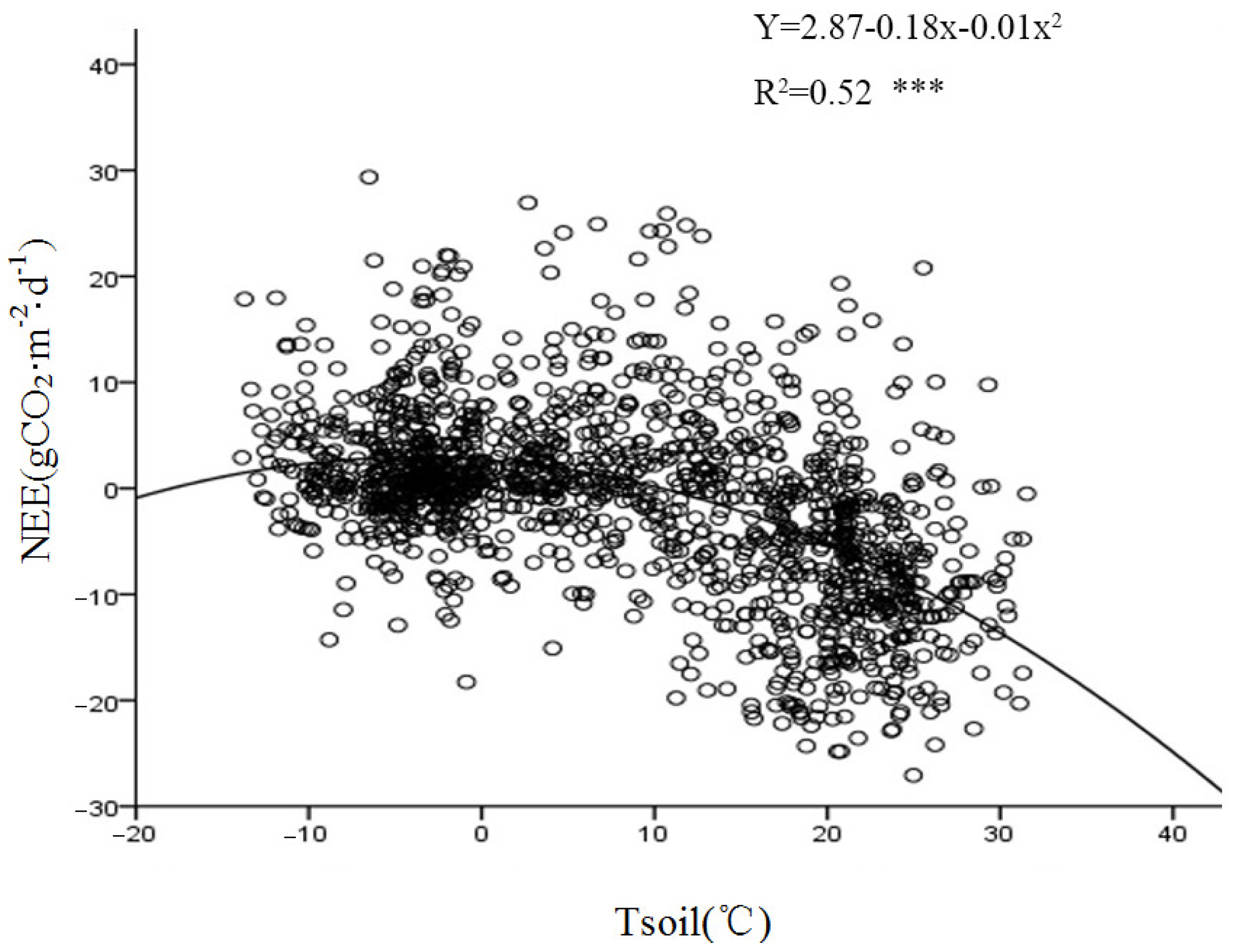
Figure 12 demonstrates a significant exponential correlation between the nighttime CO
2 flux and Tsoil at a depth of 5 cm. A comparative analysis of the relationship between the nighttime carbon flux and the 2.5 m air temperature during the growing and non-growing seasons (
Figure 13) revealed an exponential growth trend in the response of the nighttime carbon flux to Tair. This suggests that the intensity of nighttime carbon release is directly controlled by thermal conditions. Both Tair and the upper Tsoil affected the respiration rate of the ecosystem. An increase in temperature accelerated the metabolism of plants and microorganisms, thus enhancing the intensity of carbon emissions.
3.4.2. Seasonal Scale
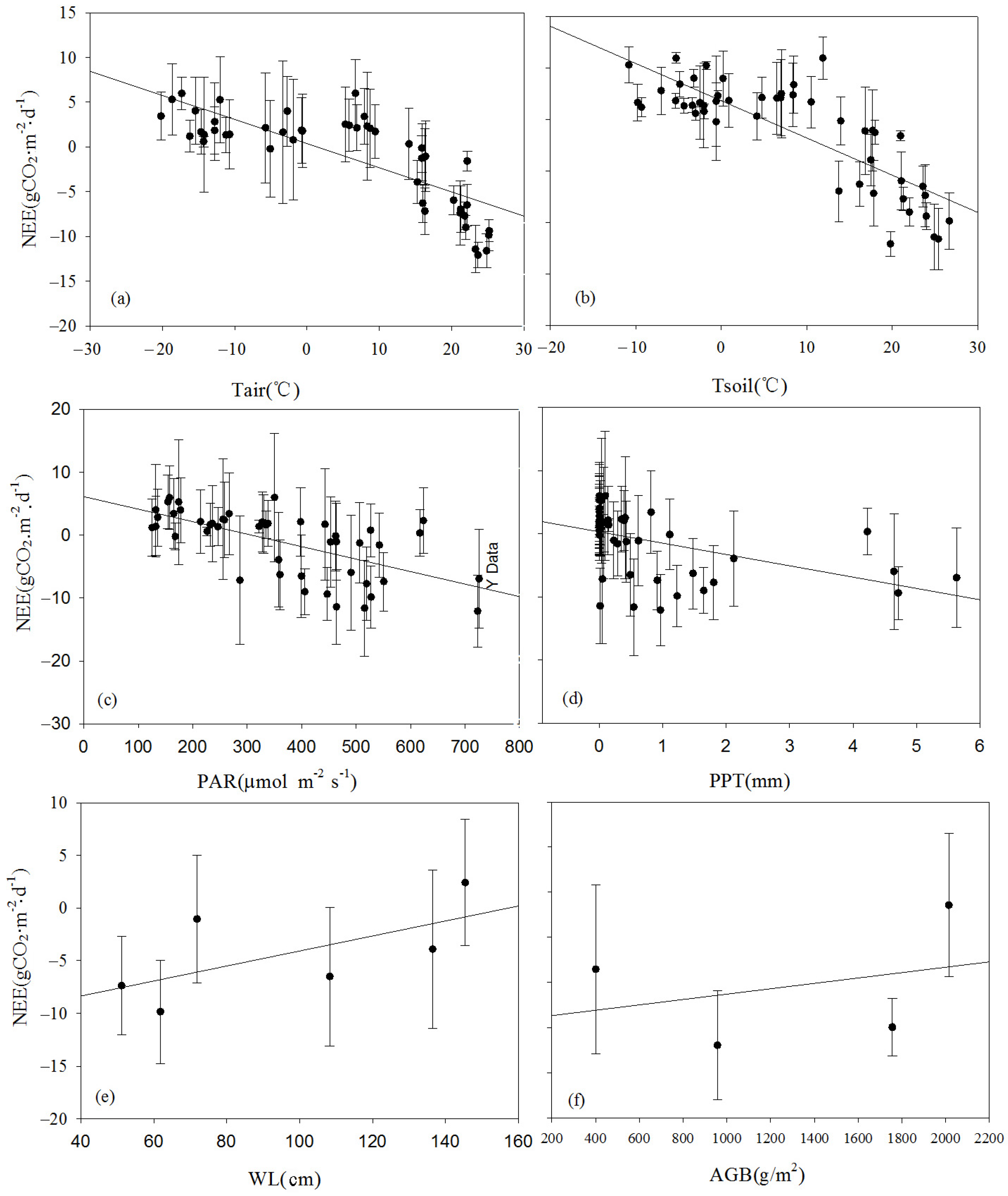
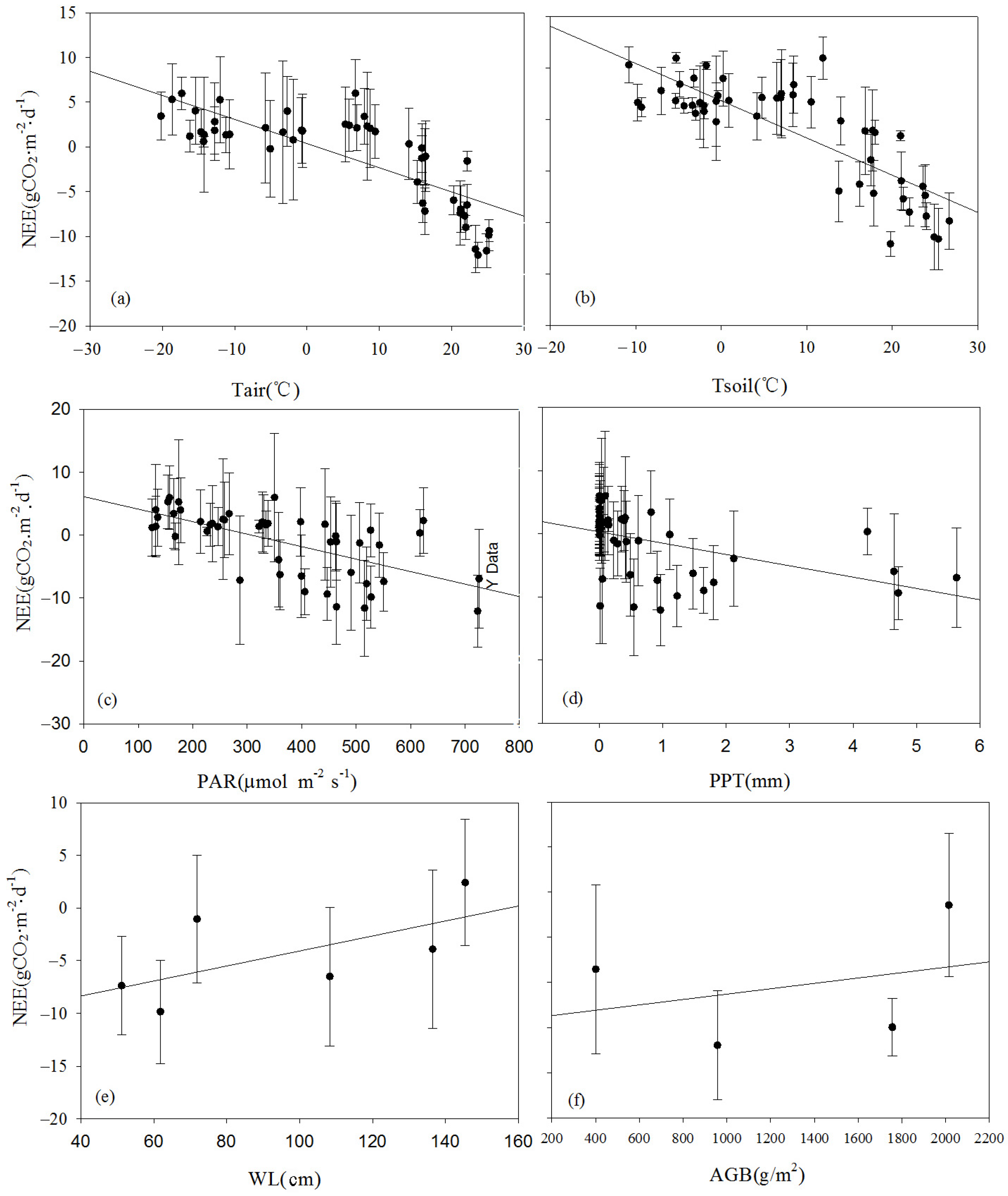
The effects of environmental control variables (Tair, Tsoil, PAR, PPT, WL, and AGB) on NEE are often complex and interrelated. To investigate the mechanisms influencing NEE, a correlation analysis was conducted between the monthly average NEE and the environmental factors at the seasonal scale (
Table 2). Apart from the WL, significant linear correlations between the monthly average NEE and the main environmental and biological factors were found (
Figure 14). They could be ranked in decreasing order of significance as follows: Tsoil > Tair > PAR > PPT > biomass (AGB). Larger values of Tsoil, Tair, PAR, and AGB indicated a stronger carbon uptake capacity of the ecosystem. Tair significantly influenced Tsoil, indicating that Tair indirectly affected NEE flux by influencing Tsoil. Concurrently, the monthly carbon uptake capacity significantly increased with an increase in Tair. The dynamics of AGB were more dependent on the variations in PAR. This was primarily because the monitoring area experienced prolonged waterlogged conditions, where the anaerobic environment inhibited the decomposition of detritus, leading to the accumulation of organic matter in the soil. As a result, the wetland became a carbon sink that suppressed the increase in the atmospheric CO
2 concentration. The impact of rainfall on biomass accumulation was relatively weak. Under waterlogged conditions, the reed biomass was relatively high, with the maximum biomass reaching up to 2.5 kg·m
−2. The plant height could exceed 270 cm, and the ground cover density was relatively high.
4. Discussion
Different wetland ecosystems exhibit varying net ecosystem CO
2 exchange rates across different time scales. Similarly, CO
2 exchange rates differ among wetland types. Even within the same wetland type, interannual CO
2 exchange varies notably. Currently, wetlands generally act as carbon sinks, with an average carbon sequestration rate of 118 gC·m
−2·yr
−1 [
7]. Numerous studies have indicated that reed wetlands and peatlands are significant carbon sinks [
15]. Our study confirmed this by indicating that the Momoge salt marsh acted as a carbon sink in the years 2015–2017 and 2021. A study monitoring the NEE in the reed wetland of the Liaohe Delta in 2006 showed that this area also acted as a carbon sink, with a carbon sequestration intensity of 230 g CO
2·m
−2·yr
−1 [
16]. In the Nanji wetland of Poyang Lake, during the non-flooded period of 2015, the ecosystem acted as a carbon sink with a carbon sequestration rate of 667.62 g C·m
−2·yr
−1. However, in the subsequent years of 2016 and 2017, it shifted to a carbon source [
17]. A study of CO
2 fluxes for the entire year of 2005 in the Haibei wetland of the Qinghai–Tibet Plateau showed that it released around 86.18 g C·m
−2·yr
−1 into the atmosphere, making it a carbon source as well [
18]. The interannual variations in NEE in wetland ecosystems are closely linked to climate [
19,
20,
21], hydrology [
22], and vegetation conditions [
23]. These factors collectively play a significant role in shaping the carbon sink functions of specific ecosystems. Furthermore, related studies have indicated that even within the same wetland ecosystem, the carbon source/sink status can exhibit uncertainty across interannual periods due to variations in vegetation composition, hydrological conditions, human disturbances, and climate change [
24]. Since wetland greenhouse gas balances are highly sensitive to changes in wetland area, the future impact of wetlands on the climate depends on the balance between degradation and restoration.
This study found that the NEE of wetlands exhibited a pattern of functioning as a carbon sink with greater fluctuations during the growing season and as a carbon source with smaller fluctuations during the non-growing season. During the spring, as plants start sprouting and growing, the amount of carbon fixed through photosynthesis is initially less than the carbon released through respiration. As a result, the wetland continues to act as a carbon source. However, as plant growth progresses and photosynthesis becomes stronger than respiration, the wetland transitions to a carbon sink. Along with the peak of plant growth, the maximum values of NEE typically occur in July and August. Subsequently, as temperatures decrease and plants enter their senescence phase, their photosynthetic capacity gradually diminishes. This reduction in photosynthesis leads to a significant decrease in the NEE. This pattern is consistent with findings from previous research [
25]. By November, the wetland vegetation is completely withered. Due to ecosystem respiration, the wetland transitions entirely to functioning as a carbon source.
A significant amount of research has indicated that when the solar radiation intensity is low, net CO
2 exchange increases with the increase in PAR, following a hyperbolic trend. Many studies have shown a significant correlation between NEE and PAR. Ecosystems such as forests, grasslands, and croplands exhibit a positive relationship between solar radiation intensity and net carbon exchange, following a hyperbolic curve [
26]. Similar conclusions were drawn by Beverland et al. [
27] in the context of wetland ecosystems. The current study also revealed that in the reed wetland, the net CO
2 exchange initially increases rapidly with an increase in PAR. However, after PAR exceeds a certain threshold, the rate of the increase in the net CO
2 exchange slows, eventually converging to an asymptote value. The phenomenon of net CO
2 absorption is closely tied to the growth activities of reeds. Furthermore, related studies have shown that the response of NEE to solar radiation is also influenced by other limiting factors, such as the atmospheric vapor pressure deficit and occasionally frost [
28,
29]. The influence of the radiation intensity on the net CO
2 exchange is more pronounced within a suitable temperature range. Additionally, a larger leaf area index corresponds to a more distinct response of net CO
2 exchange to radiation intensity.
Most studies suggest that changes in WL and temperature increases can significantly affect the production of CO
2 in wetland ecosystems [
30,
31,
32]. Research on wetlands in southern Finland found that a high temperature is the primary factor leading to an increase in net carbon uptake [
33]. This study found that nighttime carbon flux exhibits an exponential growth trend in response to Tair. Temperature affects ecosystem CO
2 flux through both direct and indirect mechanisms, ultimately influencing ecological processes [
34]. Firstly, the seasonal variation in photosynthesis is primarily controlled by temperature [
35]. Within a certain temperature range, higher temperatures are more favorable for photosynthesis [
36]. For example, studies on the seasonal cumulative GPP of corn and wheat ecosystems found that higher mean temperatures are associated with larger cumulative GPP values [
37]. Research on temperate bulrush marshes also showed that temperature is a key indicator for daily GPP [
38]. Furthermore, ecosystem respiration is primarily controlled by temperature, with respiration increasing exponentially as temperature rises [
39]. This is because higher temperatures can stimulate dark respiration in both soil and vegetation. Research on northern marshes revealed that higher temperatures correspond to higher ecosystem respiration rates [
40]. At both interannual and seasonal scales, the Reco of salt marshes was found to be significantly correlated with temperature [
41]. Furthermore, CO
2 fluxes are controlled by temperature through its effects on vegetation cover, the leaf area index, and the length of the growing season. An increase in Tsoil is accompanied by an increase in AGB [
42]. The temperature in the environment controls the humidity by regulating the vapor pressure. Research on wetlands in the southwestern plateau of China found that temperature can explain most of the seasonal variability in carbon fluxes. Warming can increase both GPP and Reco, thus influencing the carbon balance of the plateau wetlands [
43]. Although water level conditions significantly influence the uptake and release of CO
2 in the ecosystem, temperature remains the primary factor for carbon fluxes in the Momoge salt marsh.
5. Conclusions
This study analyzed the CO2 flux variation characteristics and the factors affecting them in the Momoge salt marsh ecosystem. During the growing season, we observed an initial decline and a subsequent increase in the daily variations in CO2 fluxes in the Momoge salt marsh, with uptake occurring during the day (negative values) and release taking place at night (positive values). Two patterns emerged (one peak and two peaks), with reduced carbon absorption during midday due to light saturation resulting in a midday depression phenomenon for the latter pattern. Within each annual variation, the carbon fluxes in the study area roughly exhibited a “V” shape. The carbon fluxes during the non-growing season predominantly showed positive values, indicating the release of CO2 into the atmosphere. The carbon sequestration intensities of the Momoge salt marsh ecosystem were 206.94 g CO2·m−2·yr−1, 500.28 g CO2·m−2·yr−1, 436.00 g CO2·m−2·yr−1, and 358.61 g CO2·m−2·yr−1 for the years 2015–2017 and 2021, respectively. This indicates that the system behaves as a carbon sink.
At both the hourly and daily scales, PAR was the primary factor affecting daytime variations in NEE during the growing season, while temperature was the main factor influencing nighttime NEE dynamics. At the daily scale, Tair, Tsoil, PAR, PPT, and WL all showed significant effects on net CO2 exchange. At the monthly scale, larger values of Tsoil, Tair, PAR, and AGB corresponded to a stronger carbon absorption capacity of the ecosystem. As the WL rose, the proportion of plants exposed to the surface decreased, leading to a reduction in the wetland’s carbon absorption capacity. Although the WL conditions significantly influenced the uptake and release of CO2 in the ecosystem, temperature remained the primary factor for carbon fluxes in the Momoge salt marsh.







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}