
Influence of Sports on Cortical Connectivity in Patients with Spinal Cord Injury-A High-Density EEG Study

 , , ,
, , , 
Abstract
1. Introduction
2. Methods
2.1. Ethics
2.2. Participants
2.3. Paradigm
2.4. HD-EEG Data Recording
2.5. HD-EEG Data Preprocessing
2.6. HD-EEG Connectivity Analysis
- Watching the movie + beep was subtracted from movement imagination + beep.
- Watching the movie + beep was subtracted from movement + beep.
- Watching the movie + music was subtracted from movement imagination + music.
- Watching the movie + music was subtracted from movement + music.
2.7. Statistics
3. Results
3.1. Demographic and Clinical Data
3.2. Connectivity Analysis
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brasil-Neto, J.P.; Valls-Solè, J.; Pascual-Leone, A.; Cammarota, A.; Amassian, V.E.; Cracco, R.; Maccabee, P.; Cracco, J.; Hallett, M.; Cohen, L.G. Rapid modulation of human cortical motor outputs following ischaemic nerve block. Brain 1993, 116, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Nardone, R.; Höller, Y.; Brigo, F.; Seidl, M.; Christova, M.; Bergmann, J.; Golaszewski, S.; Trinka, E. Functional brain reorganization after spinal cord injury: Systematic review of animal and human studies. Brain Res. 2013, 1504, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, J.; Humanes-Valera, D.; Alonso-Calviño, E.; Yague, J.G.; Moxon, K.A.; Oliviero, A.; Foffani, G. Spinal Cord Injury Immediately Changes the State of the Brain. J. Neurosci. 2010, 30, 7528–7537. [Google Scholar] [CrossRef] [PubMed]
- Wall, P.D.; Egger, M.D. Formation of New Connexions in Adult Rat Brains after Partial Deafferentation. Nature 1971, 232, 542–545. [Google Scholar] [CrossRef] [PubMed]
- Merzenich, M.M.; Nelson, R.J.; Stryker, M.P.; Cynader, M.S.; Schoppmann, A.; Zook, J.M. Somatosensory cortical map changes following digit amputation in adult monkeys. J. Comp. Neurol. 1984, 224, 591–605. [Google Scholar] [CrossRef] [PubMed]
- Endo, T.; Spenger, C.; Tominaga, T.; Brené, S.; Olson, L. Cortical sensory map rearrangement after spinal cord injury: fMRI responses linked to Nogo signalling. Brain 2007, 130 Pt 11, 2951–2961. [Google Scholar] [CrossRef] [PubMed]
- Humanes-Valera, D.; Foffani, G.; Alonso-Calviño, E.; Fernández-López, E.; Aguilar, J. Dual Cortical Plasticity after Spinal Cord Injury. Cereb. Cortex 2017, 27, 2926–2940. [Google Scholar] [CrossRef]
- Wrigley, P.J.; Press, S.R.; Gustin, S.M.; Macefield, V.G.; Gandevia, S.C.; Cousins, M.J.; Middleton, J.W.; Henderson, L.A.; Siddall, P.J. Neuropathic pain and primary somatosensory cortex reorganization following spinal cord injury. Pain 2009, 141, 52–59. [Google Scholar] [CrossRef]
- Henderson, L.A.; Gustin, S.M.; Macey, P.M.; Wrigley, P.J.; Siddall, P.J. Functional Reorganization of the Brain in Humans Following Spinal Cord Injury: Evidence for Underlying Changes in Cortical Anatomy. J. Neurosci. 2011, 31, 2630–2637. [Google Scholar] [CrossRef]
- Nardone, R.; Langthaler, P.B.; Bathke, A.C.; Höller, Y.; Brigo, F.; Lochner, P.; Christova, M.; Trinka, E. Effects of passive pedaling exercise on the intracortical inhibition in subjects with spinal cord injury. Brain Res. Bull. 2016, 124, 144–149. [Google Scholar] [CrossRef]
- Pearce, A.J.; Thickbroom, G.W.; Byrnes, M.L.; Mastaglia, F.L. Functional reorganisation of the corticomotor projection to the hand in skilled racquet players. Exp. Brain Res. 2000, 130, 238–243. [Google Scholar] [CrossRef]
- Fourkas, A.D.; Bonavolontà, V.; Avenanti, A.; Aglioti, S.M. Kinesthetic Imagery and Tool-Specific Modulation of Corticospinal Representations in Expert Tennis Players. Cereb. Cortex 2008, 18, 2382–2390. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.; Guillot, A.; MacIntyre, T.; Collet, C. Re-imagining mental imagery: Building bridges between cognitive neuroscience and sport psychology. Br. J. Psychol. 2012, 103, 224–247. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, F.; Guillot, A.; Daligault, S.; Delpuech, C.; Rode, G.; Collet, C. Motor inhibition during motor imagery: A MEG study with a quadriplegic patient. Neurocase 2014, 20, 524–539. [Google Scholar] [CrossRef] [PubMed]
- Lacourse, M.G.; Cohen, M.J.; Lawrence, K.E.; Romero, D.H. Cortical potentials during imagined movements in individuals with chronic spinal cord injuries. Behav. Brain Res. 1999, 104, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Cramer, S.C.; Orr, E.L.R.; Cohen, M.J.; Lacourse, M.G. Effects of motor imagery training after chronic, complete spinal cord injury. Exp. Brain Res. 2007, 177, 233–242. [Google Scholar] [CrossRef] [PubMed]
- Opsommer, E.; Chevalley, O.; Korogod, N. Motor imagery for pain and motor function after spinal cord injury: A systematic review. Spinal Cord 2020, 58, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Athanasiou, A.; Terzopoulos, N.; Pandria, N.; Xygonakis, I.; Foroglou, N.; Polyzoidis, K.; Bamidis, P.D. Functional Brain Connectivity during Multiple Motor Imagery Tasks in Spinal Cord Injury. Neural Plast. 2018, 2018, 9354207. [Google Scholar] [CrossRef]
- Mateo, S.; Di Rienzo, F.; Bergeron, V.; Guillot, A.; Collet, C.; Rode, G. Motor imagery reinforces brain compensation of reach-to-grasp movement after cervical spinal cord injury. Front. Behav. Neurosci. 2015, 9, 234. [Google Scholar] [CrossRef]
- Müller-Putz, G.R.; Scherer, R.; Pfurtscheller, G.; Rupp, R. EEG-based neuroprosthesis control: A step towards clinical practice. Neurosci. Lett. 2005, 382, 169–174. [Google Scholar] [CrossRef]
- Müller-Putz, G.R.; Zimmermann, D.; Graimann, B.; Nestinger, K.; Korisek, G.; Pfurtscheller, G. Event-related beta EEG-changes during passive and attempted foot movements in paraplegic patients. Brain Res. 2007, 1137, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Pfurtscheller, G.; Linortner, P.; Winkler, R.; Korisek, G.; Müller-Putz, G. Discrimination of Motor Imagery-Induced EEG Patterns in Patients with Complete Spinal Cord Injury. Comput. Intell. Neurosci. 2009, 2009, 104180. [Google Scholar] [CrossRef] [PubMed]
- Abiri, R.; Borhani, S.; Sellers, E.W.; Jiang, Y.; Zhao, X. A comprehensive review of EEG-based brain–computer interface paradigms. J. Neural Eng. 2019, 16, 011001. [Google Scholar] [CrossRef] [PubMed]
- Kazim, S.F.; Bowers, C.A.; Cole, C.D.; Varela, S.; Karimov, Z.; Martinez, E.; Ogulnick, J.V.; Schmidt, M.H. Corticospinal Motor Circuit Plasticity after Spinal Cord Injury: Harnessing Neuroplasticity to Improve Functional Outcomes. Mol. Neurobiol. 2021, 58, 5494–5516. [Google Scholar] [CrossRef] [PubMed]
- MATLAB, Version 9.5.0 (R2018b); The MathWorks Inc.: Natick, MA, USA, 2018.
- Brainard, D.H. The Psychophysics Toolbox. Spat. Vis. 1997, 10, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Pelli, D.G.; Vision, S. The VideoToolbox software for visual psychophysics: Transforming numbers into movies. Spat. Vis. 1997, 10, 437–442. [Google Scholar] [CrossRef] [PubMed]
- Keil, A.; Debener, S.; Gratton, G.; Junghöfer, M.; Kappenman, E.S.; Luck, S.J.; Luu, P.; Miller, G.A.; Yee, C.M. Committee report: Publication guidelines and recommendations for studies using electroencephalography and magnetoencephalography. Psychophysiology 2014, 51, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Ferree, T.C.; Luu, P.; Russell, G.S.; Tucker, D.M. Scalp electrode impedance, infection risk, and EEG data quality. Clin. Neurophysiol. 2001, 112, 536–544. [Google Scholar] [CrossRef]
- Ngo, D.; Sun, Y.; Genton, M.G.; Wu, J.; Srinivasan, R.; Cramer, S.C.; Ombao, H. An exploratory data analysis of electroencephalograms using the functional boxplots approach. Front. Neurosci. 2015, 9, 282. [Google Scholar] [CrossRef]
- Schlögl, A.; Brunner, C. BioSig: A Free and Open Source Software Library for BCI Research. Computer 2008, 41, 44–50. [Google Scholar] [CrossRef]
- Kus, R.; Kaminski, M.; Blinowska, K. Determination of EEG Activity Propagation: Pair-Wise versus Multichannel Estimate. IEEE Trans. Biomed. Eng. 2004, 51, 1501–1510. [Google Scholar] [CrossRef] [PubMed]
- Schlögl, A.; Supp, G. Analyzing event-related EEG data with multivariate autoregressive parameters. Prog. Brain Res. 2006, 159, 135–147. [Google Scholar] [CrossRef] [PubMed]
- Schlögl, A. Time Series Analysis—A Toolbox for the Use with Matlab; Version 4.6.3; The MathWorks Inc.: Natick, MA, USA, 2021. [Google Scholar]
- Korzeniewska, A.; Mańczak, M.; Kamiński, M.; Blinowska, K.J.; Kasicki, S. Determination of information flow direction among brain structures by a modified directed transfer function (dDTF) method. J. Neurosci. Methods 2003, 125, 195–207. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 1 January 2020).
- Bathke, A.C.; Friedrich, S.; Pauly, M.; Konietschke, F.; Staffen, W.; Strobl, N.; Höller, Y. Testing Mean Differences among Groups: Multivariate and Repeated Measures Analysis with Minimal Assumptions. Multivar. Behav. Res. 2018, 53, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, S.; Konietschke, F.; Pauly, M. MANOVA.RM: A Package for Calculating Test Statistics and Their Resampling Versions for Heteroscedastic Semi-Parametric Multivariate Data or Repeated Measures Designs. R-Package Version 0.1.1. 2017. Available online: https://CRAN.R-project.org/package=MANOVA.RM (accessed on 1 January 2020).
- Konietschke, F.; Friedrich, S.; Brunner, E.; Pauly, M. _rankFD: Rank-Based Tests for General Factorial Designs_. R Package Version 0.1.1. 2022. Available online: https://CRAN.R-project.org/package=rankFD (accessed on 1 January 2020).
- Benjamini, Y.; Krieger, A.M.; Yekutieli, D. Adaptive linear step-up procedures that control the false discovery rate. Biometrika 2006, 93, 491–507. [Google Scholar] [CrossRef]
- de Mello, M.T.; Lauro, F.A.A.; Silva, A.C.; Tufik, S. Incidence of periodic leg movements and of the restless legs syndrome during sleep following acute physical activity in spinal cord injury subjects. Spinal Cord 1996, 34, 294–296. [Google Scholar] [CrossRef] [PubMed]
- Mello, M.T.; Silva, A.C.; Rueda, A.D.; Poyares, D.; Tufik, S. Correlation between K complex, periodic leg movements (PLM), and myoclonus during sleep in paraplegic adults before and after an acute physical activity. Spinal Cord 1997, 35, 248–252. [Google Scholar] [CrossRef] [PubMed][Green Version]
- De Mello, M.; Silva, A.; Esteves, A.; Tufik, S. Reduction of periodic leg movement in individuals with paraplegia following aerobic physical exercise. Spinal Cord 2002, 40, 646–649. [Google Scholar] [CrossRef][Green Version]
- Cremoux, S.; Tallet, J.; Maso, F.D.; Berton, E.; Amarantini, D. Impaired corticomuscular coherence during isometric elbow flexion contractions in humans with cervical spinal cord injury. Eur. J. Neurosci. 2017, 46, 1991–2000. [Google Scholar] [CrossRef]
- Sato, G.; Osumi, M.; Morioka, S. Effects of wheelchair propulsion on neuropathic pain and resting electroencephalography after spinal cord injury. J. Rehabil. Med. 2017, 49, 136–143. [Google Scholar] [CrossRef]
- Saturno, E.; Bonato, C.; Miniussi, C.; Lazzaro, V.; Callea, L. Motor cortex changes in spinal cord injury: A TMS study. Neurol. Res. 2008, 30, 1084–1085. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Hino, T.; Komori, T.; Hirai, S. Loss of the muscle silent period evoked by transcranial magnetic stimulation of the motor cortex in patients with cervical cord lesions. Neurosci. Lett. 2000, 286, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Roy, F.D.; Zewdie, E.T.; Gorassini, M.A. Short-interval intracortical inhibition with incomplete spinal cord injury. Clin. Neurophysiol. 2011, 122, 1387–1395. [Google Scholar] [CrossRef] [PubMed]
- Vega, S.R.; Abel, T.; Lindschulten, R.; Hollmann, W.; Bloch, W.; Strüder, H. Impact of exercise on neuroplasticity-related proteins in spinal cord injured humans. Neuroscience 2008, 153, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Kraeutner, S.; Gionfriddo, A.; Bardouille, T.; Boe, S. Motor imagery-based brain activity parallels that of motor execution: Evidence from magnetic source imaging of cortical oscillations. Brain Res. 2014, 1588, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, H.; Hui, M.; Long, Z.; Jin, Z.; Liu, Y.; Yao, L. Motor execution and motor imagery: A comparison of functional connectivity patterns based on graph theory. Neuroscience 2014, 261, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Kasess, C.H.; Windischberger, C.; Cunnington, R.; Lanzenberger, R.; Pezawas, L.; Moser, E. The suppressive influence of SMA on M1 in motor imagery revealed by fMRI and dynamic causal modeling. Neuroimage 2008, 40, 828–837. [Google Scholar] [CrossRef] [PubMed]
- Solodkin, A.; Hlustik, P.; Chen, E.E.; Small, S.L. Fine Modulation in Network Activation during Motor Execution and Motor Imagery. Cereb. Cortex 2004, 14, 1246–1255. [Google Scholar] [CrossRef]
- Gotts, S.J.; Jo, H.J.; Wallace, G.L.; Saad, Z.S.; Cox, R.W.; Martin, A. Two distinct forms of functional lateralization in the human brain. Proc. Natl. Acad. Sci. USA 2013, 110, E3435–E3444. [Google Scholar] [CrossRef]
- Güntürkün, O.; Ströckens, F.; Ocklenburg, S. Brain Lateralization: A Comparative Perspective. Physiol. Rev. 2020, 100, 1019–1063. [Google Scholar] [CrossRef]
- Iturria-Medina, Y.; Fernández, A.P.; Morris, D.M.; Canales-Rodríguez, E.J.; Haroon, H.A.; Pentón, L.G.; Augath, M.; García, L.G.; Logothetis, N.; Parker, G.J.M.; et al. Brain Hemispheric Structural Efficiency and Interconnectivity Rightward Asymmetry in Human and Nonhuman Primates. Cereb. Cortex 2011, 21, 56–67. [Google Scholar] [CrossRef] [PubMed]
- Hétu, S.; Grégoire, M.; Saimpont, A.; Coll, M.-P.; Eugène, F.; Michon, P.-E.; Jackson, P.L. The neural network of motor imagery: An ALE meta-analysis. Neurosci. Biobehav. Rev. 2013, 37, 930–949. [Google Scholar] [CrossRef] [PubMed]
- Yoo, P.E.; Oxley, T.J.; Hagan, M.A.; John, S.; Ronayne, S.M.; Rind, G.S.; Brinded, A.M.; Opie, N.L.; Moffat, B.A.; Wong, Y.T. Distinct Neural Correlates Underlie Inhibitory Mechanisms of Motor Inhibition and Motor Imagery Restraint. Front. Behav. Neurosci. 2020, 14, 77. [Google Scholar] [CrossRef] [PubMed]
- Calton, J.L.; Taube, J.S. Where am I and how will I get there from here? A role for posterior parietal cortex in the integration of spatial information and route planning. Neurobiol. Learn. Mem. 2009, 91, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Head, H.; Holmes, G. Sensory disturbances from cerebral lesions. Brain 1911, 34, 102–254. [Google Scholar] [CrossRef]
- Whitlock, J.R. Posterior parietal cortex. Curr. Biol. 2017, 27, R691–R695. [Google Scholar] [CrossRef] [PubMed]
- Bazanova, O.M.; Kondratenko, A.V.; Kuzminova, O.I.; Muravlyova, K.B.; Petrova, S.E. EEG alpha indices depending on the menstrual cycle phase and salivary progesterone level. Hum. Physiol. 2014, 40, 140–148. [Google Scholar] [CrossRef]
- Solís-Ortíz, S.; Campos, R.G.; Félix, J.; Obregón, O. Coincident frequencies and relative phases among brain activity and hormonal signals. Behav. Brain Funct. 2009, 5, 18. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| w/m | Age | Traumatic/Non-t. | ASIA Score | Lesion | Complete/Incomplete | Age of Injury | Group | |
|---|---|---|---|---|---|---|---|---|
| 1 | w | 62 | NT | B | Th10 | C | 5 m | PNS |
| 2 | w | 46 | T | C | L1 | IC | 36 y | PNS |
| 3 | w | 31 | T | A | Th5 | C | 5 y | PNS |
| 4 | m | 20 | NT | A | Th10 | C | 2 m | PNS |
| 5 | m | 60 | T | D | C4 | IC | 5 y | PNS |
| 6 | m | 49 | T | D | C6 | IC | 4 y | PNS |
| 7 | m | 57 | T | A | Th12 | C | 13 y | PNS |
| 8 | w | 45 | T | B | C7 | IC | 7 y | PS |
| 9 | w | 48 | T | C | L1 | IC | 28 y | PS |
| 10 | m | 70 | T | D | L2 | IC | 5 m | PS |
| 11 | m | 25 | NT | A | T10 | C | 25 y | PS |
| 12 | m | 47 | T | A | C6 | C | 18 y | PS |
| 13 | w | 61 | T | A | Th8 | C | 6 y | PS |
| 14 | w | 52 | T | A | Th12 | C | 36 y | PS |
| 15 | w | 32 | T | A | Th7 | C | 17 y | PS |
| 16 | m | 42 | T | A | Th9 | C | 25 y | PS |
| 17 | m | 47 | T | A | Th5 | C | 25 y | PS |
| 18 | m | 54 | T | B | C6 | IC | 37y | PS |
| 19 | m | 51 | T | A | L2 | C | 33 y | PS |
| 20 | m | 66 | NT | C | L3 | IC | 7 y | PS |
| 21 | m | 55 | NT | A | L3 | C | 31 y | PS |
| 22 | w | 40 | T | B | L2 | IC | 18 y | PS |
| 23 | m | 30 | T | A | L5 | C | 10 y | PS |
| Test Statistic | df1 | df2 | p-Value | |
|---|---|---|---|---|
| frequency | 5.383 | 1 | Inf | 0.02 |
| source | 2.749 | 4.17 | Inf | 0.025 |
| frequency × source | 3.811 | 4.27 | Inf | 0.003 |
| sink | 7.752 | 5.789 | Inf | <0.001 |
| frequency × sink | 5.726 | 6.869 | Inf | <0.001 |
| source × sink | 11.349 | 9.69 | Inf | <0.001 |
| condition | 4.451 | 1 | Inf | 0.035 |
| frequency × source × condition | 2.534 | 4.099 | Inf | 0.037 |
| source × condition | 3.225 | 3.756 | Inf | 0.014 |
| sink × sound | 2.083 | 6.312 | Inf | 0.048 |
| sink × condition | 11.02 | 4.567 | Inf | <0.001 |
| frequency × sink × condition | 8.443 | 5.758 | Inf | <0.001 |
| source × sink × condition | 11.573 | 10.718 | Inf | <0.001 |
| frequency × source × sink x condition | 2.899 | 10.922 | Inf | 0.001 |
| group × sport group × frequency × sound | 4.582 | 1 | Inf | 0.032 |
| frequency × condition | 13.432 | 1 | Inf | <0.001 |
| group × sport group × sink × condition | 2.872 | 4.567 | Inf | 0.017 |
| group × sport group × frequency × sink × condition | 2.288 | 5.758 | Inf | 0.035 |
| group × sport group × source × condition | 2.454 | 3.756 | Inf | 0.047 |
| group × sport group × source × sink × sound | 1.844 | 12.54 | Inf | 0.033 |
| Source | Sink | Condition | Sound | Frequency | F-Value | df1 | df2 | p-Value | p Adj |
|---|---|---|---|---|---|---|---|---|---|
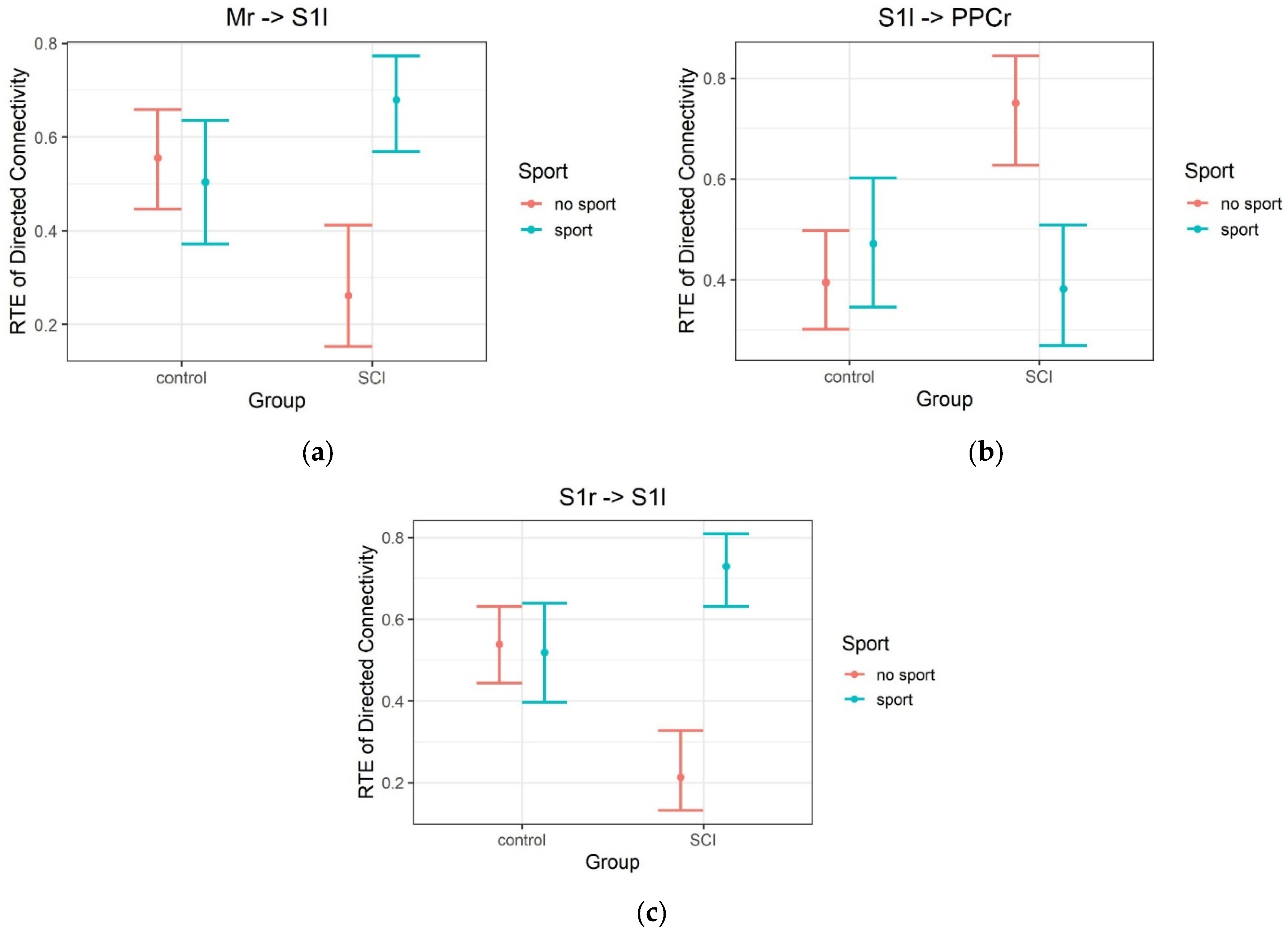
| S1l | PPCr | imagination | beep | alpha | 12.53 | 1 | 46.64 | 0.001 | 0.669 |
| Ml | PPCr | imagination | beep | alpha | 12.32 | 1 | 37.59 | 0.001 | 0.762 |
| S1l | Ml | imagination | beep | alpha | 8.75 | 1 | 47.64 | 0.005 | 1 |
| SMAr | S1l | imagination | beep | alpha | 7.88 | 1 | 46.54 | 0.007 | 1 |
| PMCl | Ml | imagination | beep | alpha | 7.62 | 1 | 48.07 | 0.008 | 1 |
| S1r | S1l | imagination | music | alpha | 20.9 | 1 | 39.44 | <0.001 | 0.137 |
| S1l | PPCr | imagination | music | alpha | 11.4 | 1 | 41.97 | 0.002 | 0.844 |
| Mr | S1l | imagination | music | alpha | 11.56 | 1 | 37.98 | 0.002 | 0.844 |
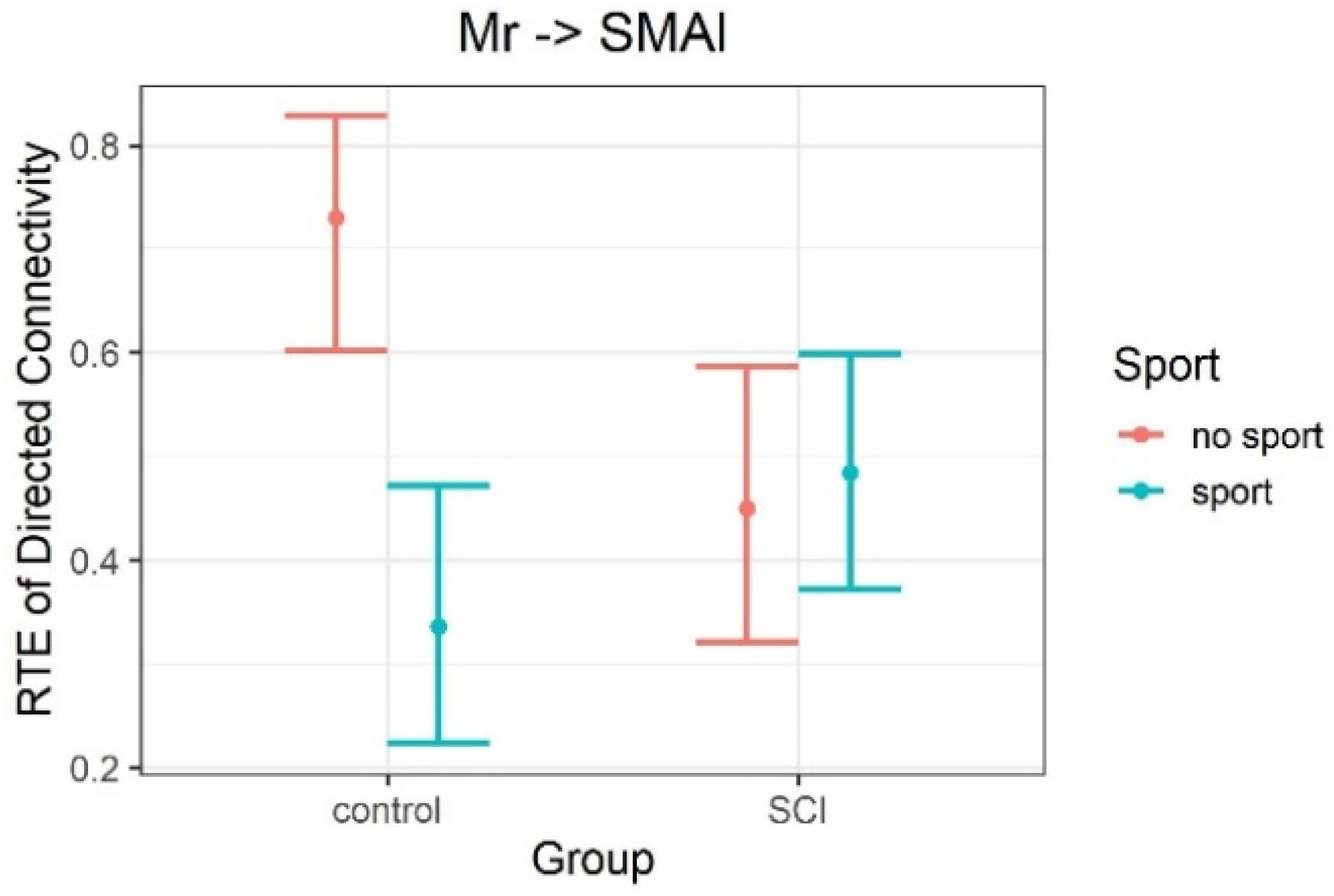
| Mr | SMAl | movement | music | alpha | 9.33 | 1 | 43.05 | 0.004 | 1 |
| Ml | S1r | imagination | music | beta | 19.09 | 1 | 39.96 | <0.001 | 0.167 |
| PMCl | S1r | imagination | music | beta | 15.74 | 1 | 42.47 | <0.001 | 0.283 |
| S1l | S1r | imagination | music | beta | 15.96 | 1 | 37.46 | <0.001 | 0.283 |
| S1l | Ml | imagination | music | beta | 10.28 | 1 | 47.66 | 0.002 | 1 |
| PMCl | Ml | imagination | music | beta | 10.21 | 1 | 39.41 | 0.003 | 1 |
| S1l | Ml | imagination | beep | beta | 27.94 | 1 | 45.24 | <0.001 | 0.02 |
| Ml | Ml | imagination | beep | beta | 16.58 | 1 | 47.56 | <0.001 | 0.254 |
| PMCl | Ml | imagination | beep | beta | 14.59 | 1 | 45.59 | <0.001 | 0.334 |
| PMCr | Mr | movement | beep | beta | 10.49 | 1 | 39.49 | 0.002 | 1 |
| S1l | PMCr | movement | beep | beta | 8.24 | 1 | 45.87 | 0.006 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frey, V.N.; Renz, N.; Thomschewski, A.; Langthaler, P.B.; van Schalkwijk, F.J.; Trinka, E.; Höller, Y. Influence of Sports on Cortical Connectivity in Patients with Spinal Cord Injury-A High-Density EEG Study. Appl. Sci. 2023, 13, 9469. https://doi.org/10.3390/app13169469
Frey VN, Renz N, Thomschewski A, Langthaler PB, van Schalkwijk FJ, Trinka E, Höller Y. Influence of Sports on Cortical Connectivity in Patients with Spinal Cord Injury-A High-Density EEG Study. Applied Sciences. 2023; 13(16):9469. https://doi.org/10.3390/app13169469
Chicago/Turabian StyleFrey, Vanessa N., Nora Renz, Aljoscha Thomschewski, Patrick B. Langthaler, Frank J. van Schalkwijk, Eugen Trinka, and Yvonne Höller. 2023. "Influence of Sports on Cortical Connectivity in Patients with Spinal Cord Injury-A High-Density EEG Study" Applied Sciences 13, no. 16: 9469. https://doi.org/10.3390/app13169469
APA StyleFrey, V. N., Renz, N., Thomschewski, A., Langthaler, P. B., van Schalkwijk, F. J., Trinka, E., & Höller, Y. (2023). Influence of Sports on Cortical Connectivity in Patients with Spinal Cord Injury-A High-Density EEG Study. Applied Sciences, 13(16), 9469. https://doi.org/10.3390/app13169469