
Assessment of the Accumulation of Trace Metals and Oxidative Stress Response Biomarkers in the Portunid Portunus segnis

Abstract
:1. Introduction
2. Materials and Methods
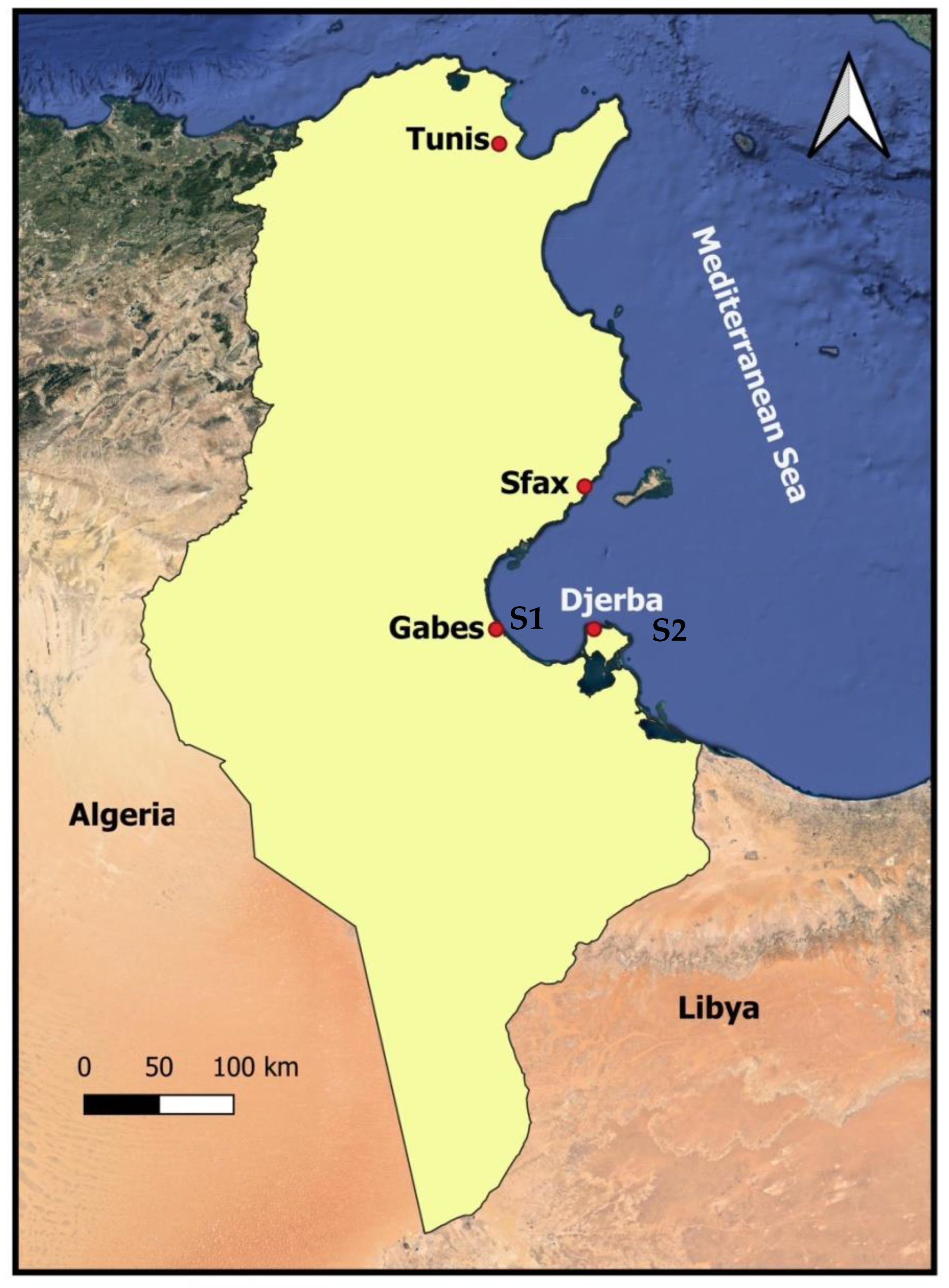
2.1. Presentation of the Studied Sites
2.2. Selection of Sampling Sites
- -
- Ghannouch (respective geographical coordinates, 33°55′ N and 10°5′ E) is the area considered the most polluted due to the establishment of the chemical group, suggesting that the marine ecosystem and therefore the fauna and flora are threatened by liquid, solid, and gaseous industrial discharges [20].
- -
- Sidi Salem (Djerba), the island of Djerba, is characterized by seasonal mass tourism, and the main sources of pollution are still anthropogenic waste.
2.3. Chemical Analysis
2.4. Biomarker Analyses
2.4.1. Tissue Preparation
2.4.2. Total Protein Quantification
2.4.3. Glutathione S-Transferase (GST) Determination
2.4.4. Catalase (CAT) Determination
2.4.5. Reduced Glutathione (GSH) Determination
2.4.6. Malonedialdehyde Level (MDA) Determination
2.5. Statistics
3. Results and Discussion
3.1. Metal Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Site | Tissues | Cu | Zn | Cd | Pb | Reference |
|---|---|---|---|---|---|---|---|
| Scylla serrata | Canada | Digestive Gland | 637 | 208 | 3.1 | - | [59] |
| Pseudocarcinus gigas | Australia | Digestive Gland | 52 | 108 | 2.2 | - | [62] |
| Menippemer scenario | South Carolina-United States | Digestive Gland | 308 | 46 | 4.6 | - | [63] |
| Ucidescordatus | Brazil | Digestive Gland | 6.6 | 8.89 | 0.16 | - | [64] |
| Xenograpsustestudinatus | Taiwan | Whole tissue | 53–290 | 119–610 | 0.49–1.29 | 1.83–2.64 | [65] |
| Eriocheir sinensis | Poland | Whole tissue | 3.17–21.9 | 1.15–26.6 | 0.016–0.025 | 0.29–0.61 | [60] |
| Carcinusmaenas | Bizerte Lagoon, Tunisia | Gills | 5.12–28.98 | 5.39–13.78 | 1.08–1.99 | 5.82–8.75 | [13] |
| Digestive Gland | 15.16–31.93 | 5.3–25.11 | 0.67–2 | 5.01–8.56 | |||
| Carcinusmaenas | Bizerte Lagoon, Tunisia | Gills | 857.3 | 627.1 | 0.61 | 21.1 | [39] |
| Monastir Coast, Tunisia | 376.5 | 83.4 | 0.38 | 15.57 | |||
| Sfax Coast, Tunisia | 238.9 | 88.8 | 0.52 | 4.94 | |||
| Bizerte Lagoon, Tunisia | Digestive Gland | 863.5 | 985.1 | 21.5 | 4.5 | ||
| Monastir Coast, Tunisia | 396.9 | 102.4 | 0.62 | 0.67 | |||
| Sfax Coast, Tunisia | 205.1 | 99.9 | 3.04 | 0.14 | |||
| Portunussegnis | Gulf of Gabès (Zarrat), Tunisia | Muscle | 206.45 | 590.04 | 0.21 | 0.4 | [6] |
| Gonad | 584.85 | 36.46 | 0.23 | 0.14 | |||
| Exoskeleton | 65.78 | 11.4 | 0.14 | 0.06 | |||
| Portunus pelagicus | Shuwaikh, Kuwait | Gills | 79.1 | 29.1 | 0.28 | 3.29 | [66] |
| Hepatopancreas | 68.6 | 18.9 | 0.43 | 4.69 | |||
| Chiromantes eulimene | Bhizolo Canal Richards Bay Harbour, South Africa | Gills | 106.74 | 219.8 | 9.72 | 23.24 | [67] |
| Digestive Gland | 153.6 | 140.57 | 7.12 | 8.94 | |||
| Scylla serrata | Punnakayal, Tuticorin, Southeast Coast of India | Gills | 3.63 | 14.26 | 0.14 | 0.19 | [68] |
| Hepatopancreas | 0.21 | 39.73 | 0.51 | 0.15 | |||
| Portunus pelagicus | Northern Bay of Bengal | Gills | 21.06 | - | 1.01 | 1.67 | [69] |
| Hepatopancreas | 12.46 | - | 2.82 | 2.07 | |||
| Cardisoma armatum | Kribi mangrove areas, Cameroon | Gills | 3.87 | 64.77 | 0.18 | 0.09 | [70] |
| Hepatopancreas | 7.88 | 51.34 | 0.35 | 2.07 | |||
| Portunus trituberculatus | Coast of Zhejiang Province, China | Gills | 35.1 | 16.4 | 0.2 | 0.08 | [71] |
| Hepatopancreas | 7.5 | 20.1 | 1.1 | 0.06 | |||
| Portunus segnis | Gulf of Gabès (Ghannouch), Tunisia | Gills | 12.53 | 37.36 | 19.21 | 10.15 | Present Study |
| Digestive Gland | 6.96 | 18.14 | 7.74 | 4.536 | |||
| Djerba Island, Tunisia | Gills | 6.45 | 10.72 | 7.2 | 4.9 | ||
| Digestive Gland | 2.91 | 8.71 | 3.18 | 2.21 |
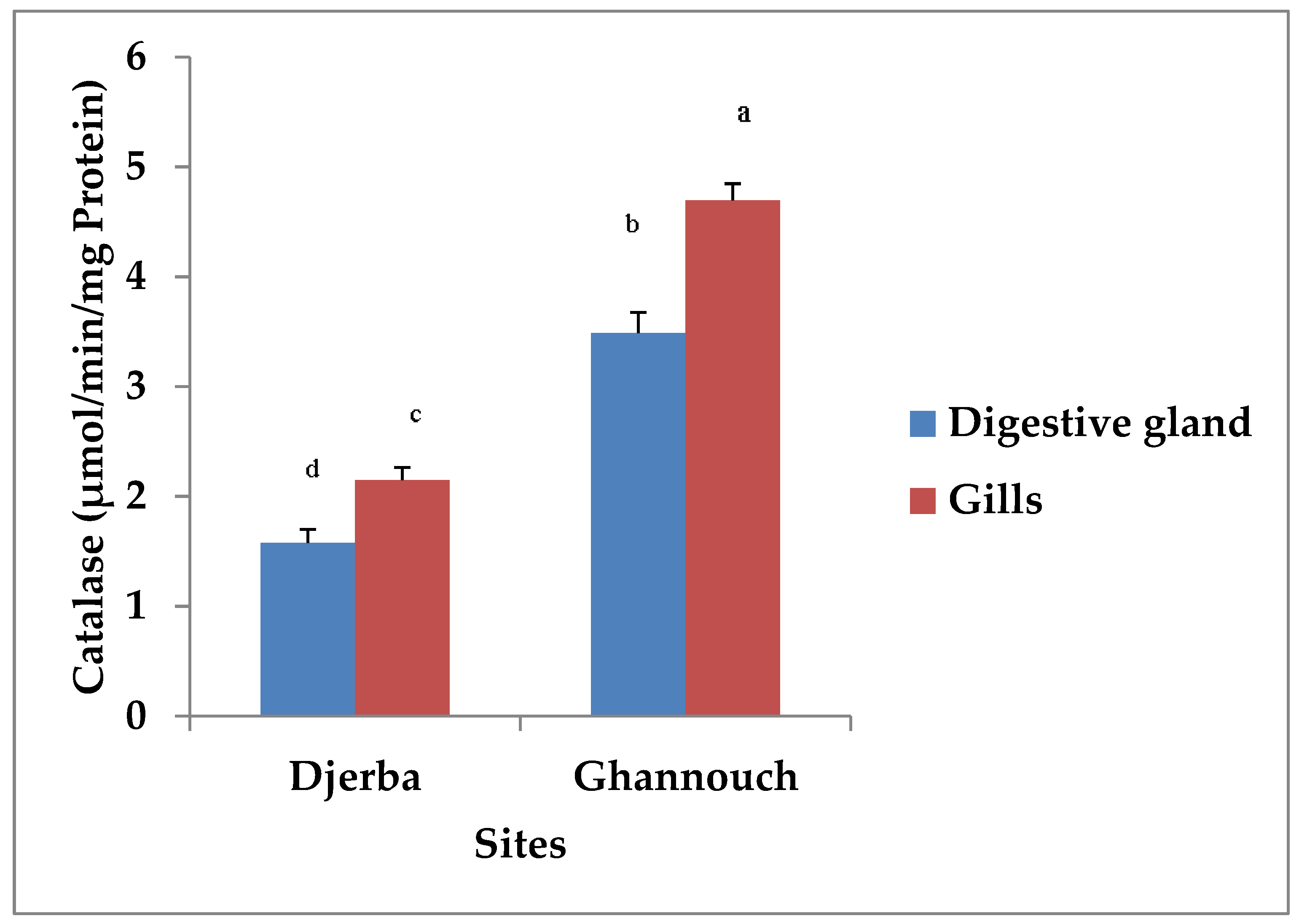
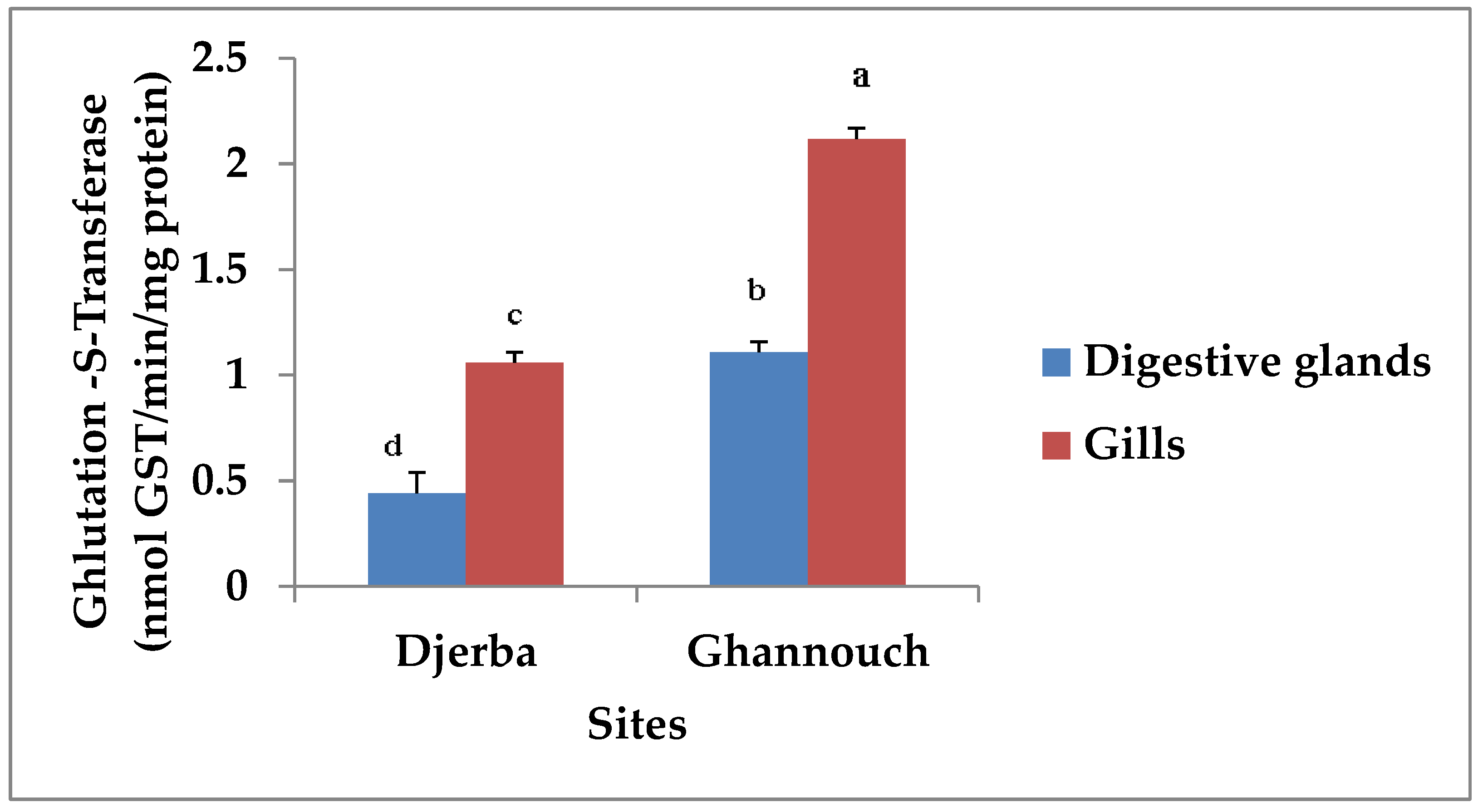
3.2. Biomarker Analysis
3.2.1. Catalase Activity
3.2.2. Glutathione S-Transferase Activity
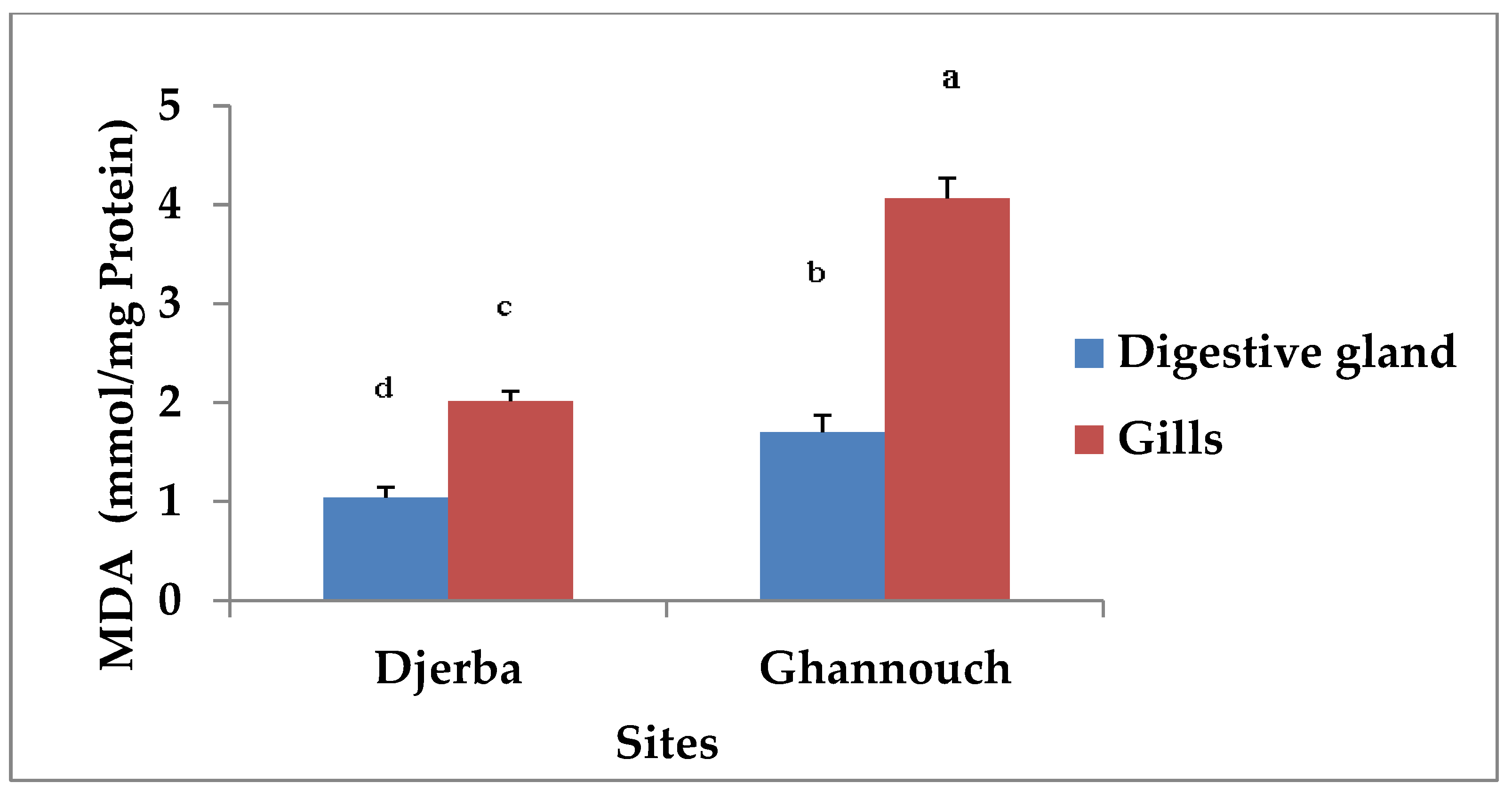
3.2.3. Malonedialdehyde Determination
3.2.4. Determination of Reduced Glutathione
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reddy, M.S.; Mehta, B.; Dave, S.; Joshi, M.; Karthikeyan, L.; Sarma, V.K.S.; Basha, S.; Ramachandraiah, G.; Bhatt, P. Bioaccumulation of heavy metals in some commercial fishes and crabs of the Gulf of Cambay. India Curr. Sci. 2007, 92, 1489–1491. [Google Scholar]
- Blaise, C.; Gagné, F.; Pellerin, J.; Hansen, P.-D.; Trottier, S. Molluscan Shellfish Biomarker Study of the Quebec, Canada, Saguenay Fjord with the Soft-Shell Clam, Mya arenaria: Molluscan Shellfish Biomarker StudyStudy. Environ. Toxicol. 2002, 17, 170–186. [Google Scholar] [CrossRef] [PubMed]
- Galloway, T.S.; Brown, R.J.; Browne, M.A.; Dissanayake, A.; Lowe, D.; Jones, M.B.; Depledge, M.H. A Multibiomarker Approach To Environmental Assessment. Environ. Sci. Technol. 2004, 38, 1723–1731. [Google Scholar] [CrossRef]
- Mayon, N.; Bertrand, A.; Leroy, D.; Malbrouck, C.; Mandiki, S.N.M.; Silvestre, F.; Goffart, A.; Thomé, J.-P.; Kestemont, P. Multiscale Approach of Fish Responses to Different Types of Environmental Contaminations: A Case Study. Sci. Total Environ. 2006, 367, 715–731. [Google Scholar] [CrossRef]
- UNEP/MAP. Draft Transboundary Diagnostic Analysis for the Mediterranean Sea, 1997 (TDA/MED) (UNEP(OCA)MED IG.ll/inf.7). Presented at MEDITERRANEAN ACTION PLAN Tenth Ordinary Meeting of the Contracting Parties to the Convention for the Protection of the Mediterranean Sea against Pollution and Its Protocols Tunis, Tunis, Tunisia, 18–21 November 1997. [Google Scholar]
- Hamza-Chaffai, A. Health Assessment of a Marine Bivalve Ruditapes decussatus from the Gulf of Gabès (Tunisia). Environ. Internat. 2003, 28, 609–617. [Google Scholar] [CrossRef]
- Smaoui-Damak, W.; Hamza-Chaffai, A.; Berthet, B.; Amiard, J.C. Preliminary Study of the Clam Ruditapes decussatus Exposed In Situ to Metal Contamination and Originating from the Gulf of Gabès, Tunisia. Bull. Environ. Contam Toxicol. 2003, 71, 961–970. [Google Scholar] [CrossRef] [PubMed]
- Rainbow, P.S. Trace Metal Concentrations in Aquatic Invertebrates: Why and so What? Environ. Pollut. 2002, 120, 497–507. [Google Scholar] [CrossRef]
- Fox, H.M. The Migration of a Red Sea Crab through the Suez Canal. Nature 1924, 113, 714–715. [Google Scholar] [CrossRef]
- Türeli, C.; Çelik, M.; Erdem, Ü. Comparison of meat composition and yield of blue crab (Callinectes sapidus RATHBUN, 1896) and sand crab (Portunus pelagicus Linne, 1758) caught in Iskenderun Bay, North-East Mediterranean. Turk. J. Vet. Anim. Sci. 2000, 24, 195–204. [Google Scholar]
- Rabaoui, L.; Arculeo, M.; Mansour, L.; Tlig-Zouari, S. Occurrence of the lessepsian species Portunus segnis (Crustacea: Decapda) in the Gulf of Gabés (Tunisia): First record and new information on its biology and ecology. Cah. Biol. Mar. 2015, 56, 169–175. [Google Scholar]
- Deidun, A.; Sciberras, A. A Further Record of the Blue Swimmer Crab Portunus segnis Forskal, 1775 (Decapoda: Brachyura: Portunidae) from the Maltese Islands (Central Mediterranean). BIR 2016, 5, 43–46. [Google Scholar] [CrossRef]
- Rifi, M.; Ounifi-BenAmor, K.; Ben Souissi, J.; Zaouali, J. Première mention du crabelessepsien Portunus segnis (Forskål, 1775) (Décapode, Brachyoure, Portunidae) dans les eaux marines Tunisiennes. In Proceedings of the du 4ème Congrès Franco-Maghrébin et 5èmes Journées Franco-Tunisiennes de Zoologie, Korba, Tunisie, 13–17 November 2014; p. 9. [Google Scholar]
- Bdioui, M. Premier signalement du crabe bleu Portunus segnis (Forskal, 1775) dans le Sud du Golfe de Hammamet (Centre-Est de la Tunisie). Bull. Inst. Natn. Scien. Tech. Mer de Salammbô. 2016, 43, 183–187. [Google Scholar]
- Ounifi-Ben Amor, K.; Rifi, Μ.; Ghanem, R.; Draeif, I.; Zaouali, J.; Ben Souissi, J. Update of Alien Fauna and New Records from Tunisian Marine Waters. Medit. Mar. Sci. 2015, 17, 124. [Google Scholar] [CrossRef]
- Burollet, P.F.; Clairefond, P.; Winnock, E. Géologie méditerranéenne, Tome VI, numéro 1. In La Mer Pélagienne: Etude Sédimentologique et écologique du Plateau Tunisien et du Golfe de Gabès; Département des sciences de la terre. Centre St Charles 13331 Marseille Cedex 3. Université de Provence: Marseille, France, 1979; p. 345. [Google Scholar]
- Hamza, A. Le Statut du Phytoplancton Dans le Golfe de Gabès. Ph.D. Thesis, Faculté des Sciences de Sfax, Sfax, Tunisie, 2003; p. 297. [Google Scholar]
- Drira, Z. Contribution à la Compréhension du Fonctionnement du Golfe de Gabès: Etude des Caractéristiques Dynamiques et Structurales des Communautés Phytozooplanctoniques. Ph.D. Thesis, Université de Sfax, Sfax, Tunisie, 2009; p. 229. [Google Scholar]
- Hattab, T.; Ben RaisLasram, F.; Albouy, C.; Romdhane, M.S.; Jarboui, O.; Halouani, G.; Cury, P.; Le Loch, F. An Ecosystem Model of an Exploited Southern Mediterranean Shelf Region (Gulf of Gabes, Tunisia) and a Comparison with Other Mediterranean Ecosystem Model Properties. J. Mar. Res. 2013, 128, 159–174. [Google Scholar] [CrossRef]
- Alaya-Ltifi, L.; Chokri, M.A.; Selmi, S. Breeding Performance of Passerines in a Polluted Oasis Habitat in Southern Tunisia. Ecotoxicol. Environ. Safe 2012, 79, 170–175. [Google Scholar] [CrossRef]
- Annabi, A.; Bardelli, R.; Vizzini, S.; Mancinelli, G. Baseline Assessment of Heavy Metals Content and Trophic Position of the Invasive Blue Swimming Crab Portunus segnis (Forskål, 1775) in the Gulf of Gabès (Tunisia). Mar. Pollut. Bull. 2018, 136, 454–463. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Almeida, J.R.; Gravato, C.; Guilhermino, L. Challenges in Assessing the Toxic Effects of Polycyclic Aromatic Hydrocarbons to Marine Organisms: A Case Study on the Acute Toxicity of Pyrene to the European Seabass (Dicentrarchus labrax L.). Chemosphere 2012, 86, 926–937. [Google Scholar] [CrossRef]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-Transferases. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Turdi, S.; Li, Q.; Lopez, F.L.; Ren, J. Catalase Alleviates Cardiomyocyte Dysfunction in Diabetes: Role of Akt, Forkhead Transcriptional Factor and Silent Information Regulator 2. Life Sci. 2007, 81, 895–905. [Google Scholar] [CrossRef]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L. Tissue Sulfhydryl Groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Catalano, D.; Torchia, G.; Pititto, F.; Greco, R. Contaminazione da metalli del “granchio americano”, Portunus pelagicus (Linnaeus, 1758) nella Rada di Augusta (Sicilia orientale). Biol. Mar. Mediterr. 2006, 13, 696–699. [Google Scholar]
- Ayas, D. The Effects of Season and Sex on the Nutritional Quality of Muscle Types of Blue Crab Callinectes sapidus (Rathbun, 1896) and Swimming Crab Portunus segnis (Forskal, 1775). NESciences 2016, 1, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Olgunoglu, M.; Olgunoglu, İ. Heavy Metal Contents in Blue Swimming Crab from the Northeastern Mediterranean Sea, Mersin Bay, Turkey. Pol. J. Environ. Stud. 2016, 25, 2233–2237. [Google Scholar] [CrossRef]
- Rainbow, P.S.; Furness, R.W. Heavy Metals in the Marine Environment. In Heavy Metals in the Marine Environment; Furness, R.W., Rainbow, P.S., Eds.; CRC Press: Boca Raton, FL, USA, 2018; pp. 1–4. ISBN 978135107315831. [Google Scholar]
- Gutiérrez, A.J.; Lozano, G.; Rubio, C.; Martín, V.; Hardisson, A.; Revert, C. Heavy Metals in Black Crabs in the Atlantic Coast (Tenerife, Spain)—Human Risk Assessment: Water. Clean Soil Air Water 2017, 45. [Google Scholar] [CrossRef]
- Nędzarek, A.; Czerniejewski, P.; Drost, A.; Harasimiuk, F.; Machula, S.; Tórz, A.; Masalski, P. The Distribution of Elements in the Body of Invasive Chinese Mitten Crabs (Eriocheir sinensis H. Milne-Edwards, 1853) from Lake Dąbie, Poland. J. Food Compos. Anal. 2017, 60, 1–9. [Google Scholar] [CrossRef]
- Turoczy, N.J.; Mitchell, B.D.; Levings, A.H.; Rajendram, V.S. Cadmium, Copper, Mercury, and Zinc Concentrations in Tissues of the King Crab (Pseudocarcinus gigas) from Southeast Australian Waters. Environ. Int. 2001, 27, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Reichmuth, J.M.; Weis, P.; Weis, J.S. Bioaccumulation and Depuration of Metals in Blue Crabs (Callinectes sapidus Rathbun) from a Contaminated and Clean Estuary. Environ. Pollut. 2010, 158, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Bondgaard, M.; Bjerregaard, P. Association between Cadmium and Calcium Uptake and Distribution during the Moult Cycle of Female Shore Crabs, Carcinus maenas: An In Vivo Study. Aquat. Toxicol. 2005, 72, 17–28. [Google Scholar] [CrossRef]
- Martín-Díaz, M.L.; Blasco, J.; Sales, D.; DelValls, T.A. Biomarkers Study for Sediment Quality Assessment in Spanish Ports Using the Crab Carcinus maenas and the Clam Ruditapes philippinarum. Arch. Environ. Contam Toxicol. 2007, 53, 66–76. [Google Scholar] [CrossRef]
- GB 5749-2006; Standards for Drinking Water Quality. Ministry of Health of the People’s Republic of China: Beijing, China, 2006.
- European Commission. Commission Regulation (EC) No 1881/2006 of the European parliament and the council of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Communities 2006, 18, L364. [Google Scholar]
- FAO. Compilation of legal limits for hazardous substances in fish and fishery products. FAO Fish Circ. 1983, 464, 5–100. Available online: http://trove.nla.gov.au/version/22206109 (accessed on 15 April 2023).
- Baden, S.P.; Eriksson, S.P. Role, routes and effects of manganese in crustaceans. Oceanogr. Mar. Biol. Annu. Rev. 2006, 44, 61–83. [Google Scholar]
- Mancinelli, G.; Papadia, P.; Ludovisi, A.; Migoni, D.; Bardelli, R.; Fanizzi, F.P.; Vizzini, S. Beyond the Mean: A Comparison of Trace- and Macroelement Correlation Profiles of Two Lacustrine Populations of the Crayfish Procambarus clarkii. Sci. Total Environ. 2018, 624, 1455–1466. [Google Scholar] [CrossRef]
- Amiard, J.; Amiardtriquet, C.; Barka, S.; Pellerin, J.; Rainbow, P. Metallothioneins in Aquatic Invertebrates: Their Role in Metal Detoxification and Their Use as Biomarkers. Aquat. Toxicol. 2006, 76, 160–202. [Google Scholar] [CrossRef]
- Schuwerack, P.-M.M.; Lewis, J.W.; Jones, P. The potential use of the South African river crab, Potamonauteswarreni, as a bioindicator species for heavy metal contamination. Ecotoxicology 2001, 10, 159–166. [Google Scholar] [CrossRef]
- Pan, L.; Zhang, H. Metallothionein, Antioxidant Enzymes and DNA Strand Breaks as Biomarkers of Cd Exposure in a Marine Crab, Charybdis Japonica. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2006, 144, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, J.D.; Da Silva, M.R.; Da Silva, A.C.B.; De Lima, S.M.A.; Malm, O.; Allodi, S. Tissue Distribution, Subcellular Localization and Endocrine Disruption Patterns Induced by Cr and Mn in the Crab Ucides Cordatus. Aquat. Toxicol. 2005, 73, 139–154. [Google Scholar] [CrossRef] [PubMed]
- Rainbow, P.S. Biomonitoring of trace metals in estuarine and marine environments. Aust. J. Ecotoxicol. 2006, 12, 107–122. [Google Scholar]
- Doherty, V.F.; Ogunkuade, O.O.; Kanife, U.C. Biomarkers of oxidative stress and heavy metal levels as indicators of environmental pollution in some selected fishes in Lagos, Nigeria. Am. Eurasian J. Agric. Envrion. Sci. 2010, 7, 359–365. [Google Scholar]
- Bjerregaard, P.; Depledge, M.H. Trace Metal Concentrations and Contents in the Tissues of the Shore Crab Carcinus maenas: Effects of Size and Tissue Hydration. Mar. Biol. 2002, 141, 741–752. [Google Scholar] [CrossRef]
- Peng, S.-H.; Hung, J.-J.; Hwang, J.-S. Bioaccumulation of Trace Metals in the Submarine Hydrothermal Vent Crab Xenograpsus testudinatus off Kueishan Island, Taiwan. Mar. Pollut. Bull. 2011, 63, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Ghedira, J.; Jebali, J.; Banni, M.; Chouba, L.; Boussetta, H.; López-Barea, J.; Alhama, J. Use of Oxidative Stress Biomarkers in Carcinus Maenas to Assess Littoral Zone Contamination in Tunisia. Aquat. Biol. 2011, 14, 87–98. [Google Scholar] [CrossRef] [Green Version]
- Ben-Khedher, S.; Jebali, J.; Houas, Z.; Nawéli, H.; Jrad, A.; Banni, M.; Boussetta, H. Metals Bioaccumulation and Histopathological Biomarkers in Carcinus maenas Crab from Bizerta Lagoon, Tunisia. Envrion. Sci. Pollut. Res. 2014, 21, 4343–4357. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, M.R. Pesticide and Trace Metal Concentrations in Queensland Estuarine Crabs. Mar. Pollut. Bull. 2000, 41, 359–366. [Google Scholar] [CrossRef]
- Reed, L.A.; Pennington, P.L.; Wirth, E. A Survey of Trace Element Distribution in Tissues of Stone Crabs (Menippe mercenaria) from South Carolina Coastal Waters. Mar. Pollut. Bull. 2010, 60, 2297–2302. [Google Scholar] [CrossRef]
- Pinheiro, M.A.A.; Silva, P.P.G.E.; Duarte, L.F.D.A.; Almeida, A.A.; Zanotto, F.P. Accumulation of Six Metals in the Mangrove Crab UcidesCordatus (Crustacea: Ucididae) and Its Food Source, the Red Mangrove Rhizophora Mangle (Angiosperma: Rhizophoraceae). Ecotoxicol. Environ. Safe 2012, 81, 114–121. [Google Scholar] [CrossRef]
- Bonanno, G.; Orlando-Bonaca, M. Trace Elements in Mediterranean Seagrasses: Accumulation, Tolerance and Biomonitoring. A Review. Mar. Pollut. Bull. 2017, 125, 8–18. [Google Scholar] [CrossRef]
- Ayadi, N.; Aloulou, F.; Bouzid, J. Assessment of Contaminated Sediment by Phosphate Fertilizer Industrial Waste Using Pollution Indices and Statistical Techniques in the Gulf of Gabes (Tunisia). Arab. J. Geosci. 2015, 8, 1755–1767. [Google Scholar] [CrossRef]
- El Zrelli, R.; Courjault-Radé, P.; Rabaoui, L.; Castet, S.; Michel, S.; Bejaoui, N. Heavy Metal Contamination and Ecological Risk Assessment in the Surface Sediments of the Coastal Area Surrounding the Industrial Complex of Gabes City, Gulf of Gabes, SE Tunisia. Mar. Pollut. Bull. 2015, 101, 922–929. [Google Scholar] [CrossRef] [PubMed]
- Jara-Marini, M.E.; Soto-Jiménez, M.F.; Páez-Osuna, F. Trophic Relationships and Transference of Cadmium, Copper, Lead and Zinc in a Subtropical Coastal Lagoon Food Web from SE Gulf of California. Chemosphere 2009, 77, 1366–1373. [Google Scholar] [CrossRef]
- Schneider, L.; Maher, W.A.; Potts, J.; Taylor, A.M.; Batley, G.E.; Krikowa, F.; Adamack, A.; Chariton, A.A.; Gruber, B. Trophic Transfer of Metals in a Seagrass Food Web: Bioaccumulation of Essential and Non-Essential Metals. Mar. Pollut. Bull. 2018, 131, 468–480. [Google Scholar] [CrossRef] [PubMed]
- El-Shenawy, N.S.; Mohammadden, A.; Al-Fahmie, Z.H. Using the Enzymatic and Non-Enzymatic Antioxidant Defense System of the Land Snail Eobania vermiculata as Biomarkers of Terrestrial Heavy Metal Pollution. Ecotoxicol. Environ. Safe 2012, 84, 347–354. [Google Scholar] [CrossRef]
- Marnett, L.J. Oxy Radicals, Lipid Peroxidation and DNA Damage. Toxicology 2002, 181–182, 219–222. [Google Scholar] [CrossRef]
- Barata, C.; Varo, I.; Navarro, J.C.; Arun, S.; Porte, C. Antioxidant Enzyme Activities and Lipid Peroxidation in the Freshwater Cladoceran daphnia magna Exposed to Redox Cycling Compounds. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2005, 140, 175–186. [Google Scholar] [CrossRef]
- Stegeman, J.J.; Marius, B.; Giulio, R.T.D.; Lars, F.; Fowler, B.A.; Sanders, B.Μ.; Veld, P.A.V. Molecular responses to environmental contamination: Enzyme and protein systems as indicators of chemical exposure and effect. In Biomarkers. Biochemical, Physiological and Histological Markers of Anthropogenic Stress; Huggett, R.J., Kimerle, R.A., Mehrle, P.M., Jr., Bergman, H.L., Eds.; Lewis Publishers: London, UK, 1992; pp. 235–335. [Google Scholar]
- Karam, Q.; Guermazi, W.; Subrahmanyam, M.N.V.; Al-Enezi, Y.; Ali, M.; Leignel, V.; Annabi-Trabelsi, N. Portunus pelagicus (Linnaeus, 1758) as a Sentinel Species to Assess Trace Metal Occurrence: A Case Study of Kuwait Waters (Northwestern Arabian Gulf). Toxics 2023, 11, 426. [Google Scholar] [CrossRef]
- Majola, N.; Mzimela, H.M.; Izegaegbe, J.I. Metal Bioaccumulation and Energy Biomarkers in Tissues of Two Populations of Chiromantes Eulimene from Richards Bay Harbour, South Africa. Sci. Afr. 2020, 10, e00558. [Google Scholar] [CrossRef]
- Yogeshwaran, A.; Gayathiri, K.; Muralisankar, T.; Gayathri, V.; Monica, J.I.; Rajaram, R.; Marimuthu, K.; Bhavan, P.S. Bioaccumulation of Heavy Metals, Antioxidants, and Metabolic Enzymes in the Crab Scylla serrata from Different Regions of Tuticorin, Southeast Coast of India. Mar. Pollut. Bull. 2020, 158, 111443. [Google Scholar] [CrossRef]
- Karar, S.; Hazra, S.; Das, S. Assessment of the Heavy Metal Accumulation in the Blue Swimmer Crab (Portunus pelagicus), Northern Bay of Bengal: Role of Salinity. Mar. Pollut. Bull. 2019, 143, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Ngo-Massou, V.M.; Kottè-Mapoko, E.F.; Din, N. Heavy Metal Accumulation in the Edible Crab Cardisoma armatum (Brachyura: Gecarcinidae) and Implications for Human Health Risks. Sci. Afr. 2022, 16, e01248. [Google Scholar] [CrossRef]
- Liu, Q.; Liao, Y.; Xu, X.; Shi, X.; Zeng, J.; Chen, Q.; Shou, L. Heavy Metal Concentrations in Tissues of Marine Fish and Crab Collected from the Middle Coast of Zhejiang Province, China. Environ. Monit. Assess 2020, 192, 285. [Google Scholar] [CrossRef] [PubMed]
- Jebali, J.; Banni, M.; Almeida, E.A.D.; Boussetta, H. Oxidative DNA Damage Levels and Catalase Activity in the Clam Ruditapes Decussatus as Pollution Biomarkers of Tunisian Marine Environment. Environ. Monit. Assess 2007, 124, 195–200. [Google Scholar] [CrossRef]
- Frías-Espericueta, M.G.; Bautista-Covarrubias, J.C.; Osuna-Martínez, C.C.; Delgado-Alvarez, C.; Bojórquez, C.; Aguilar-Juárez, M.; Roos-Muñoz, S.; Osuna-López, I.; Páez-Osuna, F. Metals and Oxidative Stress in Aquatic Decapod Crustaceans: A Review with Special Reference to Shrimp and Crabs. Aquat. Toxicol. 2022, 242, 106024. [Google Scholar] [CrossRef]
- De Jesus, W.B.; Mota Andrade, T.D.S.D.O.; Soares, S.H.; Pinheiro-Sousa, D.B.; De Oliveira, S.R.S.; Torres, H.S.; Protazio, G.D.S.; Da Silva, D.S.; Santos, D.M.S.; De Carvalho Neta, A.V.; et al. Biomarkers and Occurrences of Heavy Metals in Sediment and the Bioaccumulation of Metals in Crabs (Ucides cordatus) in Impacted Mangroves on the Amazon Coast, Brazil. Chemosphere 2021, 271, 129444. [Google Scholar] [CrossRef]
- Khessiba, P.; Hoarau, M.; Gnassia-Bar, A. Biochemical Response of the Mussel Mytilus galloprovincialis from Bizerta (Tunisia) to Chemical Pollutant Exposure. Arch. Environ. Contam. Toxicol. 2001, 40, 222–229. [Google Scholar] [CrossRef]
- Dellali, M.; GnassiaBarelli, M.; Romeo, M.; Aissa, P. The Use of Acetylcholinesterase Activity in Ruditapes Decussatus and Mytilus Galloprovincialis in the Biomonitoring of Bizerta Lagoon. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2001, 130, 227–235. [Google Scholar] [CrossRef]
- Jerome, F.C.; Hassan, A.; Omoniyi-Esan, G.O.; Odujoko, O.O.; Chukwuka, A.V. Metal Uptake, Oxidative Stress and Histopathological Alterations in Gills and Hepatopancreas of Callinectes amnicola Exposed to Industrial Effluent. Ecotoxicol. Environ. Safe 2017, 139, 179–193. [Google Scholar] [CrossRef]
- Géret, F. Influence of Metal Exposure on Metallothionein Synthesis and Lipid Peroxidation in Two Bivalve Mollusks: The Oyster (Crassostrea gigas) and the Mussel (Mytilus edulis). Aquat. Living Resour. 2002, 15, 61–66. [Google Scholar] [CrossRef]
- Bonneris, E.; Perceval, O.; Masson, S.; Hare, L.; Campbell, P.G.C. Sub-Cellular Partitioning of Cd, Cu and Zn in Tissues of Indigenous Unionid Bivalves Living along a Metal Exposure Gradient and Links to Metal-Induced Effects. Environ. Pollut. 2005, 135, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Bebianno, M.; Company, R.; Serafim, A.; Camus, L.; Cosson, R.; Fialamedoni, A. Antioxidant Systems and Lipid Peroxidation in from Mid-Atlantic Ridge Hydrothermal Vent Fields. Aquat. Toxicol. 2005, 75, 354–373. [Google Scholar] [CrossRef] [PubMed]
- Ferrat, L.; Pergent-Martini, C.; Roméo, M. Assessment of the Use of Biomarkers in Aquatic Plants for the Evaluation of Environmental Quality: Application to Seagrasses. Aquat. Toxicol. 2003, 65, 187–204. [Google Scholar] [CrossRef] [PubMed]
- Al-Mutairi, D.A.; Craik, J.D.; Batinic-Haberle, I.; Benov, L.T. Induction of Oxidative Cell Damage by Photo-Treatment with Zinc MetaN -Methylpyridylporphyrin. Free Radic. Res. 2007, 41, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Pampanin, D.M.; Camus, L.; Gomiero, A.; Marangon, I.; Volpato, E.; Nasci, C. Susceptibility to Oxidative Stress of Mussels (Mytilus galloprovincialis) in the Venice Lagoon (Italy). Mar. Pollut. Bull. 2005, 50, 1548–1557. [Google Scholar] [CrossRef] [PubMed]
- Giarratano, E.; Gil, M.N.; Malanga, G. Biomarkers of Environmental Stress in Gills of Ribbed Mussel Aulacomya atra atra (Nuevo Gulf, Northern Patagonia). Ecotoxicol. Environ. Safe 2014, 107, 111–119. [Google Scholar] [CrossRef]
- Fahmy, S.R.; Abdel-Ghaffar, F.; Bakry, F.A.; Sayed, D.A. Ecotoxicological Effect of Sublethal Exposure to Zinc Oxide Nanoparticles on Freshwater Snail Biomphalaria alexandrina. Arch. Environ. Contam. Toxicol. 2014, 67, 192–202. [Google Scholar] [CrossRef]
- Radwan, M.A.; El-Gendy, K.S.; Gad, A.F. Biomarkers of Oxidative Stress in the Land Snail, Theba Pisana for Assessing Ecotoxicological Effects of Urban Metal Pollution. Chemosphere 2010, 79, 40–46. [Google Scholar] [CrossRef]
- Jacob, L. L’insuffisance Rénale Aiguë; Springer: Paris, France, 2007; pp. 1–342. [Google Scholar] [CrossRef]
- Schwab, M. Encyclopedia of Cancer; Springer: Berlin/Heidelberg, Germany, 2011. [Google Scholar] [CrossRef]
- Morgan, J. Evidence on the Developmental and Reproductive Toxicity of Metribuzin. DRAFT, California, 2001; p. 65. Available online: https://oehha.ca.gov/media/downloads/crnr/metribuzin.pdf (accessed on 15 April 2023).
- Pelletier, É.; Campbell, P.G.C.; Denizeau, F. Écotoxicologie Moléculaire: Principes Fondamentaux et Perspectives de Développement, 1st ed.; Presses de l’Université du Québec: Québec, QC, Canada, 2004; pp. 1–502. [Google Scholar] [CrossRef]
- Poortmans, J.R.; Boisseau, N. Biochimie des Activités Physiques, 2nd ed.; De BoeckSupérieur: Bruxelles, Belgium, 2003; p. 480. [Google Scholar]
- Taleb-Senouci, D.; Ghomari, H.; Krouf, D.; Bouderbala, S.; Prost, J.; Lacaille-Dubois, M.A.; Bouchenak, M. Antioxidant Effect of Ajuga Iva Aqueous Extract in Streptozotocin-Induced Diabetic Rats. Phytomedicine 2009, 16, 623–631. [Google Scholar] [CrossRef]
- Haq, Q.; Jan, A.; Ali, A. Glutathione as an Antioxidant in Inorganic Mercury Induced Nephrotoxicity. J. Postgrad. Med. 2011, 57, 72. [Google Scholar] [CrossRef]
- Jozefczak, M.; Remans, T.; Vangronsveld, J.; Cuypers, A. Glutathione Is a Key Player in Metal-Induced Oxidative Stress Defenses. Int. J. Mol. Sci. 2012, 13, 3145–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hore, M.; Saha, R.; Bhaskar, S.; Mandal, S.; Bhattacharyya, S.; Roy, S. Oxidative Stress Responses in Puntius sarana Collected from Some Environmentally Contaminated Areas of River Mahananda, Malda, West Bengal. Ecotoxicology 2023, 32, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Singaram, G.; Harikrishnan, T.; Chen, F.-Y.; Bo, J.; Giesy, J.P. Modulation of Immune-Associated Parameters and Antioxidant Responses in the Crab (Scylla serrata) Exposed to Mercury. Chemosphere 2013, 90, 917–928. [Google Scholar] [CrossRef] [PubMed]
- Leomanni, A.; Schettino, T.; Calisi, A.; Gorbi, S.; Mezzelani, M.; Regoli, F.; Lionetto, M.G. Antioxidant and Oxidative Stress Related Responses in the Mediterranean Land Snail Cantareus Apertus Exposed to the Carbamate Pesticide Carbaryl. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2015, 168, 20–27. [Google Scholar] [CrossRef] [PubMed]

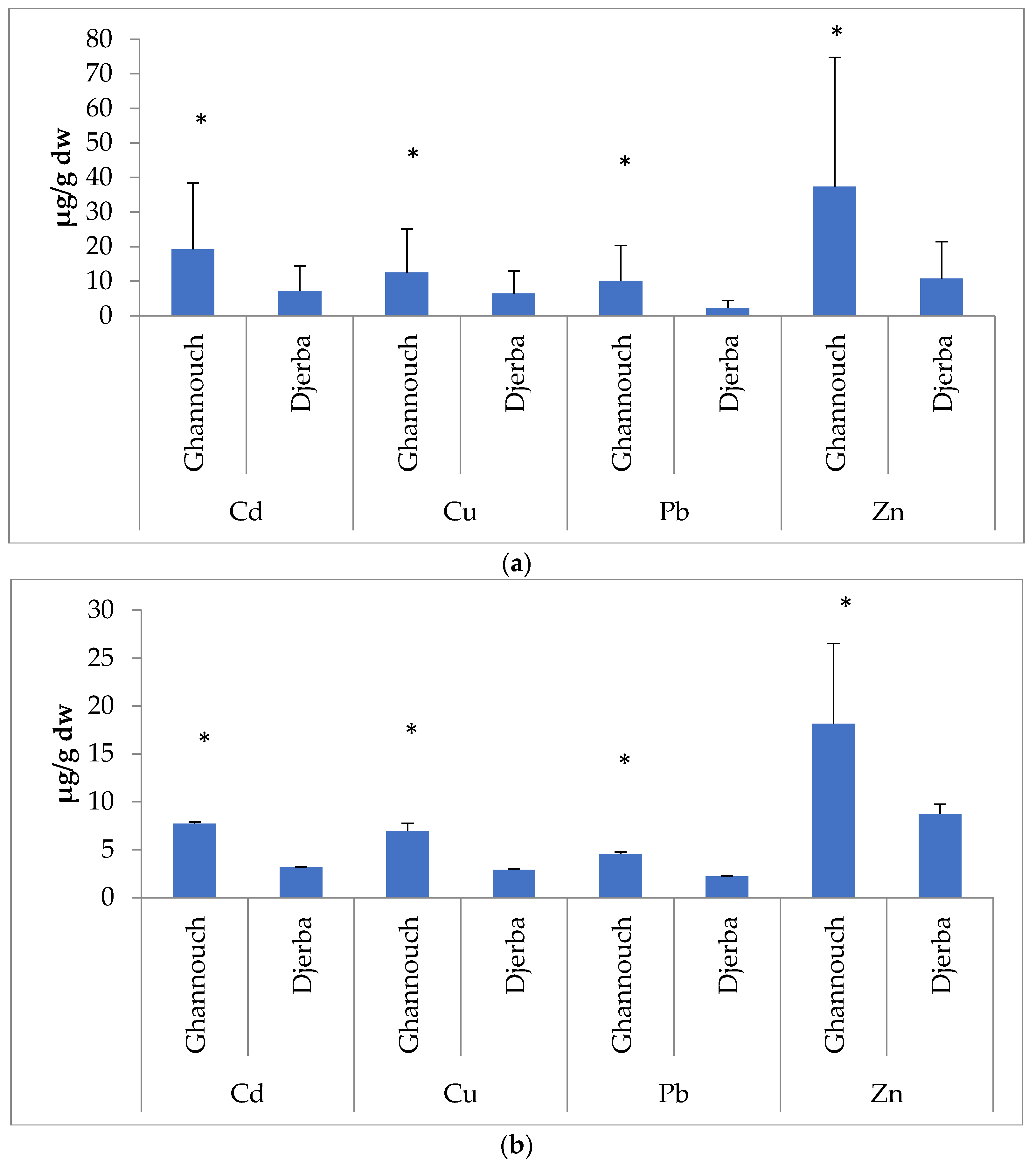

| Site | Tissues | Element | Mean ± SD | F | p |
|---|---|---|---|---|---|
| Ghannouch | Gills | Cd | 19.21 ± 0.1 | 42.28 | <0.01 |
| Cu | 12.53 ± 0.078 | ||||
| Pb | 10.15 ± 1.38 | ||||
| Zn | 37.36 ± 12.57 | ||||
| Hepatopancreas | Cd | 7.74 ± 0.14 | 72.57 | <0.01 | |
| Cu | 6.96 ± 0.79 | ||||
| Pb | 4.536 ± 0.24 | ||||
| Zn | 18.14 ± 8.38 | ||||
| Djerba | Gills | Cd | 7.2 ± 0.049 | 20.31 | <0.01 |
| Cu | 6.45 ± 0.12 | ||||
| Pb | 4.9 ± 0.01 | ||||
| Zn | 10.72 ± 2.9 | ||||
| Hepatopancreas | Cd | 3.18 ± 0.04 | 15.12 | <0.01 | |
| Cu | 2.91 ± 0.08 | ||||
| Pb | 2.21 ± 0.05 | ||||
| Zn | 8.71 ± 1.03 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dghim, A.; Ben Ameur, W.; Annabi, A. Assessment of the Accumulation of Trace Metals and Oxidative Stress Response Biomarkers in the Portunid Portunus segnis. Appl. Sci. 2023, 13, 7197. https://doi.org/10.3390/app13127197
Dghim A, Ben Ameur W, Annabi A. Assessment of the Accumulation of Trace Metals and Oxidative Stress Response Biomarkers in the Portunid Portunus segnis. Applied Sciences. 2023; 13(12):7197. https://doi.org/10.3390/app13127197
Chicago/Turabian StyleDghim, Amel, Walid Ben Ameur, and Ali Annabi. 2023. "Assessment of the Accumulation of Trace Metals and Oxidative Stress Response Biomarkers in the Portunid Portunus segnis" Applied Sciences 13, no. 12: 7197. https://doi.org/10.3390/app13127197
APA StyleDghim, A., Ben Ameur, W., & Annabi, A. (2023). Assessment of the Accumulation of Trace Metals and Oxidative Stress Response Biomarkers in the Portunid Portunus segnis. Applied Sciences, 13(12), 7197. https://doi.org/10.3390/app13127197