Abstract
Bacterial cellulose (BC) is a natural exopolysaccharide characterized by a high purity, mechanical strength, and the ability to absorb various compounds. Obtaining BC on an industrial scale is expensive, mainly due to the high cost of the culture media. Replacing classical media with waste from the agri-food industry may be a promising way to reduce the costs when obtaining BC. The aim of these studies was to compare the effectiveness of the biosynthesis of BC in classical and waste media using two strains, Komagataeibacter xylinus K2G30 and Gluconacetobacter hansenii ATCC 23769. Classical Hestrin−Schramm, YPM, GY, and GYE media were used. The waste media were banana peels, celery pomace, fish waste, hemp seed pomace, deproteinized acid whey (DAW), and non-deproteinized acid whey (NDAW). The highest efficiency of BC biosynthesis in classical media was observed in YPM and it was 5.23 g·dm−3 with K. xylinus K2G30. In the waste media, the highest yield of BC was observed in AW, when the yield of BC was 2.99 g·dm−3 with K. xylinus K2G30. In addition, it was demonstrated that the presence of proteins and lactic acid in AW influenced the efficiency when obtaining BC in the culture of K. xylinus. The results indicate that it is possible to obtain BC from industrial waste, especially from AW.
1. Introduction
The global agri-food industry generates about 1.3 billion tons of waste per year as a result of processing, among others, fruit, vegetables, dairy products, bakery products, and meat [1]. The forecasts for the generation of municipal food waste indicate an increase from 278 million in 2005 to 416 million in 2025, which is a cause of great concern [1]. The increase in the amount of environmentally hazardous waste, as well as economic and legal considerations, cause food processing companies to increasingly employ modern methods for reducing waste production and seek ways to reuse it [2]. As a result, more and more industries are trying to adapt to the circular economy model. According to the assumptions of this system, waste that is a by-product of one process can become a valuable substrate material in another process [3,4]. Waste generated by the agri-food industry is a source of many nutrients such as proteins, polysaccharides, triacylglycerols, fats, and phenolic compounds [5]. One way of using by-products from the agri-food industry is to subject them to various biotransformations involving microorganisms, for example, to biosynthesis new compounds and substances with added value [2]. For example, in recent years, orange peels have been successfully used for the biosynthesis of xanthan and curdlan using Xanthamonas campestris and Alcaligenes faecalis, respectively [6,7]. In turn, watermelon peels have been used as a source of nutrients for Saccharomyces cerevisiae in the process of bioethanol production [8]. Lactic acid is also obtained from industrial by-products through microbial biosynthesis. Lactic acid has been successfully produced from potato peels, green pea peels, orange peels, and mango peels using Lacticaseibacillus casei and Lb. delbrueckii [9].
Acetic acid bacteria (AAB) have been known to humans for many decades [10]. The most characteristic feature of AAB is the ability to oxidize ethanol to acetic acid; thus, it is widely used for vinegar production. This reaction is catalyzed by two enzymes: alcohol dehydrogenase (which converts ethanol into acetaldehyde) and aldehyde dehydrogenase (which oxidizes acetaldehyde into acetic acid) [11]. AAB also produce a number of other metabolites such as gluconic acid, dihydroxyacetone, acetate, or bacterial cellulose. In the industrial production of gluconic acid, Gluconobacter is the most commonly used genus, as in the production of dihydroxyacetone. Acetan is mainly produced by the Acetobacter and Komagataeibacter genera. Bacterial cellulose is biosynthesized with the greatest efficiency mainly by the Komagataeibacter xylinus [12].
Bacterial cellulose ((BC) biocellulose) is an exopolymer build from glucose units linked by 1,4-β-glycosidic bonds. The β-glycosidic chains then combine into subfibrils and microfibrils forming fibers, which imparts a fibrous structure to BC [13]. Despite its similar structure to plant cellulose, biocellulose is not contaminated with lignins and hemicelluloses. Thus, its purification and further use is relatively inexpensive and environmentally friendly [14]. The unique properties of BC, such as high crystallinity, porosity, and mechanical strength, make it suitable for many applications [15]. It has gained the greatest popularity in biomedicine, where it is used, for example, as a wound dressing material or as scaffolds for tissue engineering [14,16,17]. In food products, it may be used as a stabilizer, emulsifier, fat replacer, or meat analogue [13,18]. Biodegradable food packaging based on biocellulose is becoming increasingly popular [19]. In addition, biocomponents from BC can be used in the protection of the environment in water purification membranes and in cosmetics as carriers of active substances or as stabilizers of cosmetic structures [20,21,22].
The main limitations for the use of BC on a large scale include the high costs of the media necessary for the cultivation of microorganisms. One of the justifiable solutions to this problem seems to be the search for cheaper substitutes for classical media that would ensure a comparable or higher efficiency of biocellulose biosynthesis. Such substitutes may be post-production waste from the agri-food industry [23]. So far, the use of certain waste types, such as citrus peels, watermelon peels, pineapple peels, potatoes peels, or wheat broth, has exerted a positive impact on the process of obtaining BC through increasing the reaction efficiency, shortening the process time, and reducing its costs [22,23,24]. Although there is still a high demand for new knowledge in this field, it is all the more justified to continue such research. Acid whey (AW) is one of the main by-products of the dairy industry resulting from the coagulation of casein during the production of, for example, cottage cheese, ricotta, and Greek yoghurt [25]. Because of the presence of organic compounds such as lactose, proteins, lipids, organic acids, or minerals, acid whey has a high chemical oxygen demand (COD) ranging from 50 g·dm−3 to 70 g·dm−3 and biochemical oxygen demand (BOD) from 27 g·dm−3 to 60 g·dm−3, which hinders its disposal [26]. However, the processes of whey heat treatment, including deproteinization and use in microbiological processes, may reduce COD and BOD by up to 60% [27].
The aim of the study was to attempt to replace classical media for BC biosynthesis with waste media generated in the agri-food industry. In the first stage, two strains of AAB (Gluconacetobacter hansenii ATCC 23769 and Komagataeibacter xylinus K2G30) were cultured in classical (Hestrin−Schramm, YPM, GY, and GYE) and waste media (banana peels, celery pomace, fish waste, hemp seed pomace, and acid whey). Based on the results from this stage, the strain and waste media that provided the highest BC yield were selected. In the second stage, the selected strain was cultured and the changes in the composition of the waste medium (before and after de-proteinization) during the culture were analyzed, and the obtained BC was characterized.
2. Materials and Methods
2.1. Bacterial Strains
In the research considered, two strains of AAB were used, namely Komagataeibacter xylinus K2G30 (University of Modena and Reggio Emilia Microbial Culture Collection, Reggio Emilia, Italy) and Gluconacetobacter hansenii ATCC 23769 (American Type Culture Collection, Manassas, VA, USA).
- First stage of study.
2.2. Media Composition
Classical media:
- Hestrin−Schramm (HS) [g·dm−3]: glucose 20, peptone 5, yeast extract 5, sodium hydrogen phosphate (Na2HPO4) 2.7, citric acid 1.15, pH 5.0;
- YPM [g·dm−3]: yeast extract 5, mannitol 25, peptone 3, pH 5.75;
- GY [g·dm−3]: glucose 100, yeast extract 10, pH 4.7;
- GYE [g·dm−3]: glucose 30, yeast extract 30, ethanol 20, pH 5.5.
The media components were purchased from the Biomaxima Company, Lublin, Poland. The media were sterilized at 121 °C for 20 min.
Waste media:
- Banana peels (obtained from a household; after peeling the bananas, the peels were ground for 2 min with a blender; Tefal Quickchef HB656838, Warsaw, Poland);
- Celery pomace (from a household; the celery pomace was obtained after pressing the juice with the low-speed juicer; Kuvings, Wasilków, Poland);
- Fish waste (Imperial-Kołobrzeg Company, Warsaw, Poland);
- Hemp seed pomace (Kropla Omega oil mill, Starachowice, Poland);
- Acid whey (Laktopol Dairy Plant, Łosice, Poland).
All media, except whey, were supplemented with yeast extract at a concentration of 10 g·dm−3 (Biomaxima, Lublin, Poland). The concentration of organic waste in the media was 20, 50, or 70 g·dm−3, while other concentrations (10, 15, and 20 g·dm−3) were used in the hemp seed pomace media to more accurately distribute and homogenize the waste in the media. The media were sterilized at 121 °C for 20 min. The composition of the waste media used is provided in Table 1.

Table 1.
Composition of waste media.
2.3. Inoculum Preparation
First, 100 cm3 of GY medium was inoculated with 5 cm3 of K. xylinus K2G30 suspension and incubated with shaking (110 rpm) for 48 h at 28 °C [28]. The preparation of the inoculum of the bacteria Ga. hansenii ATCC 23769 was conducted in an analogous way, with the only difference being the use of the YPM medium in place of GY.
2.4. Biosynthesis and Purification of Bacterial Cellulose
The classical and waste media for BC biosynthesis were inoculated with 10% (v/v) of an appropriate inoculum of AAB. Stationary culture was then carried out for 14 days at 28 °C. After this period, the cellulose was removed, cleaned by rinsing with distilled water several times, and incubated in 0.1 M NaOH (Chempur, Piekary Śląskie Poland) for 2 h at 90 °C to eliminate bacterial cells and medium residues. The cellulose was then dried at 45 °C for 5 h until a constant mass was achieved and then weighed [20].
Based on the BC mass obtained in the first stage of the study, the strain showing the highest BC biosynthesis efficiency and the waste medium ensuring the receipt of the highest BC mass were selected for the second stage of the study.
- Second stage of study.
2.5. AW Deproteinization
After the first stage of research, AW with a concentration of 70 g·dm−3 was selected as the best waste medium for the second stage. The concentrated and powdered AW was reconstituted in sterile distilled water. The medium prepared this way was deproteinized using the chemo-thermal method. The pH of the media was determined using 2M NaOH and/or 37% HCl (Chempur, Piekary Śląskie, Poland) to 5.0, 6.0, 7.0, and 7.5 (media labelled D5, D6, D7, and D7.5 respectively). The media were heated at 92 °C for 20 min. The precipitated proteins were separated from the whey by centrifugation (6850× g, 10 min) [24]. The supernatant was decanted from and a pH of 5.0 was determined. After that, the media were sterilized at 121 °C for 20 min. Then, it was used as a deproteinized medium for BC biosynthesis. At the same time, cultures were carried out in the medium of non-deproteinized acid whey (NDAW).
2.6. Characterization of AW Media
The waste media made of NDAW and deproteinized AW were characterized in terms of the content of reducing sugars, protein and non-protein nitrogen content, calcium and magnesium content, lactic acid content, and pH changes. All of the analyses were performed both before and after conducting BC biosynthesis. The content of reducing sugars was determined using the 3,5-dinitrosalicylic acid (DNS) (Pol-Aura, Warsaw, Poland) [29]. The content of calcium and magnesium in whey was determined using the complexometric method using ethylenediaminetetraacetic acid (EDTA) with calconacarboxylic acid (Pol-Aura, Warsaw, Poland) as an indicator [30]. The level of protein nitrogen (PN) and non-protein nitrogen (NPN) was determined using the Kjeldahl method, in accordance with PN-EN ISO 8968-4 [31]. The lactic acid content was tested using the titration method [32].
2.7. BC Biosynthesis on Whey Media
Here, 50 cm3 of the prepared waste medium on the basis of deproteinized (D5, D6, D7, and D7.5) AW and NDAW was inoculated with a centrifuged (5000 rpm, 10 min) (MPV-350R centrifuge, MPV Ltd. Gliwice Poland) inoculum of Komagataeibacter xylinus K2G30 in the amount of 0.5 g. A stationary culture to obtain BC was conducted for 14 days at 28 °C. The obtained cellulose was then purified and dried as described in point 2.4.
2.8. Attenuated Total Reflactance-Fourier Transform Infrared (ATR-FTIR) Characterization of Bacterial Cellulose
FTIR spectrometry was used to identify the chemical structure of the BC obtained in the NDAW medium and four variants of deproteinized AW media. The BC obtained in the YPM medium was used as the control BC. The ATR-FTIR spectra were acquired by means of a Nicolet iS20 spectrophotometer (Thermo Scientific, Waltham, MA, USA), in the range between 4000 and 600 cm−1, at 4 cm−1 resolution for 32 scans. The ATR-FTIR air background spectrum was collected every 30 min. The spectra were normalized to 1.0 at 1030 cm−1 (COH streching vibration). Baseline corrections were obtained on Omnic Software (Thermo Scientific, Waltham, MA, USA). Before analysis, the samples were conditioned for 24 h at constant conditions of 23 ± 1 °C and 50 ± 1% humidity.
2.9. Statistical Analysis
All of the determinations were performed three times. The standard deviations are shown in the charts. The method of one-factor analysis of variance (ANOVA) was employed for the statistical analysis of the obtained results. To compare the differences between the media, Tukey’s range test with a 95% confidence level was used. For the analysis of the media composition before and after culturing, the t-test of the dependent samples with a significance level of 95% was employed.
3. Results and Discussion
- First stage of study.
3.1. BC Biosynthesis in Classical Media
The obtained results showed that in the case of the Ga. hansenii strain, there were no statistically significant differences in BC biosynthesis on different classical media. For the K. xylinus strain, HS and YPM media differed significantly from GY and GYE media, where BC production was low. In the HS and YPM media, a significant difference was observed in the BC mass obtained by each strain. In both media, more BC mass was obtained through the culture of K. xylinus than Ga. hansenii. No statistically significant differences were observed between the mass of BC produced by Ga. hansenii and K. xylinus in both the GY and GYE media. After culturing the Ga. hansenii and K. xylinus strains in classical media, it was found that the GY and YPM media ensured obtaining the highest BC yields (Figure 1). In the YPM medium, a higher yield of BC (5.23 g·dm−3) was obtained from the culture of the K. xylinus strain in comparison with the yield of BC obtained from the culture of Ga. hansenii in the GY medium (3.19 g·dm−3). The HS medium is widely considered to be one of the most versatile and optimized media for obtaining BC [30]. However, in the presented research, the cultures of AAB in the HS medium resulted in obtaining a lower BC mass (1.34 g·dm−3 for Ga. hansenii and 4.17 g·dm−3 for K. xylinus) compared with the cultures in the GY and YPM media, respectively. Therefore, it can be assumed that the efficiency of BC production in the HS medium depends on the strain used [33]. In this study, a low BC yield was obtained in the GYE medium for both of the strains tested (Figure 1).
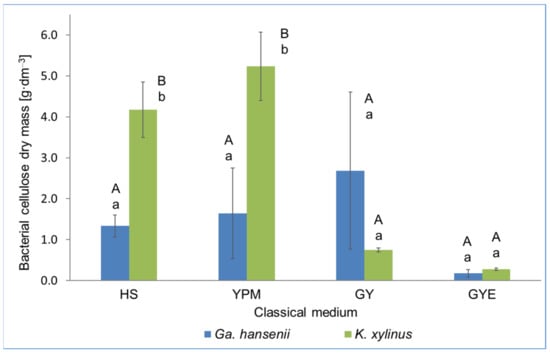
Figure 1.
Dry mass of bacterial cellulose obtained in classical media by Ga. hansenii and K. xylinus. The same uppercase letters in the same strain for different media are not statistically significantly different. The same lowercase letters in the same medium for different strains are not statistically significantly different.
Presumably, the presence of ethyl alcohol in the GYE medium led to inducing AAB metabolism towards its oxidation to acetic acid, instead of using glucose for cell proliferation, which had a direct impact on the low yield of BC biosynthesis [32]. The YPM medium contains mannitol as the carbon source, as well as peptone and yeast extract as the nitrogen source. Glucose is the source of carbon in the HS medium, while nitrogen is supplied through the yeast extract and peptone [28]. For AAB, mannitol might be a better carbon source than glucose, because mannitol metabolism gluconic acid is not produced (as opposed to glucose metabolism), which affects the pH medium changes, which in turn reduces the activity of AAB cells and has an impact on the disturbance of BC biosynthesis [31]. Gullo et al. [28] also observed a higher efficiency of BC biosynthesis in the mannitol medium compared with the glucose medium (8.8 g·dm−3 and 6.2 g·dm−3, respectively). In the case of the GY medium, a clear, but not statistically significant difference between the mass of BC produced by Ga. hansenii and K. xylinus was noted and 2.7 g·dm−3 and 0.7 g·dm−3, respectively. This is probably because the K. xylinus strain requires a more complex and nutrient-rich medium to produce BC. For example, the media in which K. xylinus produced the largest amount of BC (HS and YPM), other than with glucose and yeast extract, were enriched with, among others, peptone, mannitol, Na2HPO4, and citric acid. The low BC yield in the GY medium could be explained by the fact that instead of BC, glucose in this medium could be transformed into gluconic acid and its derivatives [34]. As noted by Liu et al. [35], two energy production pathways exist in AAB: one is the production of phosphates at the substrate level by glucose metabolism, and the other involves the oxidative production of phosphates by the reduced form of nicotinamide adenine dinucleotide (NADH) (or reduced flavin adenine dinucleotide (FADH2)) generated in the process of glucose metabolism and transferred to electron acceptors [35]. In the case of glucose availability, it was phosphorylated to glucose-6-phosphate (G6P) through glucokinase. Subsequently, G6P was transformed into 6-phosphogluconolactone (GlcL6P), which was related to an energy gain for the cell as this process was accompanied by a reduction in NADP to NADPH. However, in the presence of organic acids in the medium, e.g., citric acid (HS medium), they are included into the tricarboxylic acid (TCA) cycle to generate energy (in the form of ATP) and promote cell growth [34]. In turn, the higher content of ATP supports the production of BC in the pathway, which inhibits the activity of glucose-6-phosphate dehydrogenase, converting G6P into GlcL6P [35]. Presumably, the generation of energy in TCA processes results in the reduction of NADP to NADPH, which accompanies the transformation of G6P into GlcL6P, unnecessary; therefore, G6P can be incorporated into the BC biosynthesis pathway, increasing its yield.
3.2. BC Biosynthesis on Waste Media
The effectiveness of biocellulose synthesis by Ga. hansenii ATCC 23769 and K. xylinus K2G30 strains in various waste media was assessed. The obtained results showed that within the same medium, there were no significant differences between waste concentrations for most media (exception of banana peels and celery pomace). Statistically significant differences occurred within the same concentrations for different waste media. This means that for obtaining BC by Ga. hansenii, the type of waste was more important than its concentration. The largest yield of BC produced by Ga. hansenii ATCC 23769 was obtained in media with celery pomace with a concentration of 20 g·dm−3 (1.79 g·dm−3) and banana peels with a concentration of 70 g·dm−3 (1.53 g·dm−3; Figure 2).
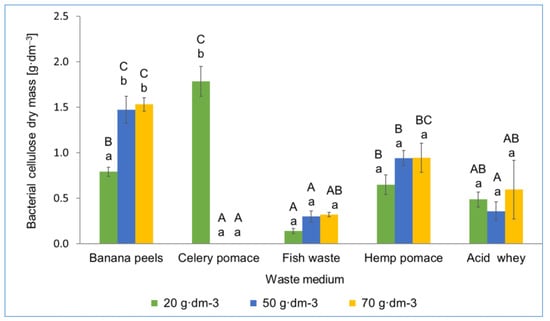
Figure 2.
Dry mass of bacterial cellulose obtained on waste media using Ga. hansenii ATCC 23769. The same uppercase letters in the same concentration for different waste media are not statistically significantly different. The same lowercase letters in the same waste media for different concentration are not statistically significantly different.
The obtained BC weight values were slightly higher compared with the results obtained in the studies by Bozdağ et al. [28] and Moukamnerd et al. [29], where in media based on hydrolysates from celery root and banana peels, 1.08 g·dm−3 and 0.89 g·dm−3 of BC were obtained by K. hansenii DSM 5602 and K. nataicola TISTR 2661, respectively [36,37]. The BC obtained in the media with celery waste at concentrations of 50 g·dm−3 and 70 g·dm−3 was so delicate and unstable that it underwent significant damage during its removal from the medium (Figure 3). Therefore, its purification and determination were not possible. BC from these medium variants was characterized by numerous embedded celery fibers, which probably reduced its mechanical strength. Similar observations were reached by Pommet et al. [38], who analyzed BC obtained in a medium with hemp fibers and observed that biocellulose was characterized by a low value of Young’s modulus and low mechanical strength. In celery pomace medium with a concentration of 20 g·dm−3, the most BC was obtained; thus, the use of this waste in the future would require one to consider reducing the concentration of waste or applying a pre-treatment, e.g., acid/enzymatic hydrolysis, which would reduce the incorporation of plant fibers into the BC structure. In the medium with fish waste, the smallest yield of BC was obtained, regardless of the concentration used. Fish waste is rich in protein, organic carbon, nitrogen, and phosphorus, but has a low sugar content, which could possibly limit BC biosynthesis [39]. According to current knowledge, this was the first study in which fish waste had been used and described for producing BC. However, there are reports in the literature on fish waste use in the microbiological biosynthesis of other compounds, e.g., lactic acid [40]. In the studies by Gao et al. [40], it was demonstrated that the largest yield of lactic acid was obtained after the initial treatment and acid hydrolysis of fish waste. It was established that the bacteria Lactobacillus rhamnosus were not able to absorb protein nutrients without their prior hydrolysis, which also influenced the uptake of, e.g., glucose. On this basis, it can be assumed that, as well as in our study, the lack of hydrolysis of fish waste negatively affected the production of BC by Ga. hansenii. Hemp pomace was also used as a waste medium (concentrations of 10, 15, and 20 g·dm−3) for BC biosynthesis. The lowest yield of BC was obtained in a medium with a concentration of 10 g·dm−3; however, the BC yield did not differ significantly from the BC yield obtained in the media with a concentration of 15 g·dm−3 and 20 g·dm−3. On the other hand, the BC yield produced by Ga. hansenii in the medium of AW reached approx. 0.5 g·dm−3, regardless of the concentration used. Most AAB exhibit a limited ability to absorb the lactose present in whey. This is caused by the lack or low activity of the β-galactosidase enzyme responsible for the hydrolysis of lactose to simple sugars [41]. Thus, the biosynthesis of BC by AAB in whey may be impeded due to the lower availability of glucose, which is the basic structural unit of cellulose.

Figure 3.
Bacterial cellulose obtained by Ga. hansenii ATCC 23769 and K. xylinus K2G30 on waste media: banana peels, celery pomace, fish waste, hemp pomace, and acid whey.
Figure 4 shows the BC yield obtained after the cultivation of K. xylinus in waste media. Statistical analysis showed that, as for the strain Ga. hansenii, when obtaining BC by K. xylinus, the type of waste was more important than its concentration.
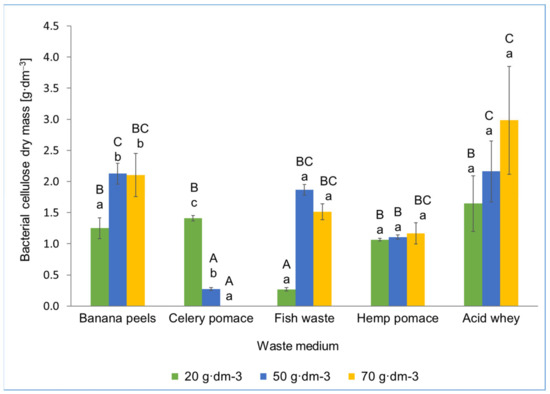
Figure 4.
Dry mass of bacterial cellulose obtained on waste media using K. xylinus K2G30. The same uppercase letters at the same concentration for different waste media are not statistically significantly different. The same lowercase letters at the same waste media for different concentrations are not statistically significantly different.
In the medium with banana peels with a concentration of 20 g·dm−3, the BC yield was about 1.4 g·dm−3. In the same medium, but with a concentration of 50 g·dm−3 and 70 g·dm−3, the yield of BC was 2.1 g·dm−3. After culturing K. xylinus in the media of celery pomace, 1.4 g·dm−3 (waste concentration of 20 g·dm −3) and 0.3 g·dm−3 (waste concentration of 50 g·dm−3) BC were obtained, respectively. It is worth mentioning that this is a surprising result because, as in the case of Ga. hansenii, the most BC was obtained at the lowest waste concentration (20 g·dm−3). This could have been due to the high content of phenolic compounds present in celery. As reported by Pastorkova et al. and Yao and Ren [42,43], luteolin, coumaric acid, and ferulic acid present in celery had an inhibitory effect on the growth of acetic bacteria. In media with 50 g·dm−3 and 70 g·dm−3 celery, the content of phenolic compounds was high enough to inhibit the growth of Ga. hansenii and K. xylinus, which prevented the production of BC. For fish waste, the highest BC yield (1.85 g·dm−3) was obtained in the medium with a concentration of 50 g·dm−3. This may indicate that 5% of fish waste was best suited for BC biosynthesis. The fish waste medium with a concentration of 20 g·dm−3 was probably too poor in nutrients to ensure high efficiency of BC biosynthesis, and thus significantly less biopolymer was obtained (0.3 g·dm−3). In turn, in the fish waste medium with a concentration of 70 g·dm−3 K. xylinus produced 1.5 g·dm−3 BC, which could have been caused by too high a concentration and inhibitory effect of some components of fish waste on the biosynthesis of BC. As reported by Coppola et al. [44], a negative impact of peptide hydrolysates from fish waste on the growth of some of the Gram-negative bacteria was demonstrated [44]. However, it should be noted that K. xylinus demonstrated a better ability to produce BC from fish waste compared with Ga. hansenii. This may indicate that K. xylinus is better adapted to BC biosynthesis under unfavourable conditions than Ga. hansenii. It can be assumed that the presence of, e.g., hydrolysates of peptides from fish waste in the medium inhibits BC biosynthesis by Ga. hansenii, while for K. xylinus, it is not largely significant. In the case of the medium from hemp waste, K. xylinus generated about 1.1 g·dm−3, regardless of the applied concentration of waste (10, 15, or 20 g·dm−3). This result was similar to the mass of BC produced by Ga. hansenii, of about 0.94 g·dm−3. The greatest quantity of BC was obtained in an AW medium (Figure 4) with a concentration of 70 g·dm−3; the BC mass was then about 3 g·dm−3. This was the highest BC mass obtained by K. xylinus among all of the waste media and accounted for 57% of the BC mass obtained on the best classical medium (YPM, 5.23 g·dm−3). This result was similar to the mass of BC obtained under similar conditions in the studies of other authors [45,46]. Salari et al. [47] obtained 3.55 g·dm−3 of BC; however, in this study, sweet whey (from rennet cheese production) was subjected to a preliminary enzymatic treatment for hydrolysing lactose. The hydrolysis of lactose to glucose and galactose might be important for BC biosynthesis, because AAB would thus have greater access to glucose, which is the basic building block of BC [47,48].
The first stage of the study shows that the K. xylinus strain had a higher BC biosynthetic efficiency than Ga. hansenii in both the classical and waste media (Table 2). On this basis, it was selected and used in the next stage of the study. Among the classical media, the highest BC production was provided by GY and YPM media for Ga. hansenii and K. xylius, respectively. More than 1.5 times higher efficiency of BC biosynthesis in the YPM medium resulted in it being selected as the control medium in the second stage of the study. In the case of waste media, celery pomace medium at 20 g·dm−3 provided the highest BC production efficiency for the strain Ga. hansenii and AW medium with a concentration of 70 g·dm−3 for K. xylinus. On the basis of the BC mass obtained in these media, the medium with AW was considered the best and was selected for further studies. In the second stage of the research, AW was modified by deproteinization under various conditions and it was verified whether the content of whey proteins affected the efficiency of BC production. Purification of BC was carried out in the same way as in the first part of the study.
Table 2.
Summary of classical and waste media providing the highest BC.
- Second stage of study.
3.3. Characterization of AW Media
Lactose is the main reducing sugar in AW; in addition to this, AW also contains small amounts of glucose, galactose, and arabinose [47]. Depending on the source of AW (curd cheese, Greek yoghurt, or casein acid coagulation), its composition may vary, which also affects the lactose content, which can range from 33 mg·mL−1 to 49 mg·mL−1 [25]. The analysis of reducing sugars in the whey demonstrated that their content was in accordance with the standards given in the scientific literature [25]. The highest content of reducing sugars, amounting to about 45 g·dm−3, was found in the medium of NDAW, while in the media of deproteinized whey, the content of reducing sugars ranged from about 33 mg·mL−1 to about 40 mg·mL−1 (Figure 5). The highest consumption of sugars during the cultivation of the bacterium K. xylinus K2G30 was observed in the medium of NDAW, which corresponded to the highest mass of BC obtained. In turn, the lowest consumption of reducing sugars, amounting to about 0.7 g·dm−3, was observed in the D7 medium, which was also consistent with the lowest BC mass obtained in this medium (Figure 9). The content of reducing sugars in the media of deproteinized AW and NDAW is shown in Figure 5.
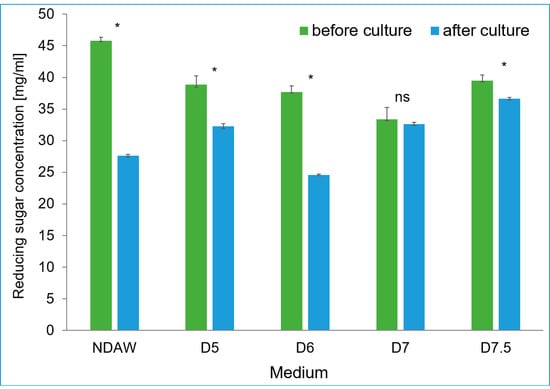
Figure 5.
The content of reducing sugars in NDAW and deproteinized media during culture of K. xylinus K2G30. * significant difference in the medium component before and after culture; p-value 0.05. ns non-significant difference.
Deproteinization of AW at 90 °C in an environment of different pH (5.0, 6.0, 7.0, and 7.5) resulted in a reduction in the protein nitrogen content in the deproteinized whey media compared with the nitrogen content in NDAW. It was observed that as the pH increased, the protein nitrogen content decreased, which was consistent with the initial assumptions based on the literature data [49]. The lowest quantity of protein nitrogen was quantified in the medium of deproteinized AW under the most alkaline conditions—pH 7.5. According to Chandrapala et al. [49], the pH level increase during whey deproteinating from pH 3.0 to pH 10.0 and an increase in temperature from 15 °C to 90 °C caused a decrease in the ratio of whey proteins to total proteins. The authors explained this phenomenon by the fact that a high pH (e.g., pH 10.0) increased the tendency to deamidation (hydrolysis reaction where a water molecule is added to an amide releasing ammonia), which resulted in an increase in the intermolecular repulsion forces, which led to the decorrugation of the protein and, as a result, to the formation of large protein aggregates. These aggregates, in turn, could be removed from the whey by centrifugation [49]. The initial content of protein nitrogen in NDAW was 0.019%, and the lowest protein nitrogen was recorded in the D7.5 medium—approx. 0.005% (Figure 6). After conducting BC biosynthesis, the largest use of protein nitrogen was found in NDAW (from 0.019% to 0.008%), which was consistent with obtaining the largest amount of BC produced in this medium (Figure 9). A statistically insignificant consumption of protein nitrogen was observed in the media D7 and D7.5, in which the protein nitrogen content reached about 0.006% and in which the least cellulose was produced (Figure 9). The protein nitrogen content in NDAW and deproteinized whey media is shown in Figure 6.
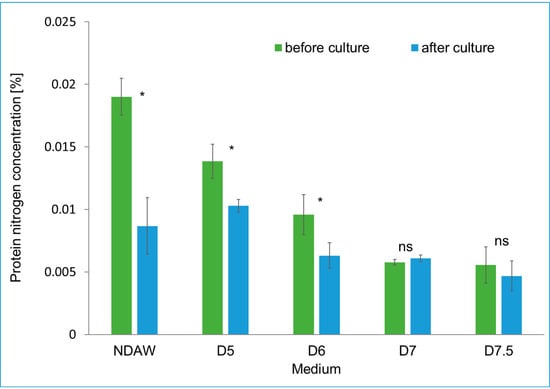
Figure 6.
The content of protein nitrogen in NDAW and deproteinized AW media during the culture of K. xylinus K2G30. * significant difference in the medium component before and after culture, p-value 0.05. ns non-significant difference.
The deproteinization of AW had little impact on the content of non-protein nitrogen (NPN). The highest initial NPN content was observed in the NDAW medium (0.047%) and the lowest in the D7.5 medium (0.038%). After culturing K. xylinus, a significant difference between the content of non-protein nitrogen was observed only in the medium of NDAW. Before the culture, the content of NPN in this medium was 0.047% and decreased after culturing to 0.044%, which corresponded to the consumption of 40 mg·dm−3 and the production of 1.57 g·dm−3 BC in this medium. In other media, the consumption of NPN was statistically insignificant, which in turn did not affect the amount of BC obtained. For comparative purposes, in the study by Lappa et al. [50] involving BC biosynthesis by A. xylinum DSM 15973, 0.58 g·dm−3 of biopolymer was obtained in the medium of deproteinized whey using 19 mg·dm−3 free amino nitrogen (FAN) and a total consumption of about 50 g·dm−3 sugar. However, in that study, the initial FAN content was 56 mg·dm−3 [50]. It is presumed that free amino acids may have had the greatest impact on the production of BC from among the low molecular weight compounds included in whey NPN, such as urea, ammonia, uric acid, free amino acids, creatine, creatinine, and biogenic amines [51]. Aspartic acid, phenylalanine, and serine have recently been recognized as indispensable amino acids for BC biosynthesis [52]. On the other hand, in other studies, methionine is also considered an important amino acid for BC biosynthesis [53]. According to Heo and Son, [54] the addition of arginine and threonine to the medium only slightly increased the yield from BC biosynthesis [54]. It is worth noting that all the amino acids mentioned above were present in AW at concentrations from 20 g·kg−1 total protein in the case of methionine to even 100 g·kg−1 total protein for aspartic acid [55]. The non-protein nitrogen content in the NDAW and deproteinized whey media is presented in Figure 7.
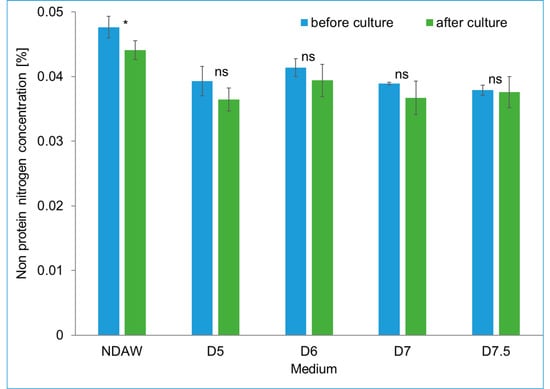
Figure 7.
The content of non-protein nitrogen in NDAW and deproteinized AW media during culture of K. xylinus K2G30. * significant difference in the medium component before and after culture, p-value 0.05. ns non-significant difference.
The total acidity and content of lactic acid in the NDAW and deproteinized AW media are presented in Table 3. For all media, an initial pH value of 5.0 was established. Subsequently, culturing was conducted, after which the change in the active acidity and titratable acidity of the media were assessed. The pH of the deproteinized AW media after culturing ranged from 5.79 ± 0.037 to 7.33 ± 0.421, compared with the pH of the of NDAW medium, amounting to 6.38 ± 0.065 (Table 3). These values indicate that the pH increased due to the conducted culturing of K. xylinus K2G30 and BC biosynthesis. During the cultivation of AAB in order to produce the BC, as a consequence of the conversion of glucose into gluconic acid, the pH of the medium decreased [30]. However, according to the data derived from the literature, the pH of the medium could increase during the production of BC in whey. In the pre-culture Revin et al. [45] study, the pH of whey was about 5.0 and increased to 8.0 after the end of culturing. The authors suggested that this is related to the consumption of the organic acids from the medium by the bacteria producing BC. This may be confirmed by the titratable acidity values of the media under study. The titratable acidity is expressed in °SH and 1 °SH corresponds to 22.5 mg of lactic acid. In the conducted studies, the concentration of lactic acid decreased during the culture in all of the analyzed media (Table 3). It should be noted that in the deproteinized media, the lactic acid content before cultivation was different and during the deproteinization process its level of concentration decreased while the pH level increased. Most probably, this was due the formation of the complexes of lactates with calcium, phosphates, and proteins; therefore, there were fewer unbound lactates [49]. The highest lactic acid content after culturing was observed for the medium D5 and the lowest one for the medium D7, with the values of 207 ± 3.674 mg·100 mL−1 and 40.5 ± 7.348 mg·100 mL−1, respectively. For all deproteinized whey media, the difference between the lactic acid contents before and after the cultivation of acetic bacteria was similar and ranged from 61.5 mg to 69 mg for medium D7 and D6, respectively. This may indicate a comparable ability to grow and use nutrients on the part of acetic bacteria in the deproteinized whey media. In the NDAW medium, the difference in lactic acid content before and after cultivation was 174 mg. The almost three times higher (compared with other media) use of lactic acid by K. xylinus K2G30 bacteria with the production of the largest BC mass at the same time could suggest that lactic acid had a positive impact on the increased efficiency of BC biosynthesis. The literature confirms that the addition of 0.4–1.25% lactic acid improves the efficiency of BC production. This is caused by the lactic acid acting as a promoter of the Krebs cycle, positively affecting the growth of the biomass of acetic bacteria and, consequently, the efficiency of the BC production [53,56].
Table 3.
Total acidity and lactic acid content in the NDAW medium and deproteinized AW media.
The calcium content in AW could be about 0.0699%, while the content of magnesium amounts to about 0.0068% [57]. This study determined the content of Ca2+ and Mg2+ ions in the media both before and after culturing, in order to assess the impact of these macroelements on BC production by the K. xylinus K2G30 strain. The calcium and magnesium contents in the NDAW and deproteinized whey media are presented in Figure 8.
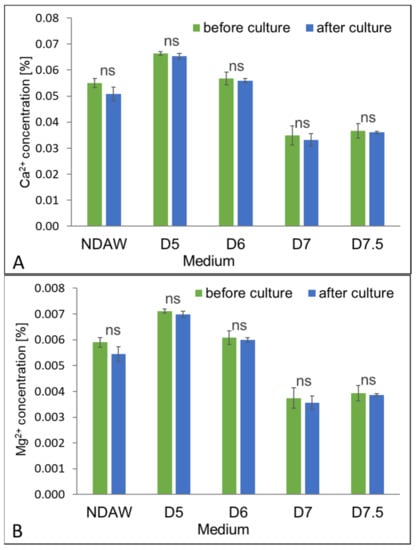
Figure 8.
The content of calcium (A) and magnesium (B) in NDAW medium and deproteinized AW media during culture of K. xylinus K2G30. ns non-significant difference.
The results indicate that the amount of calcium and magnesium in the media before cultivation depended on the conditions of whey deproteinization, and decreased as the pH rose during the process. The content of calcium ranged from 0.0349% to 0.0664% in media D7 and D5, respectively. The content of magnesium ranged from 0.0037% to 0.0071% in media D7 and D5, respectively. In NDAW, the content of Ca2+ and Mg2+ ions before cultivation amounted to 0.0551% and 0.0059%, respectively. After culturing, no significant changes in the calcium and magnesium content in any of the tested media were observed, which could indicate that these minerals were not used by bacterial cells in order to produce BC. This is interesting because, according to Mohite et al. [58], calcium and magnesium play an important role in the process of BC biosynthesis through participation in the synthesis and activation of c-di-GMP—one of the cellulose synthase activators—an enzyme responsible for the biosynthesis of biocellulose. Typically, in chemically composed media, the calcium and magnesium content ensure optimal cellulose yield at about 0.8% (w/v) and 0.04% (w/v), respectively [58]. In this study, however, the Ca2+ and Mg2+ content was definitely lower, which could have also negatively impacted cellulose production. The low consumption level of Ca2+ and Mg2+ by K. xylinus for BC biosynthesis could also be due to the low availability of these ions to bacterial cells. Probably, the relatively low proteolytic activity of AAB was insufficient to release the calcium and magnesium ions from whey protein complexes [41,59].
3.4. Bacterial Cellulose Biosynthesis in AW Media
Figure 9 shows the mass of BC produced by the K. xylinus K2G30 strain in the media of deproteinized AW and NDAW. As a result of the cultivation of the K. xylinus K2G30 strain in AW media, the highest mass of BC was obtained in NDAW, i.e., 1.57 g·dm−3. The effectiveness of BC biosynthesis in deproteinized and NDAW media was compared with the BC mass obtained in the classical YPM medium (Figure 1). As expected, the most BC (5.23 g·dm−3) was obtained in the YPM medium. However, it should be noted that the YPM medium is one of the media that are characterized by the best conditions for the growth of K. xylinus K2G30 bacteria in the literature [28]. This difference indicates that further research is required to increase the efficiency of BC production in AW. However, the results obtained in this study show that the de-proteinization of whey is not an appropriate direction for increasing the intensity of BC biosynthesis and deproteinization negatively affected the production of BC. The least BC was obtained in deproteinized medium D7 and D7.5 (0.63 g·dm−3 and 0.61 g·dm−3, respectively). This could be because some nutrients such as proteins, organic acids, or minerals could be precipitated and separated from the medium after centrifugation under the influence of a high temperature and pH.

Figure 9.
Dry mass of bacterial cellulose produced by K. xylinus K2G30 on NDAW and deproteinized AW media columns marked with the same letter belong to one homogeneous group and there are no significant differences between them. Columns marked with different letters belong to other homogeneous groups and there are statistically significant differences between them.
The wet and dry BC obtained in deproteinized AW media and NDAW media are shown in Figure 10.

Figure 10.
Wet and dry BC obtained in culture of K. xylinus K2G30 in NDAW and deproteinized AW media.
3.5. FTIR Characterization of Bacterial Cellulose
ATR-FTIR spectroscopy was carried out to compare the chemical structure of the BC variants and to verify the effect of de-proteinization conditions of AW as a culture medium on the BC structure. The FTIR spectra of the BC obtained in YPM classical medium were also presented for comparison. The FTIR spectra are shown in Figure 11.
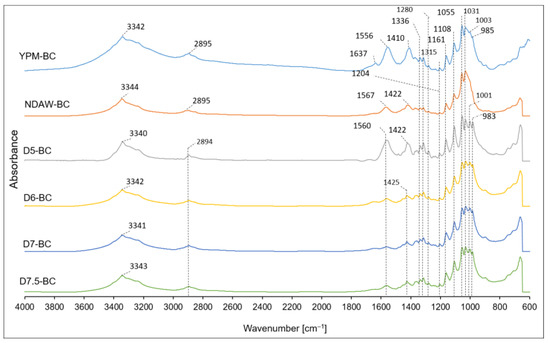
Figure 11.
ATR-FTIR spectra of the BC obtained from the culture of K. xylinus K2G30 in YPM, NDAW, and deproteinized AW media.
From the spectra, it was observed that all samples had bands characteristic of BC, with slight shifts relative to one another. There were no significant differences between the spectra of BC obtained in non-proteinized whey and deproteinized whey under different conditions. At a wavenumber of about 3340 cm−1, a band characteristic of O-H stretching vibrations was observed [60]. The peaks at 2894 cm−1 corresponded to C-H stretching in the CH2 and CH3 groups [61]. The peak presented around 1640 cm−1 was commonly attributed to the H-O-H bending vibrations of absorbed water [61,62]. Peaks around 1556 cm−1 and 1567 cm−1 were present in all BC samples, which may indicate that their presence did not depend on the medium. Peaks around 1560–1570 cm−1 were also observed by Wang et al. in studies where BC was obtained in media with different carbon sources. However, this peak was described as being unknown [63]. In this study, peaks near 1410 cm−1–1425 cm−1 and 1315 cm−1 were observed on the spectrum, which could correspond to CH2 symmetrical stretching and CH2 out-of-plane wagging, respectively, as well as confirming crystalline regions in the BC structure [61,63]. The peaks at 1335 cm−1 were attributed to C-H deformation or O-H in-plane bending. In addition, the presence of peaks at 1335 cm−1, 1315 cm−1, and 1280 cm−1 (C-H bending) could suggest that BC was composed, to some extent, of cellulose II. However, the peaks at 3340 cm−1, 1425 cm−1, 1160 cm−1, and 899 cm−1 indicated that BC was composed mainly of cellulose I [63,64]. Characteristic BC peaks at 1205 cm−1 and 1161 cm−1, attributed to symmetrical and asymmetrical stretching vibrations of C-O-C bonds, respectively, were also observed. Other BC-specific peaks were detected at 1108 cm−1, 1055 cm−1, and 1031 cm−1 corresponding to C-O-C 1,4-β-glycosidic linkages or skeletal vibrations, C-O-C pyranose ring skeletal vibrations, and C-O deformation, respectively [65]. Interestingly, Molina-Ramírez et al. [66] observed peaks in the spectra of BC obtained in whey that were characteristic of milk fat (2921 cm−1 and 2851 cm−1), esters and ethers (1743 cm−1 and 1163 cm−1, respectively), and amine and amide groups in proteins (about 3400 cm−1, 1540 cm−1, and 1577 cm−1), which were not found in this study. This may indicate the relatively high purity of BC obtained in AW, regardless of whether it was deproteinized or not.
4. Conclusions
The conducted research revealed that the K. xylinus K2G30 strain was characterized by the highest efficiency for BC production in both the classical and waste media. Based on the results, it can be concluded that the Ga. hansenii strain exhibited greater BC biosynthesis in the waste media of a plant origin (banana peels, celery pomace, hemp pomace) compared with the waste of an animal origin (fish waste and acid whey). On the contrary, the K. xylinus strain produced more BC in the medium of an animal origin (fish waste and acid whey) compared with the plant origin media. For both strains of Ga. hansenii and K. xylinus, a trend was noted that the type of waste was more important for BC biosynthesis than its concentration. The only differences between concentrations within the same waste were noted in banana peels and celery pomace media for both strains. However, AW with a concentration of 70 g·dm−3 was recognised as the best waste medium for BC obtained by K. xylinus. In truth, there were no significant differences between AW concentrations; from an economic point of view, choosing a concentration of 20 g·dm−3 would be suggested. However, the highest mass of BC was obtained at a concentration of 70 g·dm−3. In addition, it is worth noting that AW was reconstituted from powder and a concentration of 70 g·dm−3 was most similar to the unprocessed liquid AW sourced directly from industry. The use of such AW would be the most preferable because it would eliminate the AW powdering process, which could reduce the cost for obtaining BC.
In the second stage of this study, we attempted to modify the medium with AW through deproteinization. Importantly, AW deproteinization did not affect the structure of BC, which was confirmed by the analysis of the FTIR spectra. However, it was found that non-deproteinized AW was characterized by the most appropriate composition for BC biosynthesis compared with the deproteinized media. On such grounds, it may be concluded that the presence of whey proteins positively influenced the BC biosynthesis. These results also confirm the validity of using unprocessed AW for BC biosynthesis and reducing the cost of the process. Interestingly, in the case of the AW medium where the mass of BC was the highest, a considerable decrease in lactic acid content was noted. This creates opportunities to expand on research regarding BC biosynthesis in AW, with particular emphasis on the impact of lactic acid. Despite the above results, it should be remembered that the efficiency of BC biosynthesis in AW was definitely lower than in the classic YPM medium. Therefore, in order to improve the efficiency of BC production in AW, while keeping costs as low as possible, further research should be conducted.
Author Contributions
Conceptualization, J.P. and L.S.-R.; investigation, J.P., S.K. and L.S.-R.; methodology, J.P. and L.S.-R.; supervision, L.S.-R.; writing—original draft, J.P.; writing—review and editing, J.P., M.G. and L.S.-R. All authors have read and agreed to the published version of the manuscript.
Funding
Part of the research (FTIR analysis) was conducted during a Justyna Płoska scholarship funded by the Polish National Agency of Academic Exchange (NAWA) program for students and scientists as part of bilateral cooperation, No. BPN/BIL/2021/1/00171/U/00001.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lee, J.K.; Patel, S.K.S.; Sung, B.H.; Kalia, V.C. Biomolecules from municipal and food industry wastes: An overview. Bioresour. Technol. 2020, 298, 122346. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Z.; Sajjad, W.; Khan, T.; Wahid, F. Production of bacterial cellulose from industrial wastes: A review. Cellulose 2019, 26, 2895–2911. [Google Scholar] [CrossRef]
- Teuber, R.; Jensen, J.D. Definitions, measurement, and drivers of food loss and waste. In Food Industry Wastes; Kosseva, M.R., Webb, C., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 3–18. [Google Scholar]
- Neri, A.; Negri, M.; Cagno, E.; Franzò, S.; Kumar, V.; Lampertico, T.; Bassani, C.A. The role of digital technologies in supporting the implementation of circular economy practices by industrial small and medium enterprises. Bus. Strateg. Environ. 2023, 1–26. [Google Scholar] [CrossRef]
- Merino, D.; Quilez-Molina, A.I.; Perotto, G.; Bassani, A.; Spigno, G.; Athanassiou, A. A second life for fruit and vegetable waste: A review on bioplastic films and coatings for potential food protection applications. Green Chem. 2022, 24, 4703–4727. [Google Scholar] [CrossRef]
- Mohsin, A.; Zhang, K.; Hu, J.; Salim-ur-Rehman; Tariq, M.; Zaman, W.Q.; Khan, I.M.; Zhuang, Y.; Guo, M. Optimized biosynthesis of xanthan via effective valorization of orange peels using response surface methodology: A kinetic model approach. Carbohydr. Polym. 2018, 181, 793–800. [Google Scholar] [CrossRef]
- Mohsin, A.; Sun, J.; Khan, I.M.; Hang, H.; Tariq, M.; Tian, X.; Ahmed, W.; Niazi, S.; Zhuang, Y.; Chu, J.; et al. Sustainable biosynthesis of curdlan from orange waste by using Alcaligenes faecalis: A systematically modeled approach. Carbohydr. Polym. 2019, 205, 626–635. [Google Scholar] [CrossRef]
- Kassim, M.A.; Hussin, A.H.; Meng, T.K.; Kamaludin, R.; Zaki, M.S.I.M.; Zakaria, W.Z.E.W. Valorisation of watermelon (Citrullus Lanatus) rind waste into bioethanol: An optimization and kinetic studies. Int. J. Environ. Sci. Technol. 2021, 19, 2545–2558. [Google Scholar] [CrossRef]
- Panesar, P.S.; Kaur, S. Bioutilisation of agro-industrial waste for lactic acid production. Int. J. Food Sci. Technol. 2015, 50, 2143–2151. [Google Scholar] [CrossRef]
- Qiu, X.; Zhang, Y.; Hong, H. Classification of acetic acid bacteria and their acid resistant mechanism. AMB Express 2021, 11, 29. [Google Scholar] [CrossRef]
- Qin, Z.; Yu, S.; Chen, J.; Zhou, J. Dehydrogenases of acetic acid bacteria. Biotechnol. Adv. 2022, 54, 107863. [Google Scholar] [CrossRef]
- Gomes, R.J.; Borges, M.D.F.; Rosa, M.D.F.; Castro-Gómez, R.J.H.; Spinosa, W.A. Acetic acid bacteria in the food industry: Systematics, characteristics and applications. Food Technol. Biotechnol. 2018, 56, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Płoska, J.; Garbowska, M.; Pluta, A.; Stasiak-Różańska, L. Bacterial cellulose—Innovative biopolymer and possibilities of its applications in dairy industry. Int. Dairy J. 2023, 140, 105586. [Google Scholar] [CrossRef]
- Ahmed, J.; Gultekinoglu, M.; Edirisinghe, M. Bacterial cellulose micro-nano fibres for wound healing applications. Biotechnol. Adv. 2020, 41, 107549. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.U.; Ul-Islam, M.; Ahsan, H.; Ahmed, M.B.; Shehzad, A.; Fatima, A.; Sonn, J.K.; Lee, Y.S. Potential applications of bacterial cellulose and its composites for cancer treatment. Int. J. Biol. Macromol. 2021, 168, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Robbins, M.; Pisupati, V.; Azzarelli, R.; Nehme, S.I.; Barker, R.A.; Fruk, L.; Schierle, G.S.K. Biofunctionalised bacterial cellulose scaffold supports the patterning and expansion of human embryonic stem cell-derived dopaminergic progenitor Cells. Stem Cell Res. Ther. 2021, 12, 574. [Google Scholar] [CrossRef] [PubMed]
- Ludwicka, K.; Kolodziejczyk, M.; Gendaszewska-Darmach, E.; Chrzanowski, M.; Jedrzejczak-Krzepkowska, M.; Rytczak, P.; Bielecki, S. Stable composite of bacterial nanocellulose and perforated polypropylene mesh for biomedical applications. J. Biomed. Mater. Res. Part B Appl. Biomater. 2019, 107, 978–987. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Liu, Z.; Shen, R.; Chen, S.; Yang, X. Bacterial cellulose in food industry: Current research and future prospects. Int. J. Biol. Macromol. 2020, 158, 1007–1019. [Google Scholar] [CrossRef]
- Dirpan, A.; Djalal, M.; Kamaruddin, I. Application of an intelligent sensor and active packaging system based on the bacterial cellulose of Acetobacter xylinum to meat products. Sensors 2022, 22, 544. [Google Scholar] [CrossRef]
- Stasiak-Różańska, L.; Płoska, J. Study on the use of microbial cellulose as a biocarrier for 1,3-dihydroxy-2-propanone and its potential application in industry. Polymers 2018, 10, 438. [Google Scholar] [CrossRef]
- Martins, D.; Rocha, C.; Dourado, F.; Gama, M. Bacterial cellulose-carboxymethyl cellulose (BC:CMC) dry formulation as stabilizer and texturizing agent for surfactant-free cosmetic formulations. Colloids Surf. A Physicochem. Eng. Asp. 2021, 617, 126380. [Google Scholar] [CrossRef]
- Wang, F.-P.; Zhao, X.-J.; Wahid, F.; Zhao, X.-Q.; Qin, X.-T.; Bai, H.; Xie, Y.-Y.; Zhong, C.; Jia, S.-R. Sustainable, superhydrophobic membranes based on bacterial cellulose for gravity-driven oil/water separation. Carbohydr. Polym. 2021, 253, 117220. [Google Scholar] [CrossRef] [PubMed]
- Sperotto, G.; Stasiak, L.G.; Godoi, J.P.M.G.; Gabiatti, N.C.; De Souza, S.S. A review of culture media for bacterial cellulose production: Complex, chemically defined and minimal media modulations. Cellulose 2021, 28, 2649–2673. [Google Scholar] [CrossRef]
- Tomaszewska, M.; Białończyk, L. Ethanol production from whey in a bioreactor coupled with direct contact membrane distillation. Catal. Today 2016, 268, 156–163. [Google Scholar] [CrossRef]
- Rocha-Mendoza, D.; Kosmerl, E.; Krentz, A.; Zhang, L.; Badiger, S.; Miyagusuku-Cruzado, G.; Mayta-Apaza, A.; Giusti, M.; Jiménez-Flores, R.; García-Cano, I. Invited review: Acid whey trends and health benefits. J. Dairy Sci. 2021, 104, 1262–1275. [Google Scholar] [CrossRef]
- Zotta, T.; Solieri, L.; Iacumin, L.; Picozzi, C.; Gullo, M. Valorization of cheese whey using microbial fermentations. Appl. Microbiol. Biotechnol. 2020, 104, 2749–2764. [Google Scholar] [CrossRef]
- Chatterjee, S.; Guha, A.K. A Study on biochemical changes during cultivation of Rhizopus oryzae in deproteinized whey medium in relation to chitosan production. Lett. Appl. Microbiol. 2014, 59, 155–160. [Google Scholar] [CrossRef]
- Gullo, M.; La China, S.; Petroni, G.; Di Gregorio, S.; Giudici, P. Exploring K2G30 Genome: A high bacterial cellulose producing strain in glucose and mannitol based media. Front. Microbiol. 2019, 10, 58. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Schäfer, J.; Sebald, K.; Dunkel, A.; Hofmann, T.; Rosenthal, I.; Schuster, R.; Atamer, Z.; Hinrichs, J. A Feasibility study on the pilot scale manufacture of fresh cheese from skim milk retentates without acid whey production: Effect of calcium content on bitterness and texture. Int. Dairy J. 2019, 93, 72–80. [Google Scholar] [CrossRef]
- PN-EN ISO 8968-4; Milk and Milk Products—Determination of Nitrogen Content—Part 4: Determination of Protein and Non Protein Nitrogen Content and True Protein Content Calculation. ISO: Geneva, Switzerland, 2016.
- Aiello, A.; Pizzolongo, F.; Manzo, N.; Romano, R. A new method to distinguish the milk adulteration with neutralizers by detection of lactic acid. Food Anal. Methods 2019, 12, 2555–2561. [Google Scholar] [CrossRef]
- Lin, S.P.; Loira Calvar, I.; Catchmark, J.M.; Liu, J.R.; Demirci, A.; Cheng, K.C. Biosynthesis, production and applications of bacterial cellulose. Cellulose 2013, 20, 2191–2219. [Google Scholar] [CrossRef]
- Zhong, C.; Zhang, G.C.; Liu, M.; Zheng, X.T.; Han, P.P.; Jia, S.R. Metabolic flux analysis of Gluconacetobacter xylinus for bacterial cellulose production. Appl. Microbiol. Biotechnol. 2013, 97, 6189–6199. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Liu, L.; Jia, S.; Li, S.; Zou, Y.; Zhong, C. Complete genome analysis of Gluconacetobacter xylinus CGMCC 2955 for elucidating bacterial cellulose biosynthesis and metabolic regulation. Sci. Rep. 2018, 8, 6266. [Google Scholar] [CrossRef] [PubMed]
- Bozdağ, G.; Pinar, O.; Gündüz, O.; Kazan, D. Valorization of pea pod, celery root peel, and mixed-vegetable peel as a feedstock for biocellulose production from Komagataeibacter hansenii DSM 5602. Biomass Convers. Biorefinery 2021, 1, 3. [Google Scholar] [CrossRef]
- Moukamnerd, C.; Ounmuang, K.; Konboa, N.; Insomphun, C. Bacterial cellulose production by Komagataeibacter nataicola TISTR 2661 by agro-waste as a carbon source. Chiang Mai J. Sci. 2020, 47, 16–27. [Google Scholar]
- Pommet, M.; Juntaro, J.; Heng, J.Y.Y.; Mantalaris, A.; Lee, A.F.; Wilson, K.; Kalinka, G.; Shaffer, M.S.P.; Bismarck, A. Surface modification of natural fibers using bacteria: Depositing bacterial cellulose onto natural fibers to create hierarchical fiber reinforced nanocomposites. Biomacromolecules 2008, 9, 1643–1651. [Google Scholar] [CrossRef]
- Vidya, D.; Nayana, K.; Sreelakshmi, M.; Keerthi, K.V.; Mohan, K.S.; Sudhakar, M.P.; Arunkumar, K. A Sustainable cultivation of microalgae using dairy and fish wastes for enhanced biomass and bio-product production. Biomass Convers. Biorefinery 2021, 1–15. [Google Scholar] [CrossRef]
- Gao, M.T.; Hirata, M.; Toorisaka, E.; Hano, T. Acid-hydrolysis of fish wastes for lactic acid fermentation. Bioresour. Technol. 2006, 97, 2414–2420. [Google Scholar] [CrossRef]
- Kolesovs, S.; Semjonovs, P. Production of bacterial cellulose from whey—Current state and prospects. Appl. Microbiol. Biotechnol. 2020, 104, 7723–7730. [Google Scholar] [CrossRef]
- Pastorkova, E.; Zakova, T.; Landa, P.; Novakova, J.; Vadlejch, J.; Kokoska, L. Growth inhibitory effect of grape phenolics against wine spoilage yeasts and acetic acid bacteria. Int. J. Food Microbiol. 2013, 161, 209–213. [Google Scholar] [CrossRef]
- Yao, Y.; Ren, G. Effect of thermal treatment on phenolic composition and antioxidant activities of two celery cultivars. LWT Food Sci. Technol. 2011, 44, 181–185. [Google Scholar] [CrossRef]
- Coppola, D.; Lauritano, C.; Palma Esposito, F.; Riccio, G.; Rizzo, C.; de Pascale, D. Fish waste: From problem to valuable resource. Mar. Drugs 2021, 19, 116. [Google Scholar] [CrossRef] [PubMed]
- Revin, V.; Liyaskina, E.; Nazarkina, M.; Bogatyreva, A.; Shchankin, M. Cost-effective production of bacterial cellulose using acidic food Industry by-products. Braz. J. Microbiol. 2018, 49, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Carreira, P.; Mendes, J.A.S.; Trovatti, E.; Serafim, L.S.; Freire, C.S.R.; Silvestre, A.J.D.; Neto, C.P. Utilization of residues from agro-forest industries in the production of high value bacterial cellulose. Bioresour. Technol. 2011, 102, 7354–7360. [Google Scholar] [CrossRef]
- Salari, M.; Sowti Khiabani, M.; Rezaei Mokarram, R.; Ghanbarzadeh, B.; Samadi Kafil, H. Preparation and characterization of cellulose nanocrystals from bacterial cellulose produced in sugar beet molasses and cheese whey media. Int. J. Biol. Macromol. 2019, 122, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Sharma, D.K.; Sandhu, P.P.; Jadaun, J.; Sangwan, R.S.; Yadav, S.K. Sustainable process for the production of cellulose by an Acetobacter pasteurianus RSV-4 (MTCC 25117) on whey medium. Cellulose 2021, 28, 103–116. [Google Scholar] [CrossRef]
- Chandrapala, J.; Duke, M.C.; Gray, S.R.; Zisu, B.; Weeks, M.; Palmer, M.; Vasiljevic, T. Properties of acid whey as a function of pH and temperature. J. Dairy Sci. 2015, 98, 4352–4363. [Google Scholar] [CrossRef]
- Lappa, I.K.; Kachrimanidou, V.; Papadaki, A.; Stamatiou, A.; Ladakis, D.; Eriotou, E.; Kopsahelis, N. A Comprehensive bioprocessing approach to foster cheese whey valorization: On-site β-galactosidase secretion for lactose hydrolysis and sequential bacterial cellulose production. Fermentation 2021, 7, 184. [Google Scholar] [CrossRef]
- Tsermoula, P.; Khakimov, B.; Nielsen, J.H.; Engelsen, S.B. Whey–The waste-stream that became more valuable than the food product. Trends Food Sci. Technol. 2021, 118, 230–241. [Google Scholar] [CrossRef]
- Gomes, R.J.; Ida, E.I.; Spinosa, W.A. Nutritional supplementation with amino acids on bacterial cellulose production by Komagataeibacter intermedius: Effect analysis and application of response surface methodology. Appl. Biochem. Biotechnol. 2022, 194, 5017–5036. [Google Scholar] [CrossRef]
- Matsuoka, M.; Tsuchida, T.; Matsushita, K.; Adachi, O.; Yoshinaga, F. A Synthetic Medium for bacterial cellulose production by Acetobacter xylinum subsp. sucrofermentans. Biosci. Biotechnol. Biochem. 1996, 60, 575–579. [Google Scholar] [CrossRef]
- Heo, M.-S.; Son, H.-J. Development of an optimized, simple chemically defined medium for bacterial cellulose production by Acetobacter Sp. A9 in shaking cultures. Biotechnol. Appl. Biochem. 2002, 36, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Magan, J.B.; O’callaghan, T.F.; Zheng, J.; Zhang, L.; Mandal, R.; Hennessy, D.; Fenelon, M.A.; Wishart, D.S.; Kelly, A.L.; McCarthy, N.A. Impact of bovine diet on metabolomic profile of skim milk and whey protein ingredients. Metabolites 2019, 9, 305. [Google Scholar] [CrossRef] [PubMed]
- Naritomi, T.; Kouda, T.; Yano, H.; Yoshinaga, F. Effect of lactate on bacterial cellulose production from fructose in continuous culture. J. Ferment. Bioeng. 1998, 85, 89–95. [Google Scholar] [CrossRef]
- Menchik, P.; Zuber, T.; Zuber, A.; Moraru, C.I. Short communication: Composition of coproduct streams from dairy processing: Acid whey and milk permeate. J. Dairy Sci. 2019, 102, 3978–3984. [Google Scholar] [CrossRef] [PubMed]
- Mohite, B.V.; Salunke, B.K.; Patil, S.V. Enhanced production of bacterial cellulose by using Gluconacetobacter hansenii NCIM 2529 strain under shaking conditions. Appl. Biochem. Biotechnol. 2013, 169, 1497–1511. [Google Scholar] [CrossRef]
- Volova, T.G.; Prudnikova, S.V.; Sukovatyi, A.G.; Shishatskaya, E.I. Production and properties of bacterial cellulose by the strain Komagataeibacter xylinus B-12068. Appl. Microbiol. Biotechnol. 2018, 102, 7417–7428. [Google Scholar] [CrossRef]
- Molina-Ramírez, C.; Enciso, C.; Torres-Taborda, M.; Zuluaga, R.; Gañán, P.; Rojas, O.J.; Castro, C. Effects of alternative energy sources on bacterial cellulose characteristics produced by Komagataeibacter medellinensis. Int. J. Biol. Macromol. 2018, 117, 735–741. [Google Scholar] [CrossRef]
- Cielecka, I.; Ryngajłło, M.; Maniukiewicz, W.; Bielecki, S. Response surface methodology-based improvement of the yield and differentiation of properties of bacterial cellulose by metabolic enhancers. Int. J. Biol. Macromol. 2021, 187, 584–593. [Google Scholar] [CrossRef]
- Lotfy, V.F.; Basta, A.H.; Abdel-Monem, M.O.; Abdel-Hamed, G.Z. Utilization of bacteria in rotten guava for production of bacterial cellulose from isolated and protein waste. Carbohydr. Polym. Technol. Appl. 2021, 2, 100076. [Google Scholar] [CrossRef]
- Wang, S.S.; Han, Y.H.; Chen, J.L.; Zhang, D.C.; Shi, X.X.; Ye, Y.X.; Chen, D.L.; Li, M. Insights into bacterial cellulose biosynthesis from different carbon sources and the associated biochemical transformation pathways in Komagataeibacter sp. W1. Polymers 2018, 10, 963. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Han, Y.H.; Ye, Y.X.; Shi, X.X.; Xiang, P.; Chen, D.L.; Li, M. Physicochemical characterization of high-quality bacterial cellulose produced by Komagataeibacter sp. strain W1 and identification of the associated genes in bacterial cellulose production. RSC Adv. 2017, 7, 45145–45155. [Google Scholar] [CrossRef]
- Neelima, S.; Sreejith, S.; Shajahan, S.; Raj, A.; Vidya, L.; Aparna, V.M.; Radhakrishnan, E.K.; Sudarsanakumar, C. Highly Crystalline bacterial cellulose production by Novacetimonas hansenii strain isolated from rotten fruit. Mater. Lett. 2023, 333, 133622. [Google Scholar] [CrossRef]
- Molina-Ramírez, C.; Castro, C.; Zuluaga, R.; Gañán, P. Physical characterization of bacterial cellulose produced by Komagataeibacter medellinensis using food supply chain waste and agricultural by-products as alternative low-cost feedstocks. J. Polym. Environ. 2018, 26, 830–837. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).