
Black Fungi on Stone-Built Heritage: Current Knowledge and Future Outlook




Abstract
Featured Application
Abstract
1. Background
2. Black Fungi and Stone Monuments: An Intimate Connection
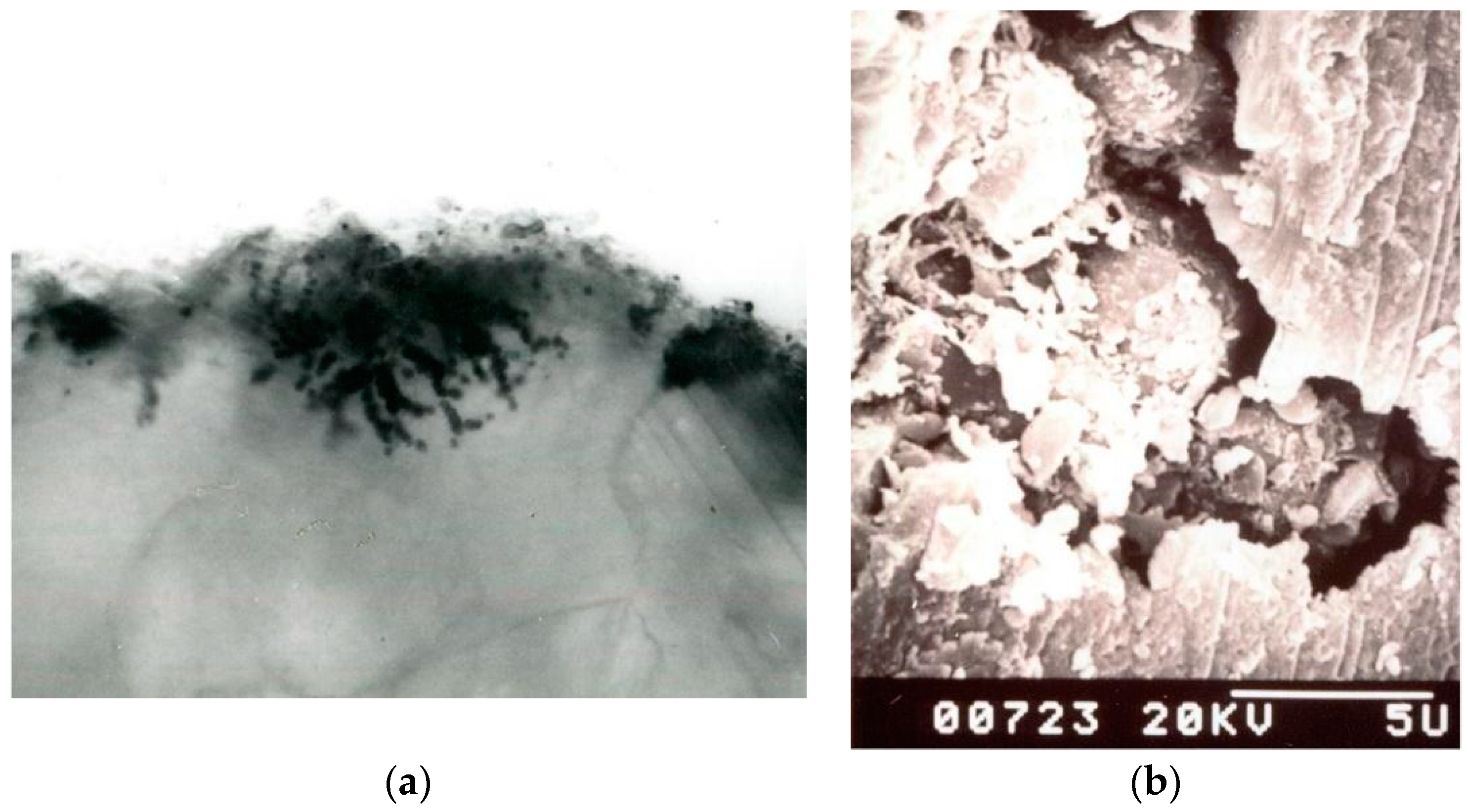
3. Mechanisms Involved in the Stone Biodeterioration
4. Multistep Analyses to Study Black Fungi from Stone Monuments
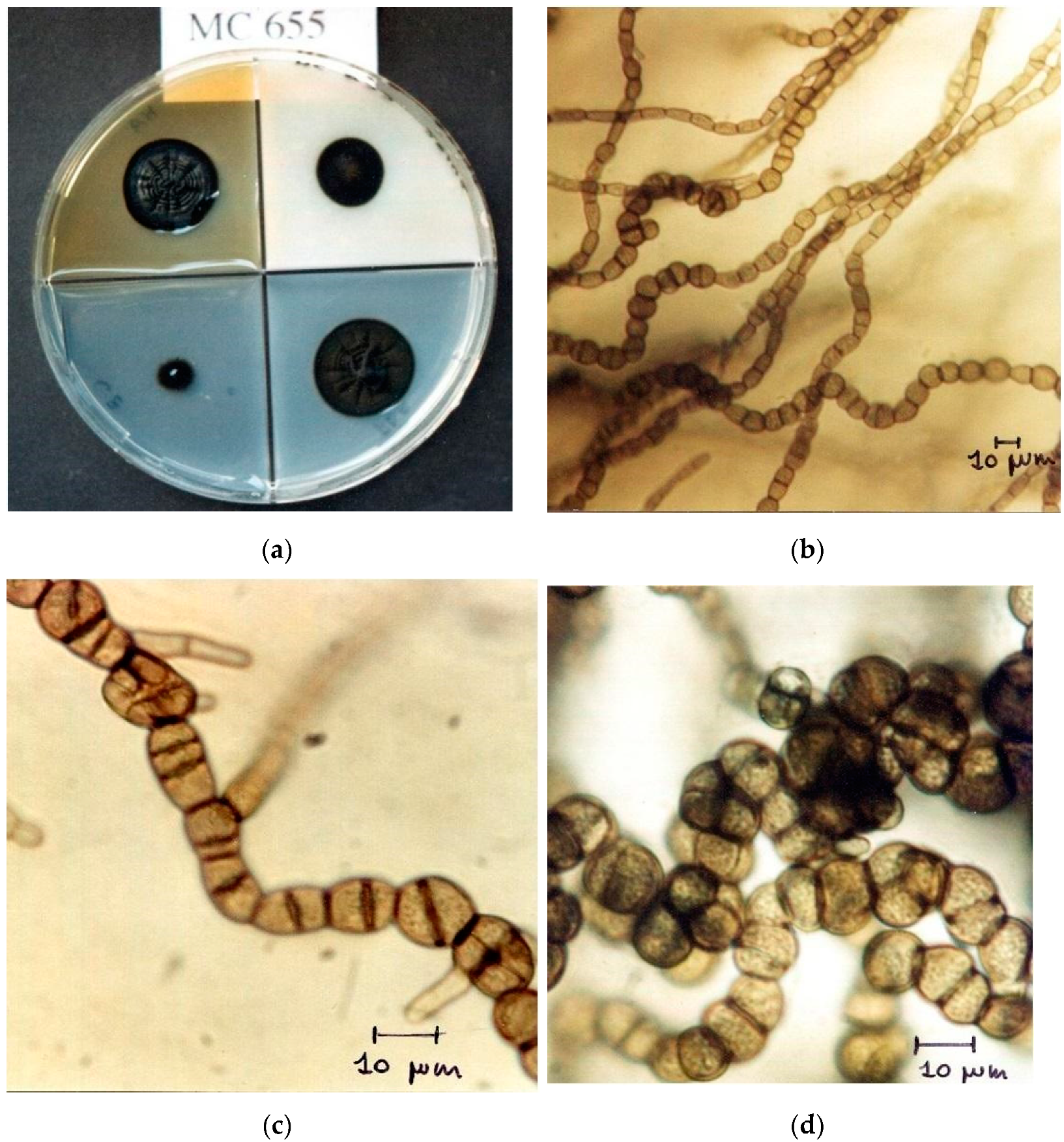
4.1. Methods of Isolation and Characterization
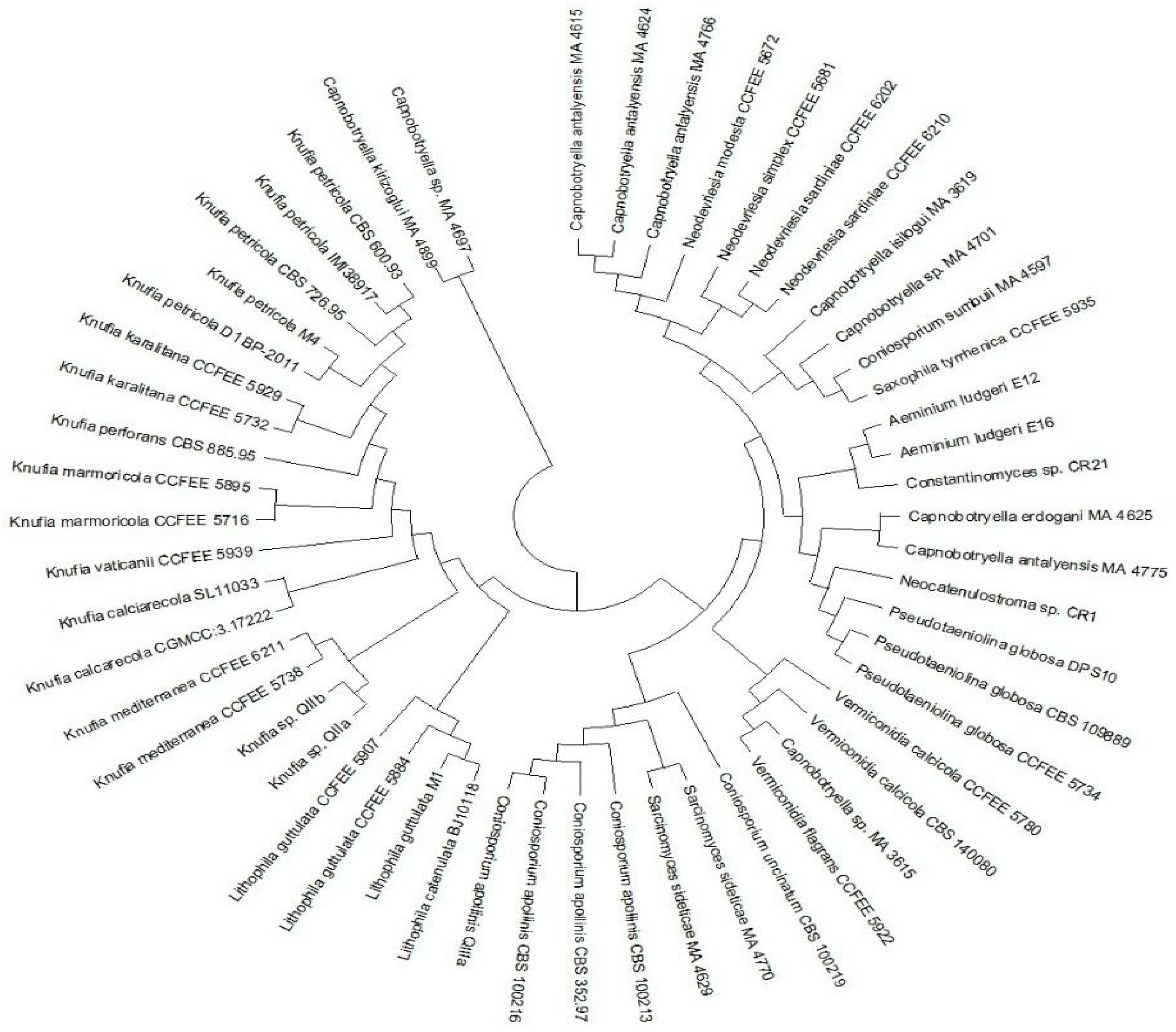
4.2. Culture Independent Analyses
5. How to Control Black Fungi
6. Concluding Remarks
- (1)
- Listen the conservators;
- (2)
- Evaluate the environmental climatic conditions and specific conditions, such as the type of material and the overall status of conservation of monument; the description of the type of alteration visible under naked eye should be also included;
- (3)
- Interact with the other experts involved;
- (4)
- Answer the questions posed by the conservators.
- (a)
- They are responsible of discolouring of the stone surface. The extended colonization of surfaces changes the global vision of the artifact, especially if different material and colour of stones were used by the artist;
- (b)
- Moreover, black fungi show an inter-crystalline pattern of growth. This pattern causes crystals to detach (so called sugaring) with loss of precious material, especially because it involves the first surface layer (very important for bas-reliefs and sculptures);
- (c)
- They could determine the biopitting. Fungi excavate cavities on the stone where they can better settle, giving the surface a pockmarked aspect. The convergence of several biopitting can often lead to larger cavities;
- (d)
- Hyphae penetrate deep into the surface, even more than a few mm;
- (e)
- Chemical and physical treatments used for other microorganisms are often non efficient in eradication;
- (f)
- Black fungi are often the first colonizers after the treatments.
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krumbein, W.E. Microbial interactions with mineral materials. In Biodeterioration 7, 1st ed.; Hougthon, D.R., Smith, R.N., Eggins, H.O., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 78–100. [Google Scholar] [CrossRef]
- Saiz-Jimenez, C. Biogeochemistry of weathering processes in monuments. Geomicrobiol. J. 1999, 16, 27–37. [Google Scholar] [CrossRef]
- Sterflinger, K. Fungi as geologic agents. Geomicrobiol. J. 2000, 17, 97–124. [Google Scholar] [CrossRef]
- Gadd, G.M. Geomicrobiology of the built environment. Nat. Microbiol. 2017, 2, 16275. [Google Scholar] [CrossRef] [PubMed]
- Urzì, C. Biodeterioramento dei manufatti artistici. In Microbiologia Ambientale ed Elementi di Ecologia Microbica; Barbieri, P., Bestetti, G., Galli, E., Zannoni, D., Eds.; Casa Editrice Ambrosiana: Milano, Italy, 2008; pp. 327–346. ISBN 978-88-08-18434-4. [Google Scholar]
- Pinna, D. Microbial growth and its effect on inorganic heritage material. In Microorganisms in the Deterioration and Preservation of Cultural Heritage, 1st ed.; Joseph, E., Ed.; Springer: Cham, Switzerland, 2021; pp. 3–35. [Google Scholar] [CrossRef]
- Gorbushina, A.A. Life on the rocks. Environ. Microbiol. 2007, 9, 1613–1631. [Google Scholar] [CrossRef] [PubMed]
- Villa, F.; Stewart, P.S.; Klapper, I.; Jacob, J.M.; Cappitelli, F. Subaerial biofilms on outdoor stone monuments: Changing the perspective toward an ecological framework. BioScience 2016, 66, 285–294. [Google Scholar] [CrossRef]
- Pinna, D. Coping with Biological Growth on Stone Heritage Objects: Methods, Products, Applications, and Perspectives, 1st ed.; Apple Academic Press: Oakville, ON, Canada, 2017; ISBN 9781771885324. [Google Scholar]
- Guillitte, O. Bioreceptivity: A new concept for building ecology studies. Sci. Total Environ. 1995, 167, 215–222. [Google Scholar] [CrossRef]
- Saiz-Jimenez, C. Deposition of anthropogenic compounds on monuments and their effect on airborne microorganisms. Aerobiologia 1995, 11, 161–175. [Google Scholar] [CrossRef]
- Urzì, C.; De Leo, F.; Salamone, P.; Criseo, G. Airborne fungal spores colonising marbles exposed in the terrace of Messina Museum, Sicily. Aerobiologia 2001, 17, 11–17. [Google Scholar] [CrossRef]
- Polo, A.; Gulotta, D.; Santo, N.; Di Benedetto, C.; Fascio, U.; Toniolo, L.; Villa, F.; Cappitelli, F. Importance of subaerial biofilms and airborne microflora in the deterioration of stonework: A molecular study. Biofouling 2012, 28, 1093–1106. [Google Scholar] [CrossRef][Green Version]
- Liu, B.; Fu, R.; Wu, B.; Liu, X.; Xiang, M. Rock-inhabiting fungi: Terminology, diversity, evolution and adaptation mechanisms. Mycology 2022, 13, 1–31. [Google Scholar] [CrossRef]
- Selbmann, L.; Egidi, E.; Isola, D.; Onofri, S.; Zucconi, Z.; de Hoog, G.S.; Chinaglia, S.; Testa, L.; Tosi, S.; Balestrazzi, A.; et al. Biodiversity, evolution and adaptation of fungi in extreme environments. Plant Biosyst. 2013, 147, 237–246. [Google Scholar] [CrossRef]
- Salvadori, O.; Municchia, A.C. The role of fungi and lichens in the biodeterioration of stone monuments. Open Conf. Proc. J. 2016, 7, 39–54. [Google Scholar] [CrossRef]
- Liu, X.; Koestler, R.J.; Warscheid, T.; Katayama, Y.; Gu, J.-D. Microbial deterioration and sustainable conservation of stone monuments and buildings. Nat. Sustain. 2020, 3, 991–1004. [Google Scholar] [CrossRef]
- Sterflinger, K. Fungi: Their role in deterioration of cultural heritage. Fungal Biol. Rev. 2010, 24, 47–55. [Google Scholar] [CrossRef]
- Urzì, C.; De Leo, F.; de Hoog, S.; Sterflinger, K. Recent advances in the molecular biology and ecophysiology of meristematic stone-inhabiting fungi. In Of Microbes and Art: The Role of Microbial Communities in the Degradation and Protection of Cultural Heritage; Ciferri, O., Tiano, P., Mastromei, G., Eds.; Springer: Boston, MA, USA, 2000; pp. 3–19. [Google Scholar] [CrossRef]
- Onofri, S.; Zucconi, L.; Isola, D.; Selbmann, L. Rock-inhabiting fungi and their role in deterioration of stone monuments in the Mediterranean area. Plant Biosyst. 2014, 148, 384–391. [Google Scholar] [CrossRef]
- Crous, P.W.; Schumacher, R.K.; Akulov, A.; Thangavel, R.; Hernández-Restrepo, M.; Carnegie, A.J.; Cheewangkoon, R.; Wingfield, M.J.; Summerell, B.A.; Quaedvlieg, W.; et al. New and interesting fungi. Fungal Syst. Evol. 2019, 3, 57–134. [Google Scholar] [CrossRef]
- Wollenzien, U.; de Hoog, G.S.; Krumbein, W.E.; Urzì, C. On the isolation of microcolonial fungi occurring on and in marble and other calcareous rocks. Sci. Total Environ. 1995, 167, 287–294. [Google Scholar] [CrossRef]
- Diakumaku, E.; Gorbushina, A.A.; Krumbein, W.E.; Panina, L.; Soukharjevski, S. Black fungi in marble and limestones–an aesthetical, chemical and physical problem for the conservation of monuments. Sci. Total Environ. 1995, 167, 295–304. [Google Scholar] [CrossRef]
- Urzì, C.; Wollenzien, U.; Criseo, G.; Krumbein, W.E. Biodiversity of the rock inhabiting microflora with special reference to black fungi and black yeasts. In Microbial Diversity and Ecosystem Function; Allsopp, D., Colwell, R.R., Hawksworth, D.L., Eds.; CAB International: Wallingford, UK, 1995; pp. 289–302. [Google Scholar]
- Sterflinger, K.; Krumbein, W.E. Dematiaceous fungi as a major agent for biopitting on mediterranean marbles and limestones. Geomicrobiol. J. 1997, 14, 219–230. [Google Scholar] [CrossRef]
- De Leo, F.; Urzì, C. Microfungi from deteriorated materials of cultural heritage. In Fungi from Different Substrates; Misra, J.K., Tewari, J.P., Deshmukh, S.K., Vágvölgyi, C., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 144–158. [Google Scholar] [CrossRef]
- Isola, D.; Zucconi, L.; Cecchini, A.; Caneva, G. Dark-pigmented biodeteriogenic fungi in etruscan hypogeal tombs: New data on their culture-dependent diversity, favouring conditions, and resistance to biocidal treatments. Fungal Biol. 2021, 125, 609–620. [Google Scholar] [CrossRef]
- de Hoog, G.S.; Guarro, J.; Gené, S.A.; Al-Hatmi, A.M.S.; Figueras, M.J.; Vitale, R.G. Atlas of Clinical Fungi, 4th ed.; Westerdijk Institute/Universitat Rovira Virgili: Utrecht, The Netherlands, 2019. [Google Scholar]
- Palmer, F.E.; Emery, D.R.; Stemmler, J.; Staley, J.T. Survival and growth of microcolonial rock fungi as affected by temperature and humidity. New Phytol. 1987, 107, 155–162. [Google Scholar] [CrossRef]
- Gueidan, C.; Ruibal, C.; de Hoog, S.; Schneider, H. Rock-inhabiting fungi originated during periods of dry climate in the late Devonian and middle Triassic. Fungal Biol. 2011, 115, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Gostinčar, C.; Grube, M.; Gunde-Cimerman, N. Evolution of fungal pathogens in domestic environments? Fungal Biol. 2011, 115, 1008–1018. [Google Scholar] [CrossRef] [PubMed]
- Gorbushina, A.A.; Whitehead, K.; Dornieden, T.; Niesse, A.; Schulte, A.; Hedges, J.I. Black fungal colonies as units of survival: Hyphal mycosporines synthesized by rock-dwelling microcolonial fungi. Can. J. Bot. 2003, 81, 131–138. [Google Scholar] [CrossRef]
- Gostinčar, C.; Muggia, L.; Grube, M. Polyextremotolerant black fungi: Oligotrophism, adaptive potential and a link to lichen symbioses. Front. Microbiol. 2012, 3, 390. [Google Scholar] [CrossRef] [PubMed]
- Zakharova, K.; Tesei, D.; Marzban, G.; Dijksterhuis, J.; Wyatt, T.; Sterflinger, K. Microcolonial fungi on rocks: A life in constant drought? Mycopathologia 2013, 175, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Marchetta, A.; van den Ende, G.B.; Al-Hatmi, A.M.S.; Hagen, F.; Zalar, P.; Sudhadham, M.; Gunde-Cimerman, N.; Urzì, C.; de Hoog, S.; De Leo, F. Global molecular diversity of the halotolerant fungus Hortaea werneckii. Life 2018, 8, 31. [Google Scholar] [CrossRef]
- Sterflinger, K.; Piñar, G. Microbial deterioration of cultural heritage and works of art—tilting at windmills? Appl. Microbiol. Biot. 2013, 97, 9637–9646. [Google Scholar] [CrossRef]
- Isola, D.; Zucconi, L.; Onofri, S.; Caneva, G.; de Hoog, G.S.; Selbmann, L. Extremotolerant rock inhabiting black fungi from Italian monumental sites. Fungal Divers. 2016, 76, 75–96. [Google Scholar] [CrossRef]
- Staley, J.T.; Palmer, F.; Adams, J.B. Microcolonial fungi: Common inhabitants on desert rocks? Science 1982, 215, 1093–1095. [Google Scholar] [CrossRef]
- De Leo, F.; Antonelli, F.; Pietrini, A.M.; Ricci, S.; Urzì, C. Study of the euendolithic activity of black meristematic fungi isolated from a marble statue in the Quirinale Palace’s Gardens in Rome, Italy. Facies 2019, 65, 18. [Google Scholar] [CrossRef]
- Santo, A.P.; Cuzman, O.A.; Petrocchi, D.; Pinna, D.; Salvatici, T.; Perito, B. Black on white: Microbial growth darkens the external marble of Florence cathedral. Appl. Sci. 2021, 11, 6163. [Google Scholar] [CrossRef]
- Marvasi, M.; Donnarumma, F.; Frandi, A.; Mastromei, G.; Sterflinger, K.; Tiano, P.; Perito, B. Black microcolonial fungi as deteriogens of two famous marble statues in Florence, Italy. Int. Biodeterior. Biodegrad. 2012, 68, 36–44. [Google Scholar] [CrossRef]
- Abdollahzadeh, J.; Groenewald, J.Z.; Coetzee, M.P.A.; Wingfield, M.J.; Crous, P.W. Evolution of lifestyles in Capnodiales. Stud. Mycol. 2020, 95, 381–414. [Google Scholar] [CrossRef]
- Sterflinger, K.; de Baere, R.; de Hoog, G.S.; de Wachter, R.; Krumbein, W.E.; Haase, G. Coniosporium perforans and C. apollinis, two new rock-inhabiting fungi isolated from marble in the Sanctuary of Delos (Cyclades, Greece). Antonie Van Leeuwenhoek J. Microb. 1997, 72, 349–363. [Google Scholar] [CrossRef]
- De Leo, F.; Urzì, C.; de Hoog, G.S. Two Coniosporium species from rock surfaces. Stud. Mycol. 1999, 43, 70–79. [Google Scholar]
- Sterflinger, K.; Prillinger, H. Molecular taxonomy and biodiversity of rock fungal communities in an urban environment (Vienna, Austria). Antonie Van Leeuwenhoek J. Microb. 2001, 80, 275–286. [Google Scholar] [CrossRef]
- Ricca, M.; Urzì, C.E.; Rovella, N.; Sardella, A.; Bonazza, A.; Ruffolo, S.A.; De Leo, F.; Randazzo, L.; Arcudi, A.; La Russa, M.F. Multidisciplinary approach to characterize archaeological materials and status of conservation of the roman Thermae of Reggio Calabria site (Calabria, south Italy). Appl. Sci. 2020, 10, 5106. [Google Scholar] [CrossRef]
- Sert, H.B.; Sterflinger, K. A new Coniosporium species from historical marble monuments. Mycol. Prog. 2010, 9, 353–359. [Google Scholar] [CrossRef]
- Sert, H.B.; Sümbül, H.; Sterflinger, K. Microcolonial fungi from antique marbles in Perge/Side/Termessos (Antalya/Turkey). Antonie Van Leeuwenhoek J. Microb. 2007, 91, 217–227. [Google Scholar] [CrossRef]
- Sazanova, K.V.; Zelenskaya, M.S.; Vlasov, A.D.; Bobir, S.Y.; Yakkonen, K.L.; Vlasov, D.Y. Microorganisms in superficial deposits on the stone monuments in Saint Petersburg. Microorganisms 2022, 10, 316. [Google Scholar] [CrossRef]
- Sert, H.B.; Sümbül, H.; Sterflinger, K. A new species of Capnobotryella from monument surfaces. Mycol. Res. 2007, 111, 1235–1241. [Google Scholar] [CrossRef] [PubMed]
- Sert, H.B.; Sümbül, H.; Sterflinger, K. Two new species of Capnobotryella from historical monuments. Mycol. Prog. 2011, 10, 333–339. [Google Scholar] [CrossRef]
- Sert, H.B.; Wuczkowski, M.; Sterflinger, K. Capnobotryella isiloglui, a new rock-inhabiting fungus from Austria. Turk. J. Bot. 2012, 3, 401–407. [Google Scholar] [CrossRef]
- Owczarek-Kościelniak, M.; Krzewicka, B.; Piątek, J.; Kołodziejczyk, Ł.M.; Kapusta, P. Is there a link between the biological colonization of the gravestone and its deterioration? Int. Biodeterior. Biodegrad. 2020, 148, 104879. [Google Scholar] [CrossRef]
- De Leo, F.; Urzì, C.; de Hoog, G.S. A new meristematic fungus, Pseudotaeniolina globosa. Antonie Van Leeuwenhoek J. Microb. 2003, 83, 351–360. [Google Scholar] [CrossRef]
- Egidi, E.; de Hoog, G.S.; Isola, D.; Onofri, S.; Quaedvlieg, W.; de Vries, M.; Verkley, G.J.M.; Stielow, J.B.; Zucconi, L.; Selbmann, L. Phylogeny and taxonomy of meristematic rock-inhabiting black fungi in the Dothideomycetes based on multi-locus phylogenies. Fungal Diver. 2014, 65, 127–165. [Google Scholar] [CrossRef]
- Rizk, S.M.; Magdy, M.; De Leo, F.; Werner, O.; Rashed, M.A.-S.; Ros, R.M.; Urzì, C. A new extremotolerant ecotype of the fungus Pseudotaeniolina globosa isolated from Djoser Pyramid, Memphis Necropolis, Egypt. J. Fungi 2021, 7, 104. [Google Scholar] [CrossRef]
- Trovão, J.; Tiago, I.; Soares, F.; Paiva, D.S.; Mesquita, N.; Coelho, C.; Catarino, L.; Gil, F.; Portugal, A. Description of Aeminiaceae fam. nov., Aeminium gen. nov. and Aeminium ludgeri sp. nov. (Capnodiales), isolated from a biodeteriorated art-piece in the old cathedral of Coimbra, Portugal. MycoKeys 2019, 45, 57–73. [Google Scholar] [CrossRef]
- Nuhoglu, Y.; Oguz, E.; Uslu, H.; Ozbek, A.; Ipekoglu, B.; Ocak, I.; Hasenekoglu, I. The accelerating effects of the microorganisms on biodeterioration of stone monuments under air pollution and continental-cold climatic conditions in Erzurum, Turkey. Sci. Total Environ. 2006, 364, 272–283. [Google Scholar] [CrossRef]
- Cappitelli, F.; Principi, P.; Pedrazzani, R.; Toniolo, L.; Sorlini, C. Bacterial and fungal deterioration of the Milan cathedral marble treated with protective synthetic resins. Sci. Total Environ. 2007, 385, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Cappitelli, F.; Nosanchuk, J.; Casadevall, A.; Toniolo, L.; Brusetti, L.; Florio, S.; Principi, P.; Borin, S.; Sorlini, C. Synthetic consolidants attacked by melanin-producing fungi: Case study of the biodeterioration of Milan (Italy) cathedral marble treated with acrylics. Appl. Environ. Microbiol. 2007, 73, 271–277. [Google Scholar] [CrossRef] [PubMed]
- Suihko, M.L.; Alakomi, H.L.; Gorbushina, A.; Fortune, I.; Marquardt, J.; Saarela, M. Characterization of aerobic bacterial and fungal microbiota on surfaces of historic Scottish monuments. Syst. Appl. Microbiol. 2007, 30, 494–508. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Morales, B.O.; Narváez-Zapata, J.; Reyes-Estebanez, M.; Quintana, P.; De la Rosa-García del, C.S.; Bullen, H.; Gómez-Cornelio, S.; Chan-Bacab, M.J. Bioweathering potential of cultivable fungi associated with semi-arid surface microhabitats of mayan buildings. Front. Microbiol. 2016, 7, 201. [Google Scholar] [CrossRef]
- Ruffolo, S.A.; De Leo, F.; Ricca, M.; Arcudi, A.; Silvestri, C.; Bruno, L.; Urzì, C.; La Russa, M.F. Medium-term in situ experiment by using organic biocides and titanium dioxide for the mitigation of microbial colonization on stone surfaces. Int. Biodeterior. Biodegrad. 2017, 123, 17–26. [Google Scholar] [CrossRef]
- Trovão, J.; Portugal, A.; Soares, F.; Paiva, D.S.; Mesquita, N.; Coelho, C.; Pinheiro, A.C.; Catarino, L.; Gil, F.; Tiago, I. Fungal diversity and distribution across distinct biodeterioration phenomena in limestone walls of the old cathedral of Coimbra, UNESCO World Heritage Site. Int. Biodeterior. Biodegrad. 2019, 142, 91–102. [Google Scholar] [CrossRef]
- Trovão, J.; Gil, F.; Catarino, L.; Soares, F.; Tiago, I.; Portugal, A. Analysis of fungal deterioration phenomena in the first Portuguese King tomb using a multi-analytical approach. Int. Biodeterior. Biodegrad. 2020, 149, 104933. [Google Scholar] [CrossRef]
- Mang, S.M.; Scrano, L.; Camele, I. Preliminary studies on fungal contamination of two rupestrian churches from Matera (Southern Italy). Sustainability 2020, 12, 6988. [Google Scholar] [CrossRef]
- Urzì, C.; De Leo, F.; Bruno, L.; Albertano, P. Microbial diversity in paleolithic caves: A study case on the phototrophic biofilms of the Cave of Bats (Zuheros, Spain). Microb. Ecol. 2010, 60, 116–129. [Google Scholar] [CrossRef]
- Urzì, C.; De Leo, F.; Lo Passo, C.; Criseo, G. Intra-specific diversity of Aureobasidium pullulans strains isolated from rocks and other habitats assessed by physiological methods and by random amplified polymorphic DNA (RAPD). J. Microbiol. Meth. 1999, 36, 95–105. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.M.; Nováková, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Two new species of the genus Ochroconis, O. lascauxensis and O. anomala isolated from black stains in Lascaux Cave, France. Fungal Biol. 2012, 116, 574–589. [Google Scholar] [CrossRef] [PubMed]
- Sert, H.B.; Sümbül, H.; Sterflinger, K. Sarcinomyces sideticae, a new black yeast from historical marble monuments in Side (Antalya, Turkey). Bot. J. Linn. Soc. 2007, 154, 373–380. [Google Scholar] [CrossRef]
- Isola, D.; Selbmann, L.; de Hoog, G.S.; Fenice, M.; Onofri, S.; Prenafeta-Boldú, F.X.; Zucconi, L. Isolation and screening of black fungi as degraders of volatile aromatic hydrocarbons. Mycopathologia 2013, 175, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Su, L.; Yang, S.; Sun, J.; Liu, B.; Fu, R.; Wu, B.; Liu, X.; Cai, L.; Guo, L.; et al. Unveiling the hidden diversity of rock-inhabiting fungi: Chaetothyriales from China. J. Fungi 2020, 6, 187. [Google Scholar] [CrossRef] [PubMed]
- Wollenzien, U.; de Hoog, G.S.; Krumbein, W.E.; Uijthof, J.M.J. Sarcinomyces petricola, a new microcolonial fungus from marble in the Mediterranean basin. Antonie Van Leeuwenhoek J. Microb. 1997, 71, 281–288. [Google Scholar] [CrossRef]
- Bogomolova, E.V.; Minter, D.W. A new microcolonial rock-inhabiting fungus from marble in Chersonesos (Crimea, Ukraine). Mycotaxon 2003, 86, 195–204. [Google Scholar]
- Tsuneda, A.; Hambleton, S.; Currah, R.S. The anamorph genus Knufia and its phylogenetically allied species in Coniosporium, Sarcinomyces, and Phaeococcomyces. Can. J. Bot. 2011, 89, 523–536. [Google Scholar] [CrossRef]
- Nai, C.; Wong, H.Y.; Pannenbecker, A.; Broughton, W.J.; Benoit, I.; de Vries, R.P.; Guedain, C.; Gorbushina, A.A. Nutritional physiology of a rock-inhabiting, model microcolonial fungus from an ancestral lineage of the Chaetothyriales (Ascomycetes). Fungal Genet. Biol. 2013, 56, 54–66. [Google Scholar] [CrossRef]
- Quaedvlieg, W.; Binder, M.; Groenewald, J.Z.; Summerell, B.A.; Carnegie, A.J.; Burgess, T.I.; Crous, P.W. Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Persoonia 2014, 33, 1–40. [Google Scholar] [CrossRef]
- Hao, L.; Chen, C.; Zhang, R.; Zhu, M.; Sun, G.; Gleason, M.L. A new species of Scolecobasidium associated with the sooty blotch and flyspeck complex on banana from China. Mycol. Prog. 2013, 12, 489–495. [Google Scholar] [CrossRef]
- Samerpitak, K.; Van der Linde, E.; Choi, H.J.; Gerrits van den Ende, A.H.G.; Machouart, M.; Gueidan, C.; de Hoog, G.S. Taxonomy of Ochroconis, genus including opportunistic pathogens on humans and animals. Fungal Diver. 2014, 65, 89–126. [Google Scholar] [CrossRef]
- Samerpitak, K.; Duarte, A.P.M.; Attili-Angelis, D.; Pagnocca, F.C.; Heinrichs, G.; Rijs, A.J.M.M.; Alfjorden, A.; Gerrits van den Ende, A.H.G.; Menken, S.B.J.; de Hoog, G.S. A new species of the oligotrophic genus Ochroconis (Sympoventuriaceae). Mycol. Progress 2015, 14, 6. [Google Scholar] [CrossRef]
- De Leo, F.; Criseo, G.; Urzì, C. Impact of Surrounding Vegetation and Soil on the Colonization of Marble Statues by Dematiaceous Fungi. In Proceedings of the 8th International Congress on Deterioration and Conservation of Stone, Berlin, Germany, 30 September–4 October 1996; Reiderer, Ed.; Möller Druck und Verlag GMBH: Berlin, Germany, 1996; pp. 625–630. [Google Scholar]
- Scheerer, S.; Ortega-Morales, O.; Gaylarde, C. Microbial deterioration of stone monuments--an updated overview. Adv. Appl. Microbiol. 2009, 66, 97–139. [Google Scholar] [CrossRef] [PubMed]
- De Leo, F.; Urzì, C. Fungal colonization on treated and untreated stone surfaces. In Molecular Biology and Cultural Heritage, 1st ed.; Saiz-Jimenez, C., Ed.; Routledge: London, UK, 2003. [Google Scholar] [CrossRef]
- Chertov, O.; Gorbushina, A.A.; Deventer, D. A model for microcolonial fungi growth on rock surfaces. Ecol. Model. 2004, 177, 415–426. [Google Scholar] [CrossRef]
- Urzì, C.; Realini, M. Colour changes of Noto’s calcareous sandstone as related to its colonization by microorganisms. Int. Biodeterior. Biodegrad. 1998, 42, 45–54. [Google Scholar] [CrossRef]
- Favero-Longo, S.E.; Gazzano, C.; Girlanda, M.; Castelli, D.; Tretiach, M.; Baiocchi, C.; Piervittori, R. Physical and chemical deterioration of silicate and carbonate rocks by meristematic microcolonial fungi and endolithic lichens (Chaetothyriomycetidae). Geomicrobiol. J. 2011, 28, 732–744. [Google Scholar] [CrossRef]
- Jacobson, E.S. Pathogenic roles for fungal melanins. Clin. Microbiol. Rev. 2000, 13, 708–717. [Google Scholar] [CrossRef]
- Tonon, C.; Breitenbach, R.; Voigt, O.; Turci, F.; Gorbushina, A.A.; Favero-Longo, S.E. Hyphal morphology and substrate porosity -rather than melanization- drive penetration of black fungi into carbonate substrates. J. Cult. Herit. 2021, 48, 244–253. [Google Scholar] [CrossRef]
- Sterflinger, K.; Little, B.; Pinar, G.; Pinzari, F.; de los Rios, A.; Gu, J.-D. Future directions and challenges in biodeterioration research on historic materials and cultural properties. Int. Biodeterior. Biodegrad. 2018, 129, 10–12. [Google Scholar] [CrossRef]
- UNI. Beni Culturali—Materiali Lapidei Naturali Ed Artificiali—Descrizione della Forma di Alterazione—Termini e Definizioni. Available online: http://store.uni.com/catalogo/uni-11182-2006/ (accessed on 28 February 2022).
- ICOMOS-ISCS. Illustrated Glossary on Stone Deterioration Patterns. Available online: http://iscs.icomos.org/glossary.html (accessed on 28 February 2020).
- Urzì, C.; De Leo, F. Sampling with adhesive tape strips: An easy and rapid method to monitor microbial colonization on monument surfaces. J. Microbiol. Meth. 2001, 44, 1–11. [Google Scholar] [CrossRef]
- Ding, X.; Lan, W.; Gu, J.-D. A review on sampling techniques and analytical methods for microbiota of cultural properties and historical architecture. Appl. Sci. 2020, 10, 8099. [Google Scholar] [CrossRef]
- Quan, Y.; van den Ende, B.G.; Shi, D.; Prenafeta-Boldu, F.X.; Liu, Z.; Al-Hatmi, A.M.S.; Ahmed, S.A.; Verweij, P.E.; Kang, Y.; de Hoog, G.S. A comparison of isolation methods for black fungi degrading aromatic toxins. Mycopathologia 2019, 184, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Urzi, C.; Lisi, S.; Criseo, G.; Zagari, M. Comparazione di terreni per l’enumerazione e l’isolamento di funghi deteriogeni isolati da materiali naturali. Ann. Microb. Enzymol. 1992, 42, 185–193. [Google Scholar]
- Wiktor, V.; De Leo, F.; Urzì, C.; Guyonnet, R.; Grosseau, P.; Garcia-Diaz, E. Accelerated laboratory test to study fungal biodeterioration of cementitious matrix. Int. Biodeterior. Biodegrad. 2009, 63, 1061–1065. [Google Scholar] [CrossRef]
- Urzì, C.; De Leo, F. Evaluation of the efficiency of water-repellent and biocide compounds against microbial colonization of mortars. Int. Biodeterior. Biodegrad. 2007, 60, 25–34. [Google Scholar] [CrossRef]
- Sterflinger, K.; Piñar, G. Molecular-based techniques for the study of microbial communities in artworks. In Microorganisms in the Deterioration and Preservation of Cultural Heritage; Joseph, E., Ed.; Springer: Cham, Switzerland, 2022; pp. 59–77. [Google Scholar] [CrossRef]
- Marvasi, M.; Cavalieri, D.; Mastromei, G.; Casaccia, A.; Perito, B. Omics technologies for an in-depth investigation of biodeterioration of cultural heritage. Int. Biodeterior. Biodegrad. 2019, 144, 104736. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, B.; He, Z.; Yang, X. Distribution and diversity of bacteria and fungi colonization in stone monuments analyzed by high-throughput sequencing. PLoS ONE 2016, 11, e0163287. [Google Scholar] [CrossRef]
- Cappitelli, F.; Cattò, C.; Villa, F. The control of cultural heritage microbial deterioration. Microorganisms 2020, 8, 1542. [Google Scholar] [CrossRef]
- Lo Schiavo, S.; De Leo, F.; Urzì, C. Present and future perspectives for biocides and antifouling products for stone-built cultural heritage: Ionic liquids as a challenging alternative. Appl. Sci. 2020, 10, 6568. [Google Scholar] [CrossRef]
- De Leo, F.; Jurado, V. Editorial for the special issue “Microbial Communities in Cultural Heritage and Their Control”. Appl. Sci. 2021, 11, 11411. [Google Scholar] [CrossRef]
- Martin-Sanchez, P.M.; Novákova, A.; Bastian, F.; Alabouvette, C.; Saiz-Jimenez, C. Use of biocides for the control of fungal outbreaks in subterranean environments: The case of the Lascaux Cave in France. Environ. Sci. Technol. 2012, 46, 3762–3770. [Google Scholar] [CrossRef] [PubMed]
- Cuzman, O.A.; Olmi, R.; Riminesi, C.; Tiano, P. Preliminary study on controlling black fungi dwelling on stone monuments by using a microwave heat system. Int. J. Cons. Sci. 2013, 4, 133–144. [Google Scholar]
- Gazzano, C.; Favero-Longo, S.E.; Iacomussi, P.; Piervittori, R. Biocidal effect of lichen secondary metabolites against rock-dwelling microcolonial fungi, cyanobacteria and green algae. Int. Biodeterior. Biodegrad. 2013, 84, 300–306. [Google Scholar] [CrossRef]
- Isola, D.; Bartoli, F.; Meloni, P.; Caneva, G.; Zucconi, L. Black fungi and stone heritage conservation: Ecological and metabolic assays for evaluating colonization potential and responses to traditional biocides. Appl. Sci. 2022, 12, 2038. [Google Scholar] [CrossRef]
- Chobba, M.B.; Weththimuni, M.L.; Messaoud, M.; Urzi, C.; Bouaziz, J.; De Leo, F.; Licchelli, M. Ag-TiO2/PDMS nanocomposite protective coatings: Synthesis, characterization, and use as a self-cleaning and antimicrobial agent. Prog. Org. Coat. 2021, 158, 106342. [Google Scholar] [CrossRef]
- De Leo, F.; Marchetta, A.; Capillo, G.; Germanà, A.; Primerano, P.; Schiavo, S.L.; Urzì, C. Surface active ionic liquids based coatings as subaerial anti-biofilms for stone built cultural heritage. Coatings 2021, 11, 26. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class/Order | Genera * | Substrate | Environmental and Climatic Features | Alterations Associated to Fungal Colonization | Refs |
|---|---|---|---|---|---|
| Dothideomycetes incertae sedis | Coniosporium | Calcarenite, granite, limestone, marble | Mediterranean climate, urban environment | Grayish-black patina, pitting, black spots, greenish to dark green patina, crater shaped lesions, chipping, exfoliation, sugaring, crumbling, superficial deposit, and biofilm | [37,39,43,44,45,46,47,48,49] |
| Dothideomycetes/Capnodiales incertae sedis | Capnobotryella | Limestone, marble | Mediterranean climate, continental climate, and urban environment | Black spots, crater shaped lesions, chipping, exfoliation, sugaring, crumbling, pitting, superficial deposit, and biofilm formation | [45,48,50,51,52] |
| Constantinomyces | Sandstone | Urban environment, temperate climate | Discolorations, patina | [53] | |
| Pseudotaeniolina | Marble, sandstone | Mediterranean climate, arid and desert climate | Biological green patina | [54,55,56] | |
| Dothideomycetes/Capnodiales | Aeminium | Limestone | Temperate climate | Black discoloration with salt efflorescence | [57] |
| Dothideomycetes/Cladosporiales | Cladosporium | Calcarenite, granite, limestone, marble, plaster, sandstone, tufa | Ubiquitous worldwide distribution in indoor environments and outdoor | Dark alterations, black spots, black patinas, detachment of marble grains, light grayish patina, crater shaped lesions, chipping, exfoliation, sugaring, crumbling, pitting, superficial deposit, biofilm, black crusts, green biofilm with salt efflorescence, stone erosion and disintegration, and discoloration | [27,40,46,48,49,58,59,60,61,62,63,64,65,66,67] |
| Verrucocladosporium | Limestone, marble, sandstone | Mediterranean climate, temperate climate, and urban environment | Black patina, discoloration | [37,53] | |
| Dothideomycetes/Dothideales | Aureobasidium | Granite, limestone, marble, plaster, sandstone | Urban environment, Mediterranean climate, temperate climate, indoor environment, and urban environment | Black patina, black spots, detachments, superficial deposit, biofilm, discolorations with or without salt efflorescence, black crusts, and stone erosion and disintegration | [37,40,45,49,53,63,64,65,68] |
| Dothideomycetes/Mycosphaerellales | Salinomyces | Marble, sandstone | Mediterranean climate | Black patina | [37] |
| Neocatenulostroma | Limestone, sandstone | Temperate climate, urban environment | Discolorations and/or patina, structural damage | [53] | |
| Neodevresia | Limestone, marble, plaster, tufa | Mediterranean climate | Black patina, discolorations, structural damage | [37,53,55,63] | |
| Saxophila | Marble | Mediterranean climate | Black patina | [37] | |
| Vermiconidia | Limestone, marble, travertine | Mediterranean climate, urban environment | Black patina | [37] | |
| Dothideomycetes/Neophaeothecales | Neophaeotheca | Marble | Mediterranean climate | Black patina | [37] |
| Dothideomycetes/Pleosporales | Alternaria | Calcarenite, granite, limestone, marble, plaster, tufa | Ubiquitous worldwide distribution in indoor environments and outdoor | Black spots, black patina, detachment of marble grains, greenish to dark green patina, biofilm, black crusts, green-black patina; and blackish patina | [40,46,49,58,59,60,63,64,66,67] |
| Epicoccum | Granite, limestone, marble | Urban environment, mediterranean climate, and temperate climate | Black spots, black patinas, detachment, superficial deposit, biofilm, blackish patina, green biofilm, and dark and green biofilm with salt efflorescence | [40,45,49,60,64] | |
| Phoma | Calcarenite, granite, limestone, marble, plaster, tufa | Mediterranean climate, temperate climate, urban environment, continental-cold climate, and indoor and outdoor environments | Black spots, black patinas, detachment of marble grains; color changes, crater shaped lesions, chipping and exfoliation, sugaring, crumbling, pitting, superficial deposit, biofilm, and black crusts | [40,46,48,49,58,63] | |
| Dothideomycetes/Venturiales | Ochroconis | Calcarenite | Subterranean environment | Black patina | [69] |
| Eurotiomycetes incertae sedis | Sarcinomyces | Marble | Mediterranean climate | Black spots | [70] |
| Eurotiomycetes/Chaetothyriales | Cyphellophora sp. | Plaster | Mediterranean climate | Black/grayish patina | [63] |
| Exophiala | Calcarenite, limestone, marble, sandstone | Mediterranean climate, urban environment, temperate climate, and hypogean environment | Dark alterations, black spots, black patinas, detachment of marble grains, discolorations, and visible structural damage | [27,37,40,45,53,71] | |
| Lithophila | Limestone, marble | Mediterranean climate, urban environment, and dry continental climate | Black spots, black patinas, detachment of marble grains | [37,40,72] | |
| Knufia | Limestone, marble, sandstone travertine | Mediterranean climate, urban environment, continental temperate climate, and dry continental climate | Black and grey spots, dark macropitting, biopitting, crater shaped lesions, chipping, exfoliation, sugaring, crumbling, discolorations, patina, and visible structural damage | [37,41,43,45,48,53,72,73,74] | |
| Rhinocladiella | Marble | Mediterranean climate | Black spots, crater shaped lesions, chipping and exfoliation, sugaring, crumbling, and pitting | [48] | |
| Eurotiomycetes/Mycocaliciales | Mycocalicium | Marble | Mediterranean climate, urban environment | Black spots, crater shaped lesions, chipping and exfoliation, sugaring, crumbling, and pitting | [45,48] |
| Taxon | Strain | ITS rDNA |
|---|---|---|
| Capnobotryella antalyensis | MA 4615 | AJ972858 |
| Capnobotryella antalyensis | MA 4624 | AJ972850 |
| Capnobotryella antalyensis | MA 4766 | AJ972851 |
| Capnobotryella antalyensis | MA 4775 | AJ972860 |
| Capnobotryella isilogui | MA 3619 | AM746201 |
| Capnobotryella erdogani | MA 4625 | AJ972857 |
| Capnobotryella kirizoglui | MA 4899 | AJ972859 |
| Capnobotryella sp. | MA 4701 | AJ972856 |
| Capnobotryella sp. | MA 4697 | AJ972855 |
| Capnobotryella sp. | MA 3615 | AM746203 |
| Neodevriesia modesta | CCFEE 5672 | KF309984 |
| Neodevriesia simplex | CCFEE 5681 | KF309985 |
| Neodevriesia sardiniae | CCFEE 6202 | KP791765 |
| Neodevriesia sardiniae | CCFEE 6210 | KP791766 |
| Saxophila tyrrhenica | CCFEE 5935 | KP791764 |
| Aeminium ludgeri | E12 | MG938054 |
| Aeminium ludgeri | E16 | MG938061 |
| Neocatenulostroma sp. | CR1 | KY111907 |
| Constantinomyces sp. | CR21 | KY111911 |
| Pseudaeniolina globosa | DPS10 | MH396690 |
| Pseudotaeniolina globosa | CBS109889 | NR136960 |
| Pseudotaeniolina globosa | CCFEE5734 | KF309976 |
| Vermiconidia calcicola | CBS 140080 | NR_145012 |
| Vermiconidia calcicola | CCFEE 5780 | KP791761 |
| Vermiconidia flagrans | CCFEE 5922 | KP791753 |
| Coniosporium uncinatum | CBS 100219 | AJ244270 |
| Coniosporium apollinis | CBS 100213 | AJ244271 |
| Coniosporium apollinis | CBS 352.97 | NR159787 |
| Coniosporium apollinis | CBS 100216 | AJ244272 |
| Coniosporium apollinis | QIIIa | MH023395 |
| Lithophila catenulata | BJ10118 | JN650519 |
| Lithophila guttulata | M1 | MW361305 |
| Lithophila guttulata | CCFEE 5884 | KP791768 |
| Lithophila guttulata | CCFEE 5907 | KP791773 |
| Knufia mediterranea | CCFEE 5738 | KP791791 |
| Knufia mediterranea | CCFEE 6211 | KP791793 |
| Knufia vaticanii | CCFEE 5939 | KP791780 |
| Knufia calcarecola | SL11033 | JQ354925 |
| Knufia calcarecola | CGMCC 3.17222 | KP174862 |
| Knufia marmoricola | CCFEE 5895 | KP791775 |
| Knufia marmoricola | CCFEE 5716 | KP791786 |
| Knufia perforans | CBS 885.95 | AJ244230 |
| Knufia karalitana | CCFEE 5732 | KP791782 |
| Knufia karalitana | CCFEE 5929 | KP791783 |
| Knufia petricola | CCFEE 726.95 | KC978746 |
| Knufia petricola | CBS 600.93 | KC978744 |
| Knufia petricola | IMI38917 | AJ507323 |
| Knufia petricola | D1 | JF749183 |
| Knufia petricola | M4 | FJ556910 |
| Knufia sp. | QIIa | MH023393 |
| Knufia sp. | QIIb | MH023394 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Leo, F.; Marchetta, A.; Urzì, C. Black Fungi on Stone-Built Heritage: Current Knowledge and Future Outlook. Appl. Sci. 2022, 12, 3969. https://doi.org/10.3390/app12083969
De Leo F, Marchetta A, Urzì C. Black Fungi on Stone-Built Heritage: Current Knowledge and Future Outlook. Applied Sciences. 2022; 12(8):3969. https://doi.org/10.3390/app12083969
Chicago/Turabian StyleDe Leo, Filomena, Alessia Marchetta, and Clara Urzì. 2022. "Black Fungi on Stone-Built Heritage: Current Knowledge and Future Outlook" Applied Sciences 12, no. 8: 3969. https://doi.org/10.3390/app12083969
APA StyleDe Leo, F., Marchetta, A., & Urzì, C. (2022). Black Fungi on Stone-Built Heritage: Current Knowledge and Future Outlook. Applied Sciences, 12(8), 3969. https://doi.org/10.3390/app12083969