
Response of Rhizosphere Microbial Community in High-PAH-Contaminated Soil Using Echinacea purpurea (L.) Moench


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Extraction and Analysis of PLFAs
2.3. Data Analysis
2.4. PLFA Nomenclature
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tao, Y.; Li, Y. Study on the sources and monitoring analysis methods of polycyclic aromatic hydrocarbons (PAHs) in the environment. Environ. Dev. 2019, 31, 96–98. (In Chinese) [Google Scholar]
- Meudec, A.; Dussauze, J.; Jourdin, M.; Deslandes, E.; Poupart, N. Gas chorolatographic-mass spectrometric method for polycyclic aromatic hydrocarbo n analysis in plant biota. J. Chromatogr. A 2006, 1108, 240–247. [Google Scholar] [CrossRef]
- Zhou, Q.X.; Song, Y. Contaminated Soil Remediation: Principles and Methods; Science Press: Beijing, China, 2004. (In Chinese) [Google Scholar]
- Zhou, Q.X.; Sun, F.H.; Liu, R. Joint chemical flushing of soils contaminated with petroleum hydrocarbons. Environ. Int. 2005, 31, 835–839. [Google Scholar] [CrossRef] [PubMed]
- Fan, S.; Li, P.; He, N.; Ren, W.; Zhang, H.; Xu, H. Research of Phytoremediation on Contaminated Soil with Polycyclic Aromatic Hydrocarbons (PAHs). J. Agro-Environ. Sci. 2007, 26, 2007–2013. [Google Scholar]
- Chen, N.; Zheng, Y.; He, X.; Li, X.; Zhang, X. Analysis of “National Survey Bulletin of Soil Pollution Status”. J. Agric. Environ. Sci. 2017, 36, 1689–1692. [Google Scholar]
- Liu, R.; Jadeja, R.N.; Zhou, Q.; Liu, Z. Treatment and remediation of petroleum-contaminated soils using selective orna mental plants. Environ. Eng. Sci. 2012, 29, 494–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J. The Effects of Plants, Carbon Sources and Bacteria on the Remedation of PAHs Contaminated Soils and Water. Master’s Thesis, Zhejiang University, Hangzhou, China, 2012. (In Chinese). [Google Scholar]
- Zhang, X.; Xu, L.; Qi, Y.; Sun, L.; Zhang, J. Effect of purple pine nut chrysanthemum on remediation of soil contaminated with polycyclic aromatic hydrocarbons. J. Ecol. 2018, 37, 492–497. [Google Scholar]
- Abbaspour, A.; Zohrabi, F.; Dorostkar, V.; Faz, A.; Acosta, J.A. Remediation of an oil-contaminated soil by two native plants treated with biochar and mycorrhizae. J. Environ. Manag. 2020, 254, 109755. [Google Scholar] [CrossRef]
- Frenk, S.; Hadar, Y.; Minz, D. Resilience of soil bacterial community to irrigation with water of different qualities under Mediterranean climate. J. Environ. Microbiol. 2014, 16, 559–569. [Google Scholar] [CrossRef] [PubMed]
- Zornoza, R.; Guerrero, C.; Mataix-Solera, J.; Scow, K.M.; Arcenegui, V.; Mataix-Beneyto, J. Changes in soil microbial community structure following the abandonment of agricultural terraces in mountainous areas of Eastern Spain. Appl. Soil Ecol. 2009, 42, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Liu, Y.; Tao, Q.; Hou, Q.; Wu, K.; Song, Y.; Liu, Y.; Guo, X.; Li, J.; Muhammad, L.R.H.; et al. Successive phytoextraction alters ammonia oxidation and associated microbial communities in heavy metal contaminated agricultural soils. Sci. Total Environ. 2019, 664, 616–625. [Google Scholar] [CrossRef]
- Francisco, R.; Stone, D.; Creamer, R.E.; Sousa, J.P.; Morais, P.V. European scale analysis of phospholipid fatty acid composition of soils to establish operating ranges. Appl. Soil Ecol. 2016, 97, 49–60. [Google Scholar] [CrossRef]
- Ganzert, L.; Lipski, A.; Hubberten, H.-W.; Wagner, D. The impact of different soil parameters on the community structure of dominant bacteria from nine different soils located on Livingston Island, South Shetland Archipelago, Antarctica. FEMS Microbiol. Ecol. 2011, 76, 476–491. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Rajkumar, M.; Oliveira, R.S.; Zhang, C.; Freitas, H. Potential of plant beneficial bacteria and arbuscular mycorrhizal fungi in phytoremediation of metal-contaminated saline soils. J. Hazard. Mater. 2019, 379, 120813. [Google Scholar] [CrossRef]
- Xiao, N.; Liu, R.; Jin, C.; Dai, Y. Efficiency of five ornamental plant species in the phytoremediation of polycyclic aromatic hydrocarbon (PAH)-contaminated soil. Ecol. Eng. 2015, 75, 384–391. [Google Scholar] [CrossRef]
- Dai, Y.; Liu, R.; Zhou, Y.; Li, N.; Ma, Q.; Gao, B. Fire Phoenix facilitates phytoremediation of PAH-Cd co-contaminated soil through promotion of beneficial rhizosphere bacterial communities. Environ. Int. 2019, 136, 105421. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Ding, N.; Wang, G.; Xu, J.; Wu, J.; Philip, C.B. Effects of different soil weights, storage times and extraction methods on soil phospholipid fatty acid analyses. Geoderma 2009, 150, 171–178. [Google Scholar] [CrossRef]
- Frostegård, A.; Tunlid, A.; Bååth, E. Use and misuse of PLFA measurements in soils. Soil Biol. Biochem. 2010, 43, 1621–1625. [Google Scholar] [CrossRef]
- Piotrowska-Seget, Z.; Cycoń, M.; Kozdrój, J. Metal-tolerant bacteria occurring in heavily polluted soil and mine spoil. Appl. Soil Ecol. 2005, 28, 237–246. [Google Scholar] [CrossRef]
- Kaiser, C.; Koranda, M.; Kitzler, B.; Fuchslueger, L.; Schnecker, J.; Schweiger, P.; Rasche, F.; Zechmeister-Boltenstern, S.; Sessitsch, A.; Richter, A. Belowground carbon allocation by trees drives seasonal patterns of extracellular enzyme activities by altering microbial community composition in a beech forest soil. New Phytol. 2010, 187, 843–858. [Google Scholar] [CrossRef] [Green Version]
- Ruess, L.; Chamberlain, P.M. The fat that matters: Soil food web analysis using fatty acids and their carbon stable isotope signature. Soil Biol. Biochem. 2010, 42, 1898–1910. [Google Scholar] [CrossRef]
- Buyer, J.S.; Teasdale, J.R.; Roberts, D.P.; Zasada, I.A.; Maul, J.E. Factors affecting microbial community structure in tomato cropping systems. Soil Biol. Biochem. 2010, 42, 831–841. [Google Scholar] [CrossRef]
- Kaur, A.; Chaudhary, A.; Kaur, A.; Choudhary, R.; Kaushik, R. Phospholipid fatty acid—A bioindicator of environment monitoring and assessment in soil ecosystem. Curr. Sci. 2005, 89, 1103–1112. [Google Scholar]
- Feng, Y.; Motta, A.C.; Reeves, D.W.; Burmester, C.H.; van Santen, E.; Osborne, J.A. Soil microbial communities under conventional-till and no-till continuous cotton systems. Soil Biol. Biochem. 2003, 35, 1693–1703. [Google Scholar] [CrossRef]
- Wolf, D.C.; Cryder, Z.; Gan, J. Soil bacterial community dynamics following surfactant addition and bioaugmentation in pyrene-contaminated soils. Chemosphere 2019, 231, 93–102. [Google Scholar] [CrossRef]
- Mustafa, S.; Kabir, S.; Shabbir, U.; Batool, R. Plant growth promoting rhizobacteria in sustainable agriculture: From theoretical to pragmatic approach. Symbiosis 2019, 78, 115–123. [Google Scholar] [CrossRef]
- Suresh, B.; Ravishankar, G.A. Phytoremediation—A novel and promising approach for environmental clean-up. Crit. Rev. Biotechnol. 2004, 24, 97–124. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Li, Q.; Ling, W.; Zhu, X. Arbuscular mycorrhizal phytoremediation of soils contaminated with phenanthrene and pyrene. J. Hazard. Mater. 2011, 185, 703–709. [Google Scholar] [CrossRef]
- Slađana, Č.A.; Biljana, S.M.; Vesna, B.R. How can plants manage polycyclic aromatic hydrocarbons? May these effects represent a useful tool for an effective soil remediation? A review. Clean Technol. Environ. Policy 2015, 17, 597–614. [Google Scholar]
- Pagé, A.P.; Yergeau, E.; Greer, C.W. Salix purpurea stimulates the expression of specific bacterial xenobiotic degradation genes in a soil contaminated with hydrocarbons. PLoS ONE 2015, 10, e0132062. [Google Scholar] [CrossRef] [Green Version]
- Phillips, L.A.; Greer, C.W.; Farell, R.E.; Germida, J.J. Plant root exudates impact the hydrocarbon degradation potential of a weathered-hydrocarbon contaminated soil. Appl. Soil Ecol. 2012, 52, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Oleszczuk, P.; Rakowska, M.; Bucheli, T.D.; Godlewska, P.; Reible, D.D. Combined effects of plant cultivation and sorbing carbon amendments on freely dissolved PAHs in contaminated soil. Environ. Sci. Technol. 2019, 53, 4860–4868. [Google Scholar] [CrossRef] [PubMed]
- Bakker, P.A.H.M.; Pieterse, C.M.J.; de Jonge, R.; Berendsen, R.L. The Soil-Borne Legacy. Cell 2018, 172, 1178–1180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berendsen, R.L.; Vismans, G.; Yu, K.; Song, Y.; de Jonge, R.; Burgman, W.P.; Burmølle, M.; Herschend, J.; Bakker, P.A.H.M.; Pieterse, C.M.J. Disease-induced assemblage of a plant-beneficial bacterial consortium. ISME J. 2018, 12, 1496–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pineda, A.; Kaplan, I.; Bezemer, T.M. Steering Soil Microbiomes to Suppress Aboveground Insect Pests. Trends Plant Sci. 2017, 22, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L. New Favorite of Root Flower—Pine Cone. Chin. Flower Hortic. 2013, 24. (In Chinese) [Google Scholar]
- Cheng, Z.; Ling, W.; Gao, Y.; Wang, J. Impacts of arbuscular mycorrhizal on plant uptake and phytoremediation of pyrene in soils. Plant Nutr. Fert. Sci. 2008, 14, 1178–1185. [Google Scholar]
- Joner, E.J.; Leyval, C. Rhizosphere gradients of polycyclic aromatic hydrocarbon (PAH) dissipation in two industrial soils and the impact of arbuscular mycorrhiza. Environ. Sci. Technol. 2003, 37, 2371–2375. [Google Scholar] [CrossRef]
- Chen, R.; Lin, X.; Yin, R. Effect of mycorrhizae on bioremediation of soil polluted by organic matters. Chin. J. Ecol. 2005, 24, 176–180. [Google Scholar]
- Liu, R.; Zhao, L.; Jin, C.; Xiao, N.; Jadeja, R.N.; Sun, T. Enzyme responses to phytoremediation of PAH-contaminated soil using Echinacea purpurea. Water Air Soil Pollut. 2014, 225, 2230.48. [Google Scholar] [CrossRef]
- Xia, J.; Li, J. Advances in Increasing Plant Drought Resistance by AM Fungi. Chin. Agric. Sci. Bull. 2005, 2, 625014. [Google Scholar]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Fanin, N.; Kardol, P.; Farrell, M.; Nilsson, M.C.; Gundale, M.J.; Wardle, D.A. The ratio of Gram-positive to Gram-negative bacterial PLFA markers as an indicator of carbon availability in organic soils. Soil Biol. Biochem. 2019, 128, 111–114. [Google Scholar] [CrossRef]
- Kumari, S.; Mangwani, N.; Das, S. Synergistic effect of quorum sensing genes in biofilm development and PAHs degradation by a marine bacterium. Bioengineered 2016, 7, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houlden, A.; Timms-Wilson, T.M.; Day, M.J.; Bailey, M.J. Influence of plant developmental stage on microbial community structure and activity in the rhizosphere of three field crops. FEMS Microbiol. Ecol. 2008, 65, 193–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Micallef, S.A.; Channer, S.; Shiaris, M.P.; Colón-Carmona, A. Plant age and genotype impact the progression of bacterial community succession in the Arabidopsis rhizosphere. Plant Signal. Behav. 2009, 4, 777–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na, X.; Li, X.; Zhang, Z.; Li, M.; Kardol, P.; Xu, T.; Wang, M.; Cao, X.; Ma, F. Bacterial community dynamics in the rhizosphere of a long-lived, leguminous shrub across a 40-year age sequence. J. Soils Sediments 2017, 18, 76–84. [Google Scholar] [CrossRef]
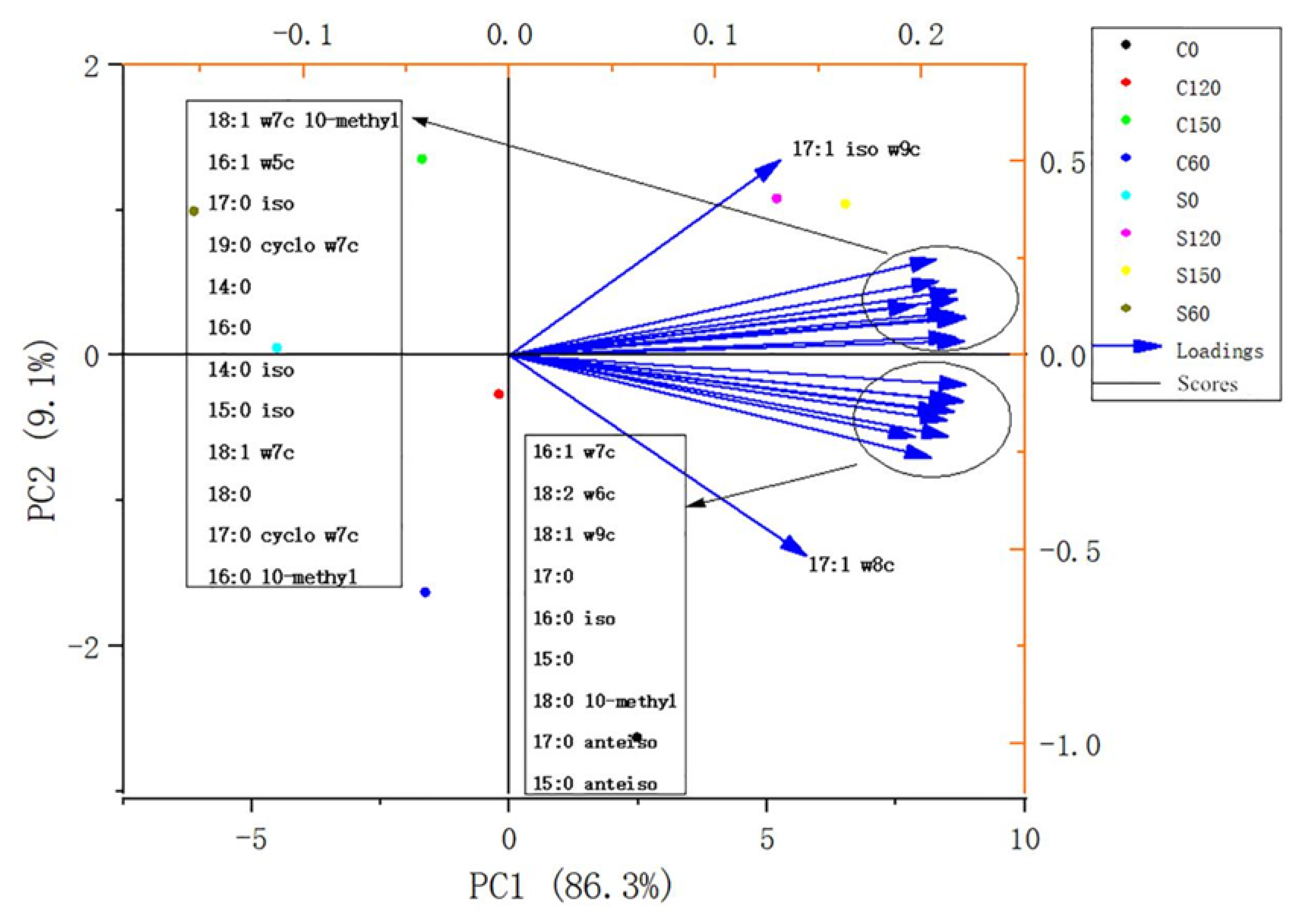

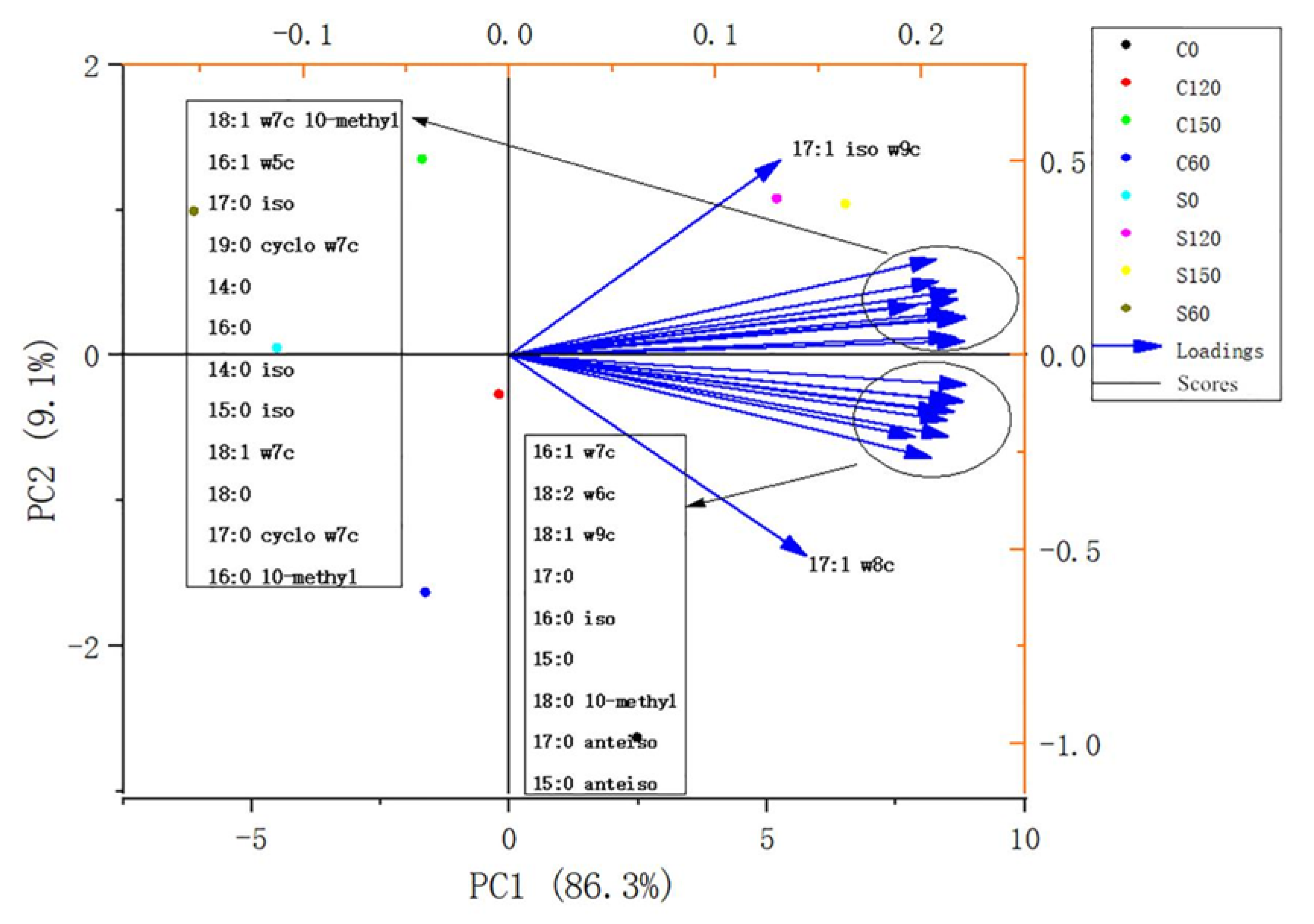{kind=link}
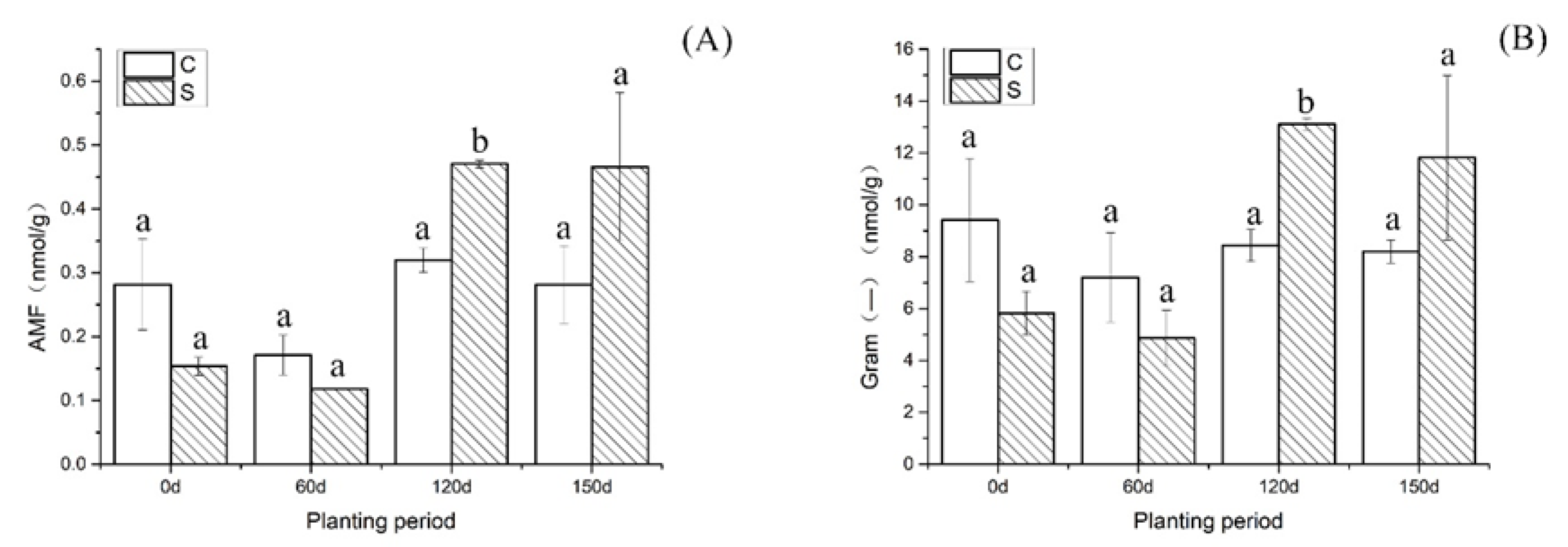
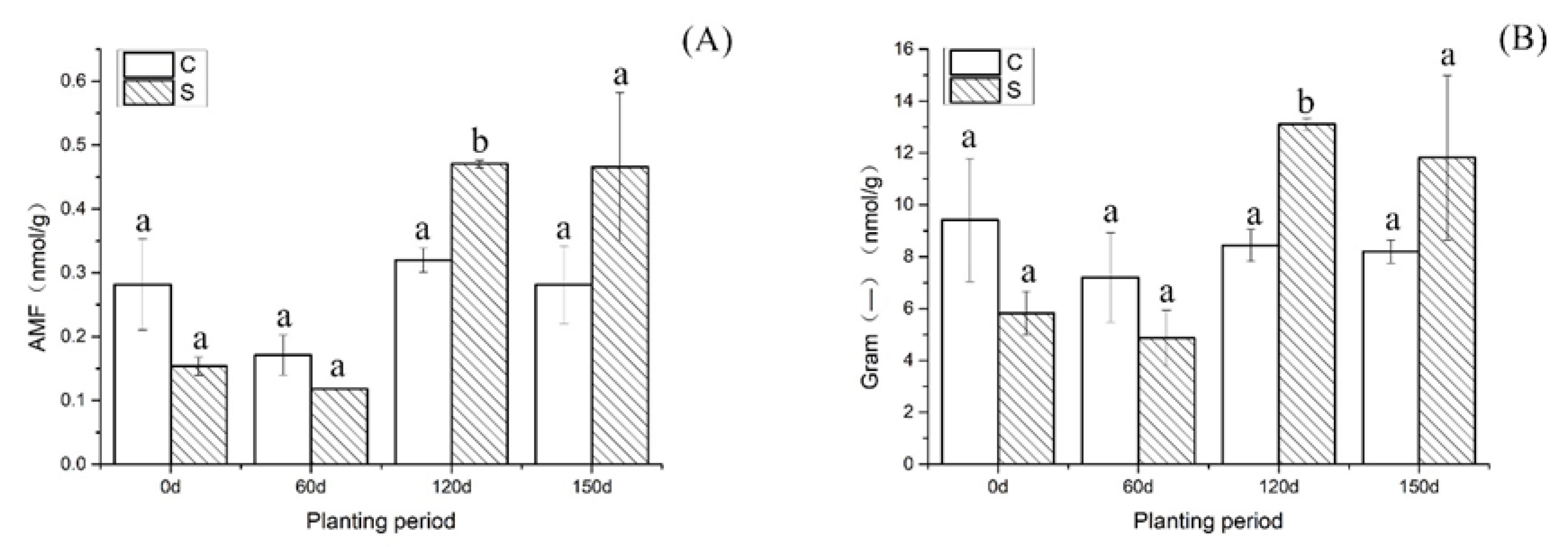{kind=link}
| Biomarkers in Soil in Different Periods before and after E. purpurea Planting (nmol/g) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Planting Period (d) | 0 | 60 | 120 | 150 | ||||
| C | S | C | S | C | S | C | S | |
| ACT | 1.18 (a) | 0.54 (a) | 0.96 (a) | 0.40 (a) | 1.00 (a) | 1.20 (b) | 0.82 (a) | 1.39 (a) |
| SD | 0.15 | 0.07 | 0.18 | 0.10 | 0.07 | 0.43 | 0.16 | 0.3 |
| G+ | 4.34 (ac) | 2.27 (ac) | 3.20 (ac) | 1.76 (ac) | 3.59 (ac) | 6.40 (ac) | 2.95 (ac) | 8.27 (ac) |
| SD | 0.90 | 0.37 | 0.73 | 0.01 | 0.09 | 0.10 | 0.22 | 5.21 |
| G− | 9.42 (ac) | 5.82 (ac) | 7.21 (ac) | 4.86 (ac) | 8.14 (ac) | 13.11 (bd) | 7.43 (ac) | 13.53 (ac) |
| SD | 2.37 | 0.84 | 1.72 | 0.03 | 0.62 | 0.23 | 0.45 | 3.17 |
| BAC | 16.38 (ac) | 10.13 (ac) | 12.70 (ac) | 8.55 (ac) | 14.12 (ac) | 20.00 (bc) | 12.43 (ac) | 27.22 (ac) |
| SD | 3.53 | 1.32 | 2.56 | 0.004 | 0.77 | 0.12 | 0.74 | 7.49 |
| FUN | 2.25 (ac) | 1.20 (ac) | 1.70 (ac) | 0.93 (ad) | 1.76 (ac) | 2.47 (bd) | 1.47 (ac) | 2.36 (ac) |
| SD | 0.60 | 0.20 | 0.38 | 0.009 | 0.07 | 0.07 | 0.14 | 0.51 |
| AMF | 0.28 (a) | 0.15 (a) | 0.17 (a) | 0.11 (a) | 0.33 (a) | 0.47 (b) | 0.31 (a) | 0.46 (a) |
| SD | 0.07 | 0.01 | 0.03 | 0.0003 | 0.004 | 0.01 | 0.03 | 0.12 |
| B/F | 7.35 (a) | 8.47 (a) | 7.51 (a) | 9.29 (a) | 8.06 (a) | 8.55 (a) | 8.43 (a) | 9.65 (a) |
| SD | 0.41 | 0.32 | 0.45 | 0.11 | 0.08 | 0.22 | 0.32 | 1.69 |
| G−/G+ | 2.16 (a) | 2.57 (a) | 2.28 (a) | 2.78 (a) | 2.27 (a) | 2.14 (b) | 2.53 (a) | 2.13 (b) |
| SD | 0.23 | 0.14 | 0.38 | 0.05 | 0.11 | 0.07 | 0.03 | 0.07 |
| TOT | 29.72 (a) | 18.22 (a) | 22.72 (a) | 13.38 (a) | 26.69 (a) | 46.14 (b) | 24.31 (a) | 41.07 (a) |
| SD | 7.07 | 2.90 | 4.99 | 0.10 | 1.52 | 0.61 | 1.66 | 10.80 |
| Biomarker Group | PLFAs | References |
|---|---|---|
| Actinomycetes | 16:0 10-methyl, 18:0 10-methyl | Kaiser et al., 2010 [22] |
| Gram-positive bacteria | 14:0 iso, 15:0 iso, 16:0 iso, 17:0 iso, 17:0 anteiso, 15:0 anteiso | Kaiser et al., 2010; Ruess and Chamberlain, 2010 [23]; Buyer and Sasser, 2012 [24]; |
| Gram-negative bacteria | 16:1 w7c, 18:1 w7c, 17:1 w8c | Kaur et al., 2005 [25] |
| Bacteria | Gram-positive and Gram-negative biomarkers, 14:0, 15:0, 17:0, 18:0, 16:0, 18:1 w7c 10-methyl, 17:1 iso w9c, 19:0 cyclo w7c, 17:0 cyclo w7c | Kaur et al., 2005; Feng et al., 2003 [26]; Kaiser et al., 2010 |
| Fungi | 18:1 w9c, 18:2 w6c | Ruess and Chamberlain, 2010; Buyer and Sasser, 2012; |
| Arbuscular mycorrhizal fungi | 16:1 w5c | Buyer et al., 2010 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, K.; Liu, R.; Xiao, Y.; Song, M.; Deng, X.; Dai, T.; Wang, Y.; Wu, X. Response of Rhizosphere Microbial Community in High-PAH-Contaminated Soil Using Echinacea purpurea (L.) Moench. Appl. Sci. 2022, 12, 2973. https://doi.org/10.3390/app12062973
Liu K, Liu R, Xiao Y, Song M, Deng X, Dai T, Wang Y, Wu X. Response of Rhizosphere Microbial Community in High-PAH-Contaminated Soil Using Echinacea purpurea (L.) Moench. Applied Sciences. 2022; 12(6):2973. https://doi.org/10.3390/app12062973
Chicago/Turabian StyleLiu, Kaikai, Rui Liu, Yao Xiao, Mengting Song, Xingyu Deng, Tingting Dai, Yu Wang, and Xin Wu. 2022. "Response of Rhizosphere Microbial Community in High-PAH-Contaminated Soil Using Echinacea purpurea (L.) Moench" Applied Sciences 12, no. 6: 2973. https://doi.org/10.3390/app12062973
APA StyleLiu, K., Liu, R., Xiao, Y., Song, M., Deng, X., Dai, T., Wang, Y., & Wu, X. (2022). Response of Rhizosphere Microbial Community in High-PAH-Contaminated Soil Using Echinacea purpurea (L.) Moench. Applied Sciences, 12(6), 2973. https://doi.org/10.3390/app12062973