
Improved Gamma-Decalactone Synthesis by Yarrowia lipolytica Yeast Using Taguchi Robust Design Method




Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Microorganism and Culture Conditions
2.3. Gamma-Decalactone Extraction and Quantification
2.4. Determination of Biomass Yield
2.5. Growth Parameters
2.6. Hydrophobicity of Cells
2.7. Statistical Analysis
3. Results
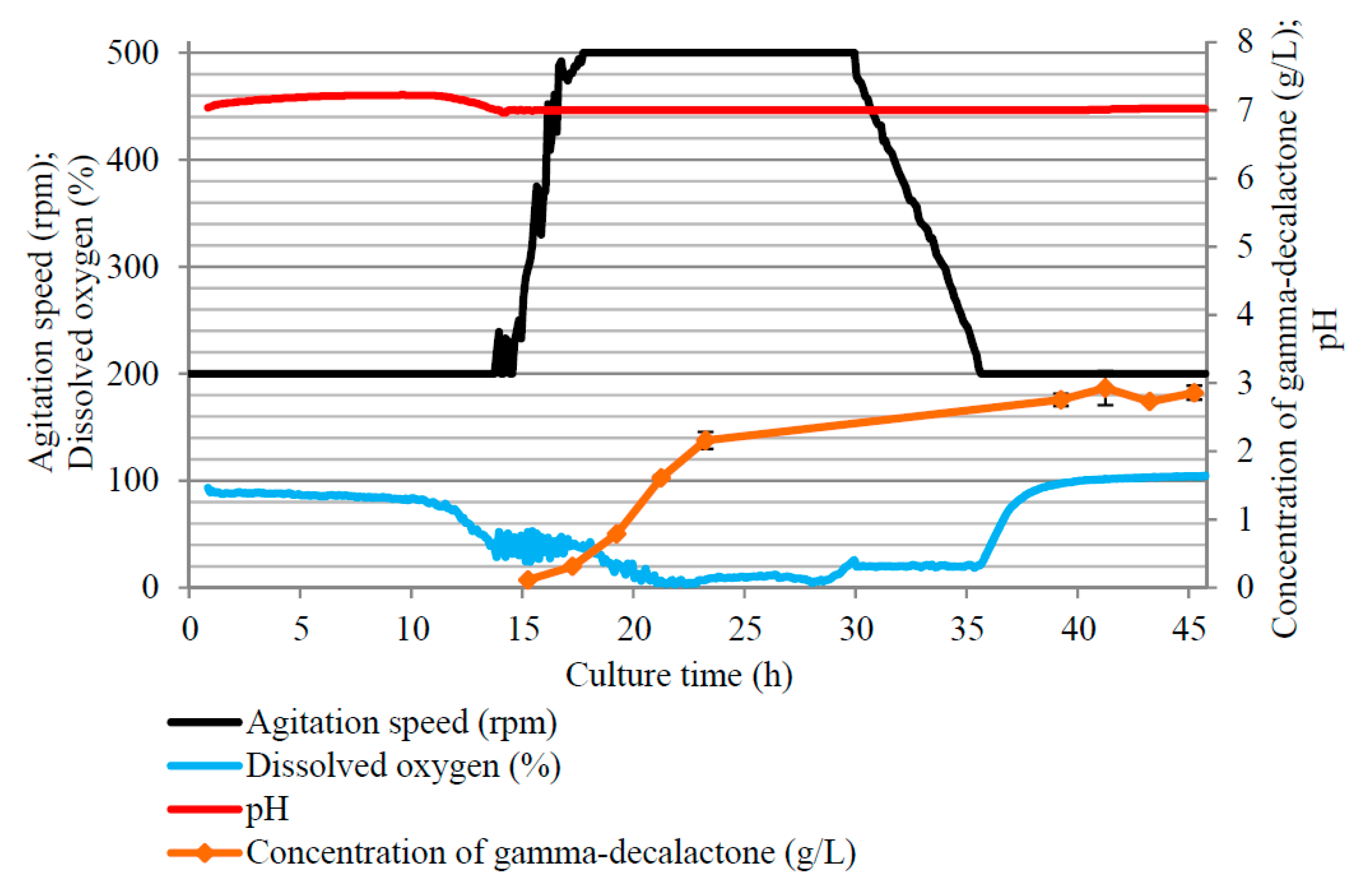
3.1. Impact of Concentration of Castor Oil, pH of the Medium, and Agitation Speed on Synthesis of γ-Decalactone in the Batch Culture Carried Out in the Bioreactor
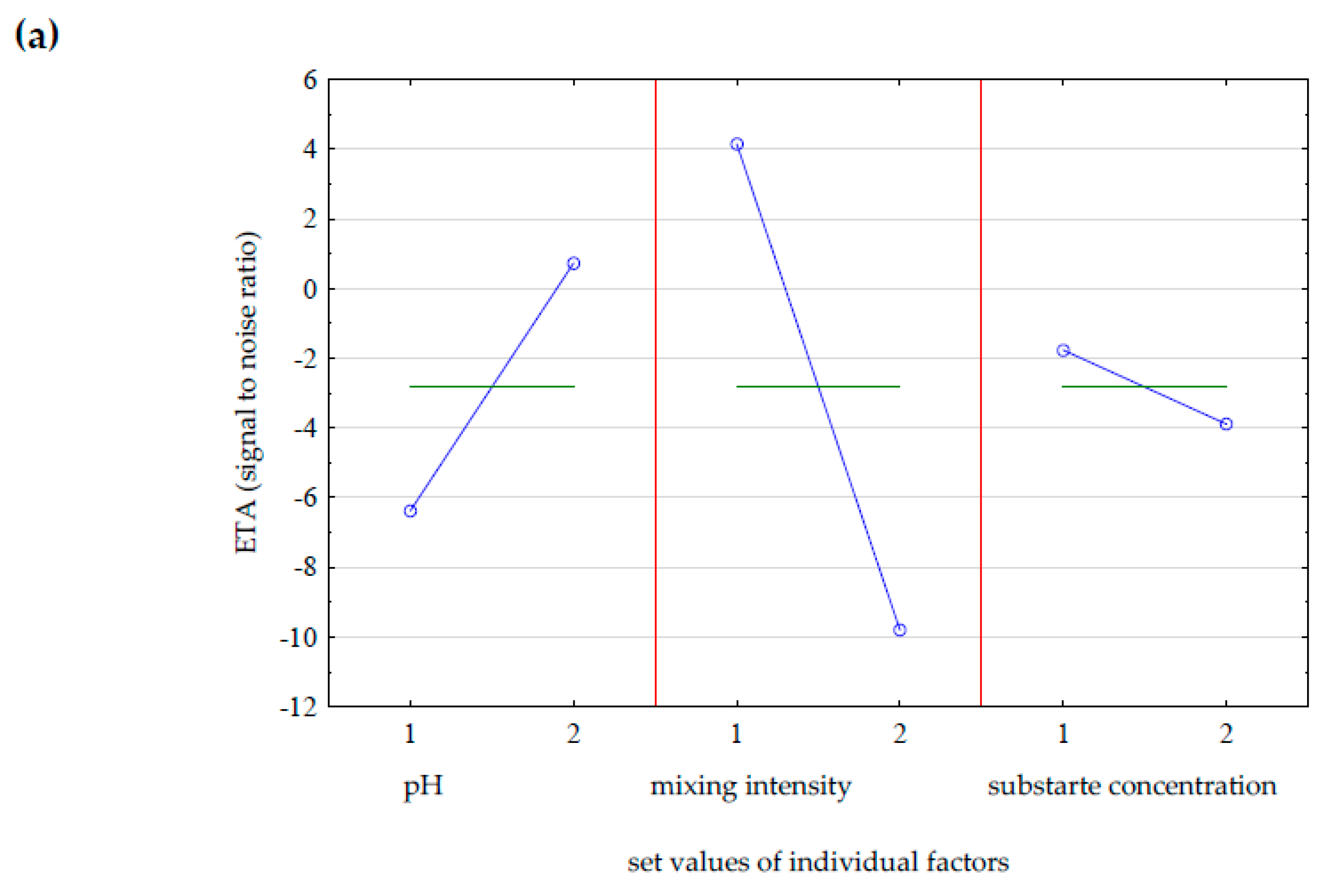
3.2. Evaluation of Optimal Conditions for γ-Decalactone Biosynthesis in Bioreactor Culture Means of the Taguchi Statistical Design Method
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- GlobeNewswire: Food Flavors Market to Reach USD 19.72 Billion by 2026. Available online: https://www.globenewswire.com/news-release/2019/10/10/1928250/0/en/Food-Flavors-Market-To-Reach-USD-19-72-Billion-By-2026-Reports-And-Data.html (accessed on 15 August 2022).
- Flavors and Fragrances Market Size, Share & Trends Analysis Report, 2019–2025. Available online: https://www.grandviewresearch.com/industry-analysis/flavors-fragrances-market (accessed on 15 August 2022).
- Braga, A.; Guerreiro, C.; Belo, I. Generation of flavors and fragrances through biotransformation and de novo synthesis. Food Bioprocess Technol. 2018, 11, 2217–2228. [Google Scholar] [CrossRef]
- Sales, A.; Paulino, B.N.; Pastore, G.M.; Bicas, J.L. Biogeneration of aroma compounds. Curr. Opin. Food Sci. 2018, 19, 77–84. [Google Scholar] [CrossRef]
- Paulino, B.N.; Sales, A.; Felipe, L.; Pastore, G.M.; Molina, G.; Bicas, J.L. Recent advances in the microbial and enzymatic production of aroma compounds. Curr. Opin. Food Sci. 2021, 37, 98–106. [Google Scholar] [CrossRef]
- Blin-Perrin, C.; Molle, D.; Duffose, L.; Le-Quere, J.L.; Viel, C.; Mauvais, G.; Feron, G. Metabolism of ricinoleic acid into γ-decalactone: β-oxidation and long chain acyl intermediates of ricinoleic acid in the genus Sporidiobolus sp. FEMS Microbiol. Lett. 2000, 188, 69–74. [Google Scholar] [CrossRef]
- Romero-Guido, C.; Belo, I.; Ta, T.M.N.; Cao-Hoang, L.; Alchihab, M.; Gomes, N.; Thonart, P.; Teixeira, J.A.; Destain, J.; Waché, Y. Biochemistry of lactone formation in yeast and fungi and its utilisation for the production of flavor and fragrance compounds. Appl. Microbiol. Biotechnol. 2011, 89, 535–547. [Google Scholar] [CrossRef]
- AL Mualad, W.N.A.; Bouchedja, D.N.; Selmania, A.; Maadadi, R.; Ikhlef, A.; Kabouche, Z.; Elmechta, L.; Boudjellal, A. Yeast Yarrowia lipolytica as a biofactory for the production of lactone-type aroma gamma-decalactone using castor oil as substrate. Chem. Pap. 2022. [Google Scholar] [CrossRef]
- Małajowicz, J.; Kozłowska, M. Factors affecting the yield in formation of fat-derived fragrance compounds by Yarrowia lipolytica yeast. Appl. Sci. 2021, 11, 9843. [Google Scholar] [CrossRef]
- Okui, S.; Uchiyama, M.; Mizugaki, M. Metabolism of hydroxy fatty acids: 2 Intermediates of the oxidative breakdown of ricinoleic acid by Genus Candida. J. Biochem. 1963, 54, 536–540. [Google Scholar] [CrossRef]
- Guan, S.; Rong, S.; Wang, M.; Cai, B.; Li, Q.; Zhang, S. Enhanced biotransformation productivity of gamma-decalactone from ricinoleic acid based on the expanded vermiculite delivery system. J. Microbiol. Biotechnol. 2019, 29, 1071–1077. [Google Scholar] [CrossRef]
- Rong, S.; Yang, S.; Li, Q.; Cai, B.; Guan, S.; Wang, J.; Zhou, Y.; Chen, Y. Improvement of γ-decalactone production by stimulating the import of ricinoleic acid and suppressing the degradation of γ-decalactone in Saccharomyces cerevisiae. Biocatalal. Biotransfor. 2017, 35, 96–102. [Google Scholar] [CrossRef]
- Małajowicz, J.; Nowak, D.; Fabiszewska, A.; Iuliano, A. Comparison of gamma-decalactone biosynthesis by yeast Yarrowia lipolytica MTLY40-2p and W29 in batch-cultures. Biotechnol. Biotechnol. Equip. 2020, 34, 330–340. [Google Scholar] [CrossRef]
- Narayana, A.V.; Sumalatha, B.; Swamy, A.V.N.; Chary, G.H.V.C. Production of γ-decalactone through fermentation: A review. Int. J. Mod. Agric. 2021, 10, 2189–2192. [Google Scholar]
- Alchihab, M.; Aldric, J.M.; Aguedo, M.; Destain, J.; Wathelet, J.P.; Thonart, P. The use of Macronet resins to recover γ-decalactone produced by Rhodotorula aurantiaca from the culture broth. J. Ind. Microbiol. Biotechnol. 2010, 37, 167–172. [Google Scholar] [CrossRef]
- Kothari, S.D.; Vadgama, R.N.; Bhat, K.H.; Lali, A.M.; Odaneth, A.A. Process optimization for production and purification of γ-decalactone from ricinoleic acid using Yarrowia lipolytica. Biocatal. Agric. Biotechnol. 2022, 39, 1–11. [Google Scholar] [CrossRef]
- Małajowicz, J.; Górska, A.; Bryś, J.; Ostrowska-Ligęza, E.; Wirkowska-Wojdyła, M. Attempt to develop an effective method for the separation of gamma-decalactone from biotransformation medium. Appl. Sci. 2022, 12, 2084. [Google Scholar] [CrossRef]
- Alchihab, M.; Destain, J.; Aguedo, M.; Thonart, P. Production d’arômes de type lactone par des levures. Biotechnol. Agron. Soc. Environ. 2010, 14, 681–691. [Google Scholar]
- Lee, S.L.; Lin, S.J.; Chou, C.C. Growth of and production of γ-decalactone by Sporobolomyces odorus in jar fermentors as affected by pH, aeration and fed-batch technique. J. Ferment. Bioeng. 1995, 80, 195–199. [Google Scholar] [CrossRef]
- Alchihab, M.; Destain, J.; Aguedo, M.; Majad, L.; Ghalfi, H.; Wathelet, J.P.; Thonart, P. Production of γ-decalactone by a psychrophilic and a mesophilic strain of the yeast Rhodotorula aurantiaca. Appl. Biochem. Biotechnol. 2009, 158, 41–50. [Google Scholar] [CrossRef]
- García, E.E.; Belin, J.M.; Waché, Y. Use of a Doehlert factorial design to investigate the effects of pH and aeration on the accumulation of lactones by Yarrowia lipolytica. J. Appl. Microbiol. 2007, 103, 1508–1515. [Google Scholar] [CrossRef]
- Moradi, H.; Asadollahi, M.A.; Nahvi, I. Optimization of gamma-decalactone production by yeast Yarrowia lipolytica using the Taguchi method. J. Microbiol. Biotechnol. Food Sci. 2016, 6, 685–688. [Google Scholar] [CrossRef]
- Braga, A.; Belo, I. Production of γ-decalactone by Yarrowia lipolytica: Insights into experimental conditions and operating mode optimization. J. Chem. Technol. Biotechnol. 2015, 90, 559–565. [Google Scholar] [CrossRef]
- Darvishi, F.; Meidani, A.K. Optimization of gamma-decalactone production by Yarrowia lipolytica mutant strain via Response surface methodology. Modares J. Biotechnol. 2021, 12, 13–31. [Google Scholar]
- Pereira de Andrade, D.; Carvalho, B.F.; Schwan, R.F.; Dias, D.R. Production of γ-decalactone by yeast strains under different conditions. Food Technol. Biotechnol. 2017, 55, 225–230. [Google Scholar] [CrossRef]
- Braga, A.; Mesquita, D.P.; Amaral, A.L.; Ferreira, E.C.; Belo, I. Aroma production by Yarrowia lipolytica in airlift and stirred tank bioreactors: Differences in yeast metabolism and morphology. Biochem. Eng. J. 2015, 93, 55–62. [Google Scholar] [CrossRef]
- Puthli, M.S.; Rathod, V.K.; Pandit, A.B. Gaz-liquid mass transfer studies with triple impeller system on a laboratory scale bioreactor. Biochem. Eng. J. 2005, 23, 25–30. [Google Scholar] [CrossRef]
- Gomes, N.; Teixeira, J.A.; Belo, I. Empirical modelling as an experimental approach to optimize lactone production. Catal. Sci. Technol. 2011, 1, 86–92. [Google Scholar] [CrossRef][Green Version]
- Rahul, D.; Pretesh, J. Application of Taguchi-Based Design of Experiments for Industrial Chemical Processes. In Statistical Approaches with Emphasis on Design of Experiments Applied to Chemical Processes; IntechOpen: London, UK, 2018; Chapter 9. [Google Scholar]
- Rong, L.; Liu, S.; Zhu, K.; Kong, J.; Miao, L.; Wang, S.; Xiao, D.; Yu, A. Production of carboxylic acids by metabolically engineered Yarrowia lipolytica: A review. Sheng Wu Gong Cheng Xue Bao 2022, 38, 1360–1372. [Google Scholar] [CrossRef]
- Jach, M.E.; Malm, A. Yarrowia lipolytica as an alternative and valuable source of nutritional and bioactive compounds for humans. Molecules 2022, 27, 2300. [Google Scholar] [CrossRef]
- Fickers, P.; Marty, A.; Nicaud, J.M. The Lipases from Yarrowia lipolytica: Genetics, production, regulation, biochemical characterization and biotechnological applications. Biotechnol. Adv. 2011, 29, 632–664. [Google Scholar] [CrossRef]
- Colacicco, M.; Ciliberti, C.; Agrimi, G.; Biundo, A.; Pisano, I. Towards the physiological understanding of Yarrowia lipolytica growth and lipase production using waste cooking oils. Energies 2022, 15, 5217. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part II: Technology and potential applications. Eur. J. Lipid Sci. Technol. 2011, 113, 1052–1073. [Google Scholar] [CrossRef]
- Aguedo, M.; Waché, Y.; Coste, F.; Husson, F.; Belin, J.M. Impact of surfactants on the biotransformation of methyl ricinoleate into γ-decalactone by Yarrowia lipolytica. J. Mol. Catal. B Enzym. 2004, 29, 31–36. [Google Scholar] [CrossRef]
- Taguchi, G.; Konishi, S. Orthogonal Arrays and Linear Graphs; American Supplier Institute Inc.: Dearborn, MI, USA, 1987. [Google Scholar]
- Białecka-Florjańczyk, E.; Małajowicz, J.; Dudziak, B. Chemical and biotechnological synthesis of gamma-decalactone. Przem. Chem. 2016, 95, 999–1005. [Google Scholar] [CrossRef]
- Waché, Y.; Aguedo, M.; LeDall, M.T.; Nicaud, J.M.; Belin, J.M. Optimization of Yarrowia lipolytica’s β-oxidation pathway for γ-decalactone production. J. Mol. Catal. B Enzym. 2002, 19–20, 347–351. [Google Scholar] [CrossRef]
- Escamilla-García, E.; O’Riordana, S.; Gomes, N.; Aguedo, A.; Belo, I.; Teixeira, J.A.; Belina, J.M.; Wache, Y. An air-lift biofilm reactor for the production of γ-decalactones by Yarrowia lipolytica. Process Biochem. 2014, 49, 1377–1382. [Google Scholar] [CrossRef]
- Aguedo, M.; Gomes, N.; García, E.E.; Waché, Y.; Mota, M.; Teixeira, J.A.; Belo, I. Decalactone production by Yarrowia lipolytica under increased O2 transfer rates. Biotechnol. Lett. 2005, 27, 1617–1621. [Google Scholar] [CrossRef][Green Version]
- Gomes, N.; Aguedo, M.; Teixeira, J.A.; Belo, I. Oxygen mass transfer in a biphasic medium: Influence on the biotransformation of methyl ricinoleate into γ-decalactone by the yeast Yarrowia lipolytica. Biochem. Eng. J. 2007, 35, 380–386. [Google Scholar] [CrossRef]
- Gomes, N.; Teixeira, J.A.; Belo, I. Oxygen effect in γ-decalactone production through biotransformation of ricinoleic acid. In Proceedings of the 8th World Congress of Chemical Engineering, Montréal, QC, Canada, 23–27 August 2009. [Google Scholar]
- Le Do, T.T.; Vu, N.T.; Phan-Thi, H.; Cao-Hoang, L.; Ta, T.M.N.; Waché, Y.; Nguyen, T.H.T. Traditional fermented sausage ‘Nem chua’ as a source of yeast biocatalysts efficient for the production of the aroma compound γ-decalactone. Int. J. Food Sci. Technol. 2014, 49, 1099–1105. [Google Scholar] [CrossRef]
- Gomes, N.; Waché, Y.; Teixeira, J.A.; Belo, I. Oil-in-water emulsions characterization by laser granulometry and impact on γ-decalactone production in Yarrowia lipolytica. Biotechnol. Lett. 2011, 33, 1601–1606. [Google Scholar] [CrossRef][Green Version]
- Moradi, H.; Asadollahi, M.A.; Nahvi, I. Improved γ-decalactone production from castor oil by fed-batch cultivation of Yarrowia lipolytica. Biocatal. Agric. Biotechnol. 2013, 2, 64–68. [Google Scholar] [CrossRef]
- Gomes, N.; Teixeira, J.A.; Belo, I. The use of methyl ricinoleate in lactone production by Yarrowia lipolytica: Aspects of bioprocess operation that influence the overall performance. Biocatal. Biotransform. 2010, 28, 227–234. [Google Scholar] [CrossRef]
- Waché, Y.; Bergmark, K.; Courthaudon, J.L.; Aguedo, M.; Nicaud, J.M.; Belin, J.M. Medium-size droplets of methyl ricinoleate are reduced by cell-surface activity in the γ-decalactone production by Yarrowia lipolytica. Lett. Appl. Microbiol. 2000, 30, 183–187. [Google Scholar] [CrossRef][Green Version]
- Bouchedja, D.N.; Danthine, S.; Kar, T.; Fickers, P.; Sassi, H.; Boudjellal, A.; Blecker, C.; Delvigne, F. pH level has a strong impact on population dynamics of the yeast Yarrowia lipolytica and oil micro-droplets in multiphasic bioreactor. FEMS Microbiol. Lett. 2018, 365, fny173. [Google Scholar] [CrossRef]
- Timoumi, A.; Cléret, M.; Bideaux, C.; Guillouet, S.E.; Allouche, Y.; Molina-Jouve, C.; Fillaudeau, L.; Gorret, N. Dynamic behavior of Yarrowia lipolytica in response to pH perturbations: Dependence of the stress response on the culture mode. Appl. Microb. Cell Physiol. 2017, 101, 351–366. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Bioreactor Culture Number | |||
|---|---|---|---|---|
| 1 | 2 | 3 | 4 | |
| pH | Level 1 | Level 2 | Level 1 | Level 2 |
| Agitation speed | Level 1 | Level 1 | Level 2 | Level 2 |
| Substrate concentration | Level 1 | Level 2 | Level 2 | Level 1 |
| Level | pH Regulation | Agitation Speed (rpm) | Substrate Concentration (g/L) |
|---|---|---|---|
| 1 | Initially 7, without regulation | Variable (200–500) | 75 |
| 2 | Regulated | Constant (500) | 50 |
| Parameter | Unit | Bioreactor Batch Cultures | |||
|---|---|---|---|---|---|
| 1 * | 2 | 3 | 4 | ||
| Initial concentration of carbon source (S) | g/L | 75 | 50 | 50 | 75 |
| Time (t) | h | 37 (64) | 45 | 45 | 46 |
| Biomass yield (X) | gd.w./L | 24.43 ± 1.73 | 12.93 ± 1.31 | 7.17 ± 1.45 | 9.60 ± 0.95 |
| Maximum concentration of GDL (Lmax) | g/L | 1.56 ± 0.04 | 2.15 ± 0.04 | 0.19 ± 0.01 | 0.55 ± 0.01 |
| Conversion yield of biomass per carbon substrate (YX/S) | gd.w./g | 0.326 | 0.259 | 0.143 | 0.128 |
| Conversion yield of GDL per biomass formed (YL/X) | g/gd.w. | 0.064 | 0.166 | 0.026 | 0.057 |
| Conversion yield of GDL per carbon substrate (YL/S) | g/g | 0.021 | 0.043 | 0.004 | 0.007 |
| Volumetric rate of GDL production (qLv) | g/L/h | 0.042 (0.024) | 0.050 | 0.004 | 0.011 |
| Specific rate of GDL production (qL) | g/gd.w./h | 0.0017 (0.0010) | 0.0039 | 0.0006 | 0.0012 |
| pH | Mixing Intensity | Substrate Concentration |
|---|---|---|
| regulated | variable | 75 g/L |
| Parameter | Unit | Bioreactor Batch Culture |
|---|---|---|
| Initial concentration of carbon source (S) | g/L | 75 |
| Time (t) | h | 45 |
| Biomass yield (X) | gd.w./L | 16.35 ± 2.1 |
| Maximum concentration of GDL (Lmax) | g/L | 2.93 ± 0.33 |
| Conversion yield of GDL per biomass formed (YL/X) | g/gd.w. | 0.179 |
| Conversion yield of GDL per carbon substrate (YL/S) | g/g | 0.039 |
| Volumetric rate of GDL production (qLv) | g/L/h | 0.071 |
| Specific rate of GDL production (qL) | g/gd.w./h | 0.0044 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Małajowicz, J.; Fabiszewska, A.; Nowak, D.; Kuśmirek, S. Improved Gamma-Decalactone Synthesis by Yarrowia lipolytica Yeast Using Taguchi Robust Design Method. Appl. Sci. 2022, 12, 10231. https://doi.org/10.3390/app122010231
Małajowicz J, Fabiszewska A, Nowak D, Kuśmirek S. Improved Gamma-Decalactone Synthesis by Yarrowia lipolytica Yeast Using Taguchi Robust Design Method. Applied Sciences. 2022; 12(20):10231. https://doi.org/10.3390/app122010231
Chicago/Turabian StyleMałajowicz, Jolanta, Agata Fabiszewska, Dorota Nowak, and Sabina Kuśmirek. 2022. "Improved Gamma-Decalactone Synthesis by Yarrowia lipolytica Yeast Using Taguchi Robust Design Method" Applied Sciences 12, no. 20: 10231. https://doi.org/10.3390/app122010231
APA StyleMałajowicz, J., Fabiszewska, A., Nowak, D., & Kuśmirek, S. (2022). Improved Gamma-Decalactone Synthesis by Yarrowia lipolytica Yeast Using Taguchi Robust Design Method. Applied Sciences, 12(20), 10231. https://doi.org/10.3390/app122010231