

The Impact of Physical Activity on the Circadian System: Benefits for Health, Performance and Wellbeing



{kind=link}
Abstract
:Featured Application
Abstract
1. Introduction
2. Physical Activity and the Circadian System
2.1. The Diagnostic Value of Circadian Activity Rhythms
2.2. Beneficial Effects of Increasing the Daily Activity Level
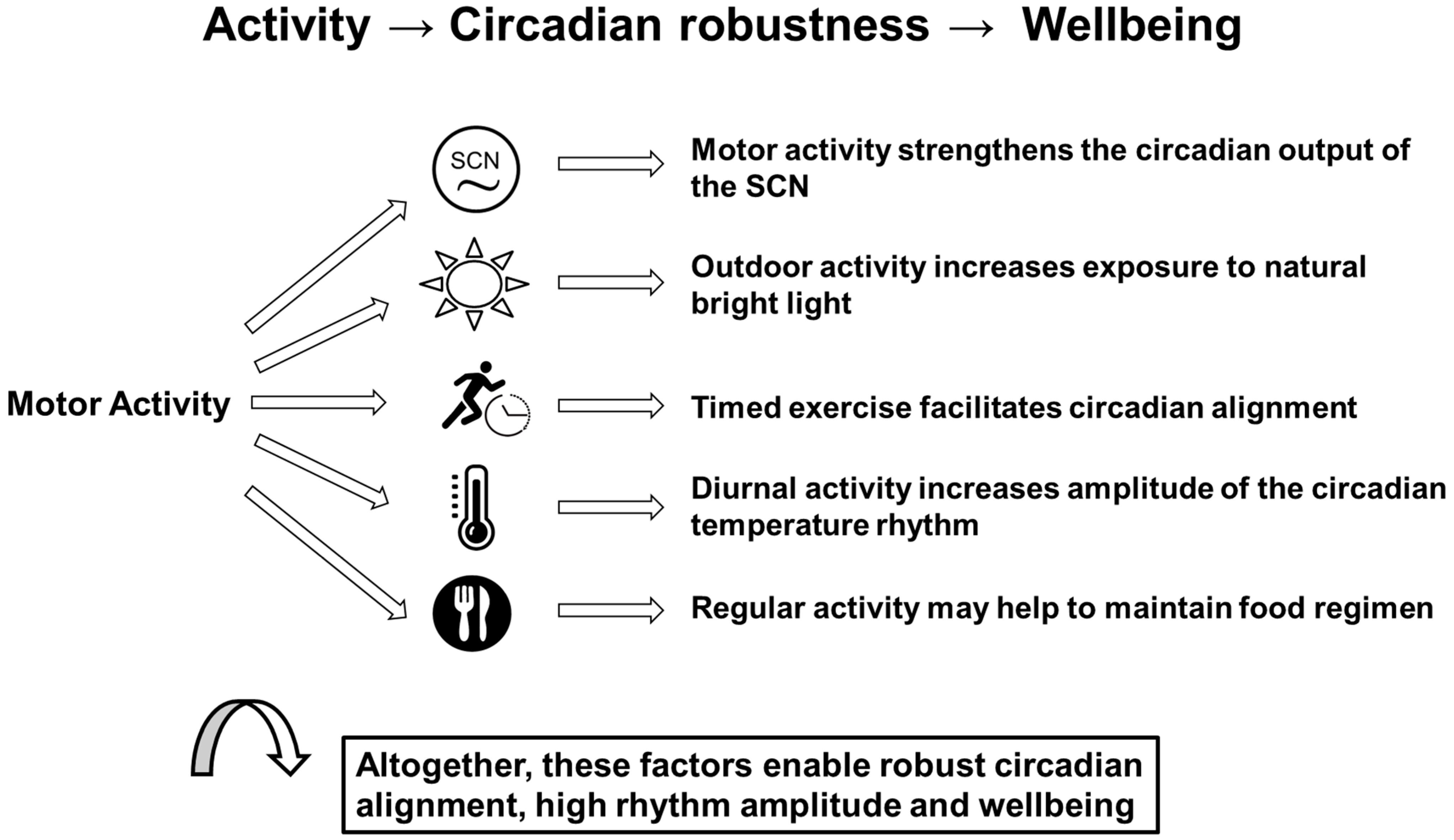
2.3. Effects of Motor Activity on the Circadian System
2.4. Beneficial Effects of Stabilized Circadian Rhythms
3. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef]
- Dijk, D.J.; Lockley, S.W. Integration of human sleep-wake regulation and circadian rhythmicity. J. Appl. Physiol. 2002, 92, 852–862. [Google Scholar] [CrossRef]
- Wollnik, F.; Turek, F.W. SCN lesions abolish ultradian and circadian components of activity rhythms in LEW/Ztm rats. Am. J. Physiol. 1989, 256, R1027–R1039. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Scheer, F.A.; Perreau-Lenz, S.; La Fleur, S.E.; Yi, C.X.; Fliers, E.; Buijs, R.M. Circadian disruption and SCN control of energy metabolism. FEBS Lett. 2011, 585, 1412–1426. [Google Scholar] [CrossRef]
- Waterhouse, J.; Drust, B.; Weinert, D.; Edwards, B.; Gregson, W.; Atkinson, G.; Kao, S.; Aizawa, S.; Reilly, T. The circadian rhythm of core temperature: Origin and some implications for exercise performance. Chronobiol. Int. 2005, 22, 207–225. [Google Scholar] [CrossRef]
- Hurd, M.W.; Ralph, M.R. The significance of circadian organization for longevity in the golden hamster. J. Biol. Rhythm. 1998, 13, 430–436. [Google Scholar] [CrossRef]
- Müller, L.; Weinert, D. Individual recognition of social rank and social memory performance depends on a functional circadian system. Behav Process. 2016, 132, 85–93. [Google Scholar] [CrossRef]
- Sharma, A.; Tiwari, S.; Singaravel, M. Circadian rhythm disruption: Health consequences. Biol. Rhythm Res. 2015, 47, 191–213. [Google Scholar] [CrossRef]
- Smolensky, M.H.; Hermida, R.C.; Reinberg, A.; Sackett-Lundeen, L.; Portaluppi, F. Circadian disruption: New clinical perspective of disease pathology and basis for chronotherapeutic intervention. Chronobiol. Int. 2016, 33, 1101–1119. [Google Scholar] [CrossRef]
- Weinert, D.; Schöttner, K.; Müller, L.; Wienke, A. Intensive voluntary wheel running may restore circadian activity rhythms and improves the impaired cognitive performance of arrhythmic Djungarian hamsters. Chronobiol. Int. 2016, 33, 1161–1170. [Google Scholar] [CrossRef]
- Gubin, D.G.; Weinert, D. Temporal order deterioration and circadian disruption with age. 1. Central and peripheral mechanisms. Adv. Gerontol. 2015, 5, 209–218. [Google Scholar] [CrossRef]
- Gubin, D.G.; Weinert, D. Deterioration of Temporal Order and Circadian Disruption with Age 2: Systemic Mechanisms of Aging Related Circadian Disruption and Approaches to Its Correction. Adv. Gerontol. 2016, 6, 10–20. [Google Scholar] [CrossRef]
- Gubin, D.G.; Weinert, D.; Bolotnova, T.V. Age-Dependent Changes of the Temporal Order—Causes and Treatment. Curr. Aging Sci. 2016, 9, 14–25. [Google Scholar] [CrossRef]
- Weinert, D. Age-dependent changes of the circadian system. Chronobiol. Int. 2000, 17, 261–283. [Google Scholar] [CrossRef]
- Casiraghi, L.P.; Oda, G.A.; Chiesa, J.J.; Friesen, W.O.; Golombek, D.A. Forced Desynchronization of Activity Rhythms in a Model of Chronic Jet Lag in Mice. J. Biol. Rhythm. 2012, 27, 59–69. [Google Scholar] [CrossRef]
- Waterhouse, J.; Kao, S.; Edwards, B.; Weinert, D.; Atkinson, G.; Reilly, T. Transient changes in the pattern of food intake following a simulated time-zone transition to the east across eight time zones. Chronobiol. Int. 2005, 22, 299–319. [Google Scholar] [CrossRef]
- Waterhouse, J.; Reilly, T.; Atkinson, G.; Edwards, B. Jet lag: Trends and coping strategies. Lancet 2007, 369, 1117–1129. [Google Scholar] [CrossRef]
- Shurkevich, N.P.; Vetoshkin, A.S.; Gapon, L.I.; Dyachkov, S.M.; Gubin, D.G. Prognostic value of blood pressure circadian rhythm disturbances in normotensive shift workers of the Arctic polar region. Arter. Hypertens. 2017, 23, 36–46. (In Russian) [Google Scholar]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between light at night, melatonin secretion, sleep deprivation, and the internal clock: Health impacts and mechanisms of circadian disruption. Life Sci. 2017, 173, 94–106. [Google Scholar] [CrossRef]
- Gubin, D.G.; Nelaeva, A.A.; Uzhakova, A.E.; Hasanova, Y.V.; Cornelissen, G.; Weinert, D. Disrupted circadian rhythms of body temperature, heart rate and fasting blood glucose in prediabetes and type 2 diabetes mellitus. Chronobiol. Int. 2017, 34, 1136–1148. [Google Scholar] [CrossRef]
- Kent, B.A. Synchronizing an aging brain: Can entraining circadian clocks by food slow Alzheimer’s disease? Front. Aging Neurosci. 2014, 6, 234. [Google Scholar] [CrossRef] [PubMed]
- Neroev, V.; Malishevskaya, T.; Weinert, D.; Astakhov, S.; Kolomeichuk, S.; Cornelissen, G.; Kabitskaya, Y.; Boiko, E.; Nemtsova, I.; Gubin, D. Disruption of 24-Hour Rhythm in Intraocular Pressure Correlates with Retinal Ganglion Cell Loss in Glaucoma. Int. J. Mol. Sci. 2021, 22, 359. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, A.; Kalsbeek, A.; Buijs, R.M. Functional changes of the SCN in spontaneous hypertension but not after the induction of hypertension. Chronobiol. Int. 2018, 35, 1221–1235. [Google Scholar] [CrossRef]
- Cardinali, D.P. Melatonin as a chronobiotic/cytoprotector: Its role in healthy aging. Biol. Rhythm Res. 2019, 50, 28–45. [Google Scholar] [CrossRef]
- Cardinali, D.P.; Brown, G.M.; Pandi-Perumal, S.R. Can Melatonin Be a Potential “Silver Bullet” in Treating COVID-19 Patients? Diseases 2020, 8, 44. [Google Scholar] [CrossRef]
- Hardeland, R. Divergent Importance of Chronobiological Considerations in High- and Low-dose Melatonin Therapies. Diseases 2021, 9, 18. [Google Scholar] [CrossRef]
- Lewy, A.J.; Ahmed, S.; Jackson, J.M.; Sack, R.L. Melatonin shifts human circadian rhythms according to a phase-response curve. Chronobiol. Int. 1992, 9, 380–392. [Google Scholar] [CrossRef]
- Gubin, D.G.; Gubin, G.D.; Gapon, L.I.; Weinert, D. Daily Melatonin Administration Attenuates Age-Dependent Disturbances of Cardiovascular Rhythms. Curr. Aging Sci. 2016, 9, 5–13. [Google Scholar] [CrossRef]
- Gubin, D.G.; Gubin, G.D.; Waterhouse, J.; Weinert, D. The circadian body temperature rhythm in the elderly: Effect of single daily melatonin dosing. Chronobiol. Int. 2006, 23, 639–658. [Google Scholar] [CrossRef]
- Daan, S. The Colin S. Pittendrigh Lecture. Colin Pittendrigh, Jurgen Aschoff, and the natural entrainment of circadian systems. J. Biol. Rhythm. 2000, 15, 195–207. [Google Scholar] [CrossRef]
- Golombek, D.A.; Rosenstein, R.E. Physiology of circadian entrainment. Physiol. Rev. 2010, 90, 1063–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.; Ryu, S.-H.; Lee, B.R.; Kim, K.H.; Lee, E.; Choi, J. Effects of artificial light at night on human health: A literature review of observational and experimental studies applied to exposure assessment. Chronobiol. Int. 2015, 32, 1294–1310. [Google Scholar] [CrossRef] [PubMed]
- Green, A.; Cohen-Zion, M.; Haim, A.; Dagan, Y. Evening light exposure to computer screens disrupts human sleep, biological rhythms, and attention abilities. Chronobiol. Int. 2017, 34, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Gubin, D.G.; Weinert, D.; Rybina, S.V.; Danilova, L.A.; Solovieva, S.V.; Durov, A.M.; Prokopiev, N.Y.; Ushakov, P.A. Activity, sleep and ambient light have a different impact on circadian blood pressure, heart rate and body temperature rhythms. Chronobiol. Int. 2017, 34, 632–649. [Google Scholar] [CrossRef] [PubMed]
- Smolensky, M.H.; Sackett-Lundeen, L.L.; Portaluppi, F. Nocturnal light pollution and underexposure to daytime sunlight: Complementary mechanisms of circadian disruption and related diseases. Chronobiol. Int. 2015, 32, 1029–1048. [Google Scholar] [CrossRef]
- Cajochen, C.; Freyburger, M.; Basishvili, T.; Garbazza, C.; Rudzik, F.; Renz, C.; Kobayashi, K.; Shirakawa, Y.; Stefani, O.; Weibel, J. Effect of daylight LED on visual comfort, melatonin, mood, waking performance and sleep. Light. Res. Technol. 2019, 51, 1044–1062. [Google Scholar] [CrossRef]
- Lowden, A.; Ozturk, G.; Reynolds, A.; Bjorvatn, B. Working Time Society consensus statements: Evidence based interventions using light to improve circadian adaptation to working hours. Ind. Health 2019, 57, 213–227. [Google Scholar] [CrossRef]
- Soltic, S.; Chalmers, A. Optimization of LED Lighting for Clinical Settings. J. Healthc. Eng. 2019, 2019, 5016013. [Google Scholar] [CrossRef]
- Castillo, M.R.; Hochstetler, K.J.; Tavernier, R.J., Jr.; Greene, D.M.; Bult-Ito, A. Entrainment of the master circadian clock by scheduled feeding. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2004, 287, R551–R555. [Google Scholar] [CrossRef]
- Froy, O.; Miskin, R. Effect of feeding regimens on circadian rhythms: Implications for aging and longevity. Aging 2010, 2, 7–27. [Google Scholar] [CrossRef]
- Lamont, E.W.; Diaz, L.R.; Barry-Shaw, J.; Stewart, J.; Amir, S. Daily restricted feeding rescues a rhythm of period2 expression in the arrhythmic suprachiasmatic nucleus. Neuroscience 2005, 132, 245–248. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokkan, K.A.; Yamazaki, S.; Tei, H.; Sakaki, Y.; Menaker, M. Entrainment of the circadian clock in the liver by feeding. Science 2001, 291, 490–493. [Google Scholar] [CrossRef]
- Acosta-Rodriguez, V.A.; Rijo-Ferreira, F.; Green, C.B.; Takahashi, J.S. Importance of circadian timing for aging and longevity. Nat. Commun. 2021, 12, 2862. [Google Scholar] [CrossRef] [PubMed]
- Mistlberger, R.E.; Antle, M.C. Entrainment of circadian clocks in mammals by arousal and food. Essays Biochem. 2011, 49, 119–136. [Google Scholar]
- Minors, D.; Atkinson, G.; Bent, N.; Rabbitt, P.; Waterhouse, J. The effects of age upon some aspects of lifestyle and implications for studies on circadian rhythmicity. Age Ageing 1998, 27, 67–72. [Google Scholar] [CrossRef]
- Monk, T.H.; Reynolds, C.F., III; Kupfer, D.J.; Hoch, C.C.; Carrier, J.; Houck, P.R. Differences over the life span in daily life-style regularity. Chronobiol. Int. 1997, 14, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Hallal, P.C.; Andersen, L.B.; Bull, F.C.; Guthold, R.; Haskell, W.; Ekelund, U. Lancet Physical Activity Series Working G. Global physical activity levels: Surveillance progress, pitfalls, and prospects. Lancet 2012, 380, 247–257. [Google Scholar] [CrossRef]
- Lee, I.M.; Shiroma, E.J.; Lobelo, F.; Puska, P.; Blair, S.N.; Katzmarzyk, P.T. Effect of physical inactivity on major non-communicable diseases worldwide: An analysis of burden of disease and life expectancy. Lancet 2012, 380, 219–229. [Google Scholar] [CrossRef]
- de Souza Teixeira, A.A.; Lira, F.S.; Rosa-Neto, J.C. Aging with rhythmicity. Is it possible? Physical exercise as a pacemaker. Life Sci. 2020, 261, 118453. [Google Scholar] [CrossRef]
- Weinert, D.; Waterhouse, J. The circadian rhythm of core temperature: Effects of physical activity and aging. Physiol. Behav. 2007, 90, 246–256. [Google Scholar] [CrossRef]
- Bellone, G.J.; Plano, S.A.; Cardinali, D.P.; Chada, D.P.; Vigo, D.E.; Golombek, D.A. Comparative analysis of actigraphy performance in healthy young subjects. Sleep Sci. 2016, 9, 272–279. [Google Scholar] [CrossRef]
- Ancoli-Israel, S.; Cole, R.; Alessi, C.; Chambers, M.; Moorcroft, W.; Pollak, C.P. The role of actigraphy in the study of sleep and circadian rhythms. Sleep 2003, 26, 342–392. [Google Scholar] [CrossRef] [Green Version]
- Carvalho-Bos, S.S.; Riemersma-van der Lek, R.F.; Waterhouse, J.; Reilly, T.; Van Someren, E.J. Strong association of the rest-activity rhythm with well-being in demented elderly women. Am. J. Geriatr. Psychiatry 2007, 15, 92–100. [Google Scholar] [CrossRef]
- Chevalier, V.; Mormont, M.C.; Cure, H.; Chollet, P. Assessment of circadian rhythms by actimetry in healthy subjects and patients with advanced colorectal cancer. Oncol. Rep. 2003, 10, 733–737. [Google Scholar] [PubMed]
- Weinert, D.; Sturm, J.; Waterhouse, J. Different behavior of the circadian rhythms of activity and body temperature during resynchronization following an advance of the LD cycle. Biol. Rhythm Res. 2002, 33, 187–197. [Google Scholar] [CrossRef]
- Weinert, D.; Weiss, T. A nonlinear interrelationship between period length and the amount of activity—Age-dependent changes. Biol. Rhythm Res. 1997, 28, 105–120. [Google Scholar] [CrossRef]
- Gubin, D.; Weinert, D.; Cornélissen, G. Chronotheranostics and chronotherapy—Frontiers for personalized medicine. J. Chronomed. 2020, 22, 3–23. [Google Scholar] [CrossRef]
- Martinez-Nicolas, A.; Madrid, J.A.; García, F.J.; Campos, M.; Moreno-Casbas, M.T.; Almaida-Pagán, P.F.; Lucas-Sánchez, A.; Rol, M.A. Circadian monitoring as an aging predictor. Sci. Rep. 2018, 8, 15027. [Google Scholar] [CrossRef]
- Vitale, J.A.; Banfi, G.; Tivolesi, V.; Pelosi, C.; Borghi, S.; Negrini, F. Rest-activity daily rhythm and physical activity levels after hip and knee joint replacement: The role of actigraphy in orthopedic clinical practice. Chronobiol. Int. 2021, 38, 1692–1701. [Google Scholar] [CrossRef]
- Minors, D.; Akerstedt, T.; Atkinson, G.; Dahlitz, M.; Folkard, S.; Levi, F.; Mormont, C.; Parkes, D.; Waterhouse, J. The difference between activity when in bed and out of bed. I. Healthy subjects and selected patients. Chronobiol. Int. 1996, 13, 27–34. [Google Scholar] [CrossRef]
- Mormont, M.C.; Langouet, A.M.; Claustrat, B.; Bogdan, A.; Marion, S.; Waterhouse, J.; Touitou, Y.; Levi, F. Marker rhythms of circadian system function: A study of patients with metastatic colorectal cancer and good performance status. Chronobiol. Int. 2002, 19, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Mormont, M.C.; Waterhouse, J. Contribution of the rest-activity circadian rhythm to quality of life in cancer patients. Chronobiol. Int. 2002, 19, 313–323. [Google Scholar] [CrossRef]
- Mormont, M.C.; Waterhouse, J.; Bleuzen, P.; Giacchetti, S.; Jami, A.; Bogdan, A.; Lellouch, J.; Misset, J.L.; Touitou, Y.; Levi, F. Marked 24-h rest/activity rhythms are associated with better quality of life, better response, and longer survival in patients with metastatic colorectal cancer and good performance status. Clin. Cancer Res. 2000, 6, 3038–3045. [Google Scholar]
- Hoopes, E.K.; Witman, M.A.; D’Agata, M.N.; Berube, F.R.; Brewer, B.; Malone, S.K.; Grandner, M.A.; Patterson, F. Rest-activity rhythms in emerging adults: Implications for cardiometabolic health. Chronobiol. Int. 2021, 38, 543–556. [Google Scholar] [CrossRef]
- Huang, T.; Mariani, S.; Redline, S. Sleep Irregularity and Risk of Cardiovascular Events. J. Am. Coll. Cardiol. 2020, 75, 991–999. [Google Scholar] [CrossRef]
- Borisenkov, M.; Tserne, T.; Bakutova, L.; Gubin, D. Food Addiction and Emotional Eating Are Associated with Intradaily Rest–Activity Rhythm Variability. Eat Weight Disord. 2022. [Google Scholar] [CrossRef]
- Gubin, D.G.; Malishevskaya, T.N.; Astakhov, Y.S.; Astakhov, S.Y.; Cornelissen, G.; Kuznetsov, V.A.; Weinert, D. Progressive retinal ganglion cell loss in primary open-angle glaucoma is associated with temperature circadian rhythm phase delay and compromised sleep. Chronobiol. Int. 2019, 36, 564–577. [Google Scholar] [CrossRef]
- Schöttner, K.; Limbach, A.; Weinert, D. Re-entrainment behavior of Djungarian hamsters (Phodopus sungorus) with different rhythmic phenotype following light-dark shifts. Chronobiol. Int. 2011, 28, 58–69. [Google Scholar] [CrossRef]
- Weinert, H.; Weinert, D. Circadian activity rhythms of laboratory mice during the last weeks of their life. Biol. Rhythm Res. 1998, 29, 159–178. [Google Scholar] [CrossRef]
- Weinert, H.; Weinert, D.; Waterhouse, J. The circadian activity and body temperature rhythms of mice during their last days of life. Biol. Rhythm Res. 2002, 33, 199–212. [Google Scholar] [CrossRef]
- Sturm, J.; Weinert, H.; Weinert, D. Age-dependent changes in the stability of the daily activity rythms of laboratory mice. Z. Saugetierkd. Int. J. Mamm. Biol. 2000, 65, 21–32. [Google Scholar]
- Weinert, H.; Weinert, D.; Schurov, I.; Maywood, E.S.; Hastings, M.H. Impaired expression of the mPer2 circadian clock gene in the suprachiasmatic nuclei of aging mice. Chronobiol. Int. 2001, 18, 559–565. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Gao, L.; Gaba, A.; Yu, L.; Cui, L.; Fan, W.; Lim, A.S.P.; Bennett, D.A.; Buchman, A.S.; Hu, K. Circadian disturbances in Alzheimer’s disease progression: A prospective observational cohort study of community-based older adults. Lancet Healthy Longev. 2020, 1, e96–e105. [Google Scholar] [CrossRef]
- Werth, E.; Savaskan, E.; Knoblauch, V.; Gasio, P.F.; van Someren, E.J.; Hock, C.; Wirz-Justice, A. Decline in long-term circadian rest-activity cycle organization in a patient with dementia. J. Geriatr. Psychiatry Neurol. 2002, 15, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Zampogna, A.; Manoni, A.; Asci, F.; Liguori, C.; Irrera, F.; Suppa, A. Shedding Light on Nocturnal Movements in Parkinson’s Disease: Evidence from Wearable Technologies. Sensors 2020, 20, 5171. [Google Scholar] [CrossRef] [PubMed]
- Targa, A.D.S.; Benítez, I.D.; Dakterzada, F.; Fontenele-Araujo, J.; Minguez, O.; Zetterberg, H.; Blennow, K.; Barbé, F.; Piñol-Ripoll, G. The circadian rest-activity pattern predicts cognitive decline among mild-moderate Alzheimer’s disease patients. Alzheimers Res. Ther. 2021, 13, 161. [Google Scholar] [CrossRef]
- Charansonney, O.L. Physical activity and aging: A life-long story. Discov. Med. 2011, 12, 177–185. [Google Scholar]
- Choi, Y.; Cho, J.; No, M.-H.; Heo, J.-W.; Cho, E.-J.; Chang, E.; Park, D.-H.; Kang, J.-H.; Kwak, H.-B. Re-Setting the Circadian Clock Using Exercise against Sarcopenia. Int. J. Mol. Sci. 2020, 21, 3106. [Google Scholar] [CrossRef]
- Kume, Y.; Kodama, A.; Maekawa, H. Preliminary report; Comparison of the circadian rest-activity rhythm of elderly Japanese community-dwellers according to sarcopenia status. Chronobiol. Int. 2020, 37, 1099–1105. [Google Scholar] [CrossRef]
- Balbus, J.M.; Barouki, R.; Birnbaum, L.S.; Etzel, R.A.; Gluckman, P.D.; Grandjean, P.; Hancock, C.; Hanson, M.A.; Heindel, J.J.; Hoffman, K.; et al. Early-life prevention of non-communicable diseases. Lancet 2013, 38, 3–4. [Google Scholar] [CrossRef]
- Kelly, S.A.; Pomp, D. Genetic determinants of voluntary exercise. Trends Genet. 2013, 29, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Stensel, D. Primary prevention of CVD: Physical activity. BMJ Clin. Evid. 2009, 2009, 0218. [Google Scholar] [PubMed]
- Gallanagh, S.; Quinn, T.J.; Alexander, J.; Walters, M.R. Physical activity in the prevention and treatment of stroke. ISRN Neurol. 2011, 2011, 953818. [Google Scholar] [CrossRef] [PubMed]
- Denlinger, C.S.; Engstrom, P.F. Colorectal cancer survivorship: Movement matters. Cancer Prev. Res. 2011, 4, 502–511. [Google Scholar] [CrossRef]
- Wadden, T.A.; Webb, V.L.; Moran, C.H.; Bailer, B.A. Lifestyle modification for obesity: New developments in diet, physical activity, and behavior therapy. Circulation 2012, 125, 1157–1170. [Google Scholar] [CrossRef] [Green Version]
- Matura, S.; Carvalho, A.F.; Alves, G.S.; Pantel, J. Physical Exercise for the Treatment of Neuropsychiatric Disturbances in Alzheimer’s Dementia: Possible Mechanisms, Current Evidence and Future Directions. Curr. Alzheimer Res. 2016, 13, 1112–1123. [Google Scholar] [CrossRef]
- Vyazovskiy, V.V.; Ruijgrok, G.; Deboer, T.; Tobler, I. Running wheel accessibility affects the regional electroencephalogram during sleep in mice. Cereb. Cortex. 2006, 16, 328–336. [Google Scholar] [CrossRef]
- Greenwood, B.N.; Fleshner, M. Exercise, learned helplessness, and the stress-resistant brain. Neuromol. Med. 2008, 10, 81–98. [Google Scholar] [CrossRef]
- Kingston, R.C.; Smith, M.; Lacey, T.; Edwards, M.; Best, J.N.; Markham, C.M. Voluntary exercise increases resilience to social defeat stress in Syrian hamsters. Physiol. Behav. 2018, 188, 194–198. [Google Scholar] [CrossRef]
- Groot, C.; Hooghiemstra, A.M.; Raijmakers, P.G.H.M.; van Berckel, B.N.M.; Scheltens, P.; Scherder, E.J.A.; van der Flier, W.M.; Ossenkoppele, R. The effect of physical activity on cognitive function in patients with dementia: A meta-analysis of randomized control trials. Ageing Res. Rev. 2016, 25, 13–23. [Google Scholar] [CrossRef]
- Winchester, J.; Dick, M.B.; Gillen, D.; Reed, B.; Miller, B.; Tinklenberg, J.; Mungas, D.; Chui, H.; Galasko, D.; Hewett, L.; et al. Walking stabilizes cognitive functioning in Alzheimer’s disease (AD) across one year. Arch. Gerontol. Geriatr. 2013, 56, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Fabel, K.; Kempermann, G. Physical activity and the regulation of neurogenesis in the adult and aging brain. Neuromol. Med. 2008, 10, 59–66. [Google Scholar] [CrossRef]
- Feter, N.; Spanevello, R.M.; Soares, M.S.P.; Spohr, L.; Pedra, N.S.; Bona, N.P.; Freitas, M.P.; Gonzales, N.G.; Ito, L.G.M.S.; Stefanello, F.M.; et al. How does physical activity and different models of exercise training affect oxidative parameters and memory? Physiol. Behav. 2019, 201, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Mustroph, M.L.; Chen, S.; Desai, S.C.; Cay, E.B.; DeYoung, E.K.; Rhodes, J.S. Aerobic exercise is the critical variable in an enriched environment that increases hippocampal neurogenesis and water maze learning in male C57BL/6J mice. Neuroscience 2012, 219, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Rajizadeh, M.A.; Esmaeilpour, K.; Masoumi-Ardakani, Y.; Bejeshk, M.A.; Shabani, M.; Nakhaee, N.; Ranjbar, M.P.; Borzadaran, F.M.; Sheibani, V. Voluntary exercise impact on cognitive impairments in sleep-deprived intact female rats. Physiol. Behav. 2018, 188, 58–66. [Google Scholar] [CrossRef]
- van Praag, H.; Christie, B.R.; Sejnowski, T.J.; Gage, F.H. Running enhances neurogenesis, learning, and long-term potentiation in mice. Proc. Natl. Acad. Sci. USA 1999, 96, 13427–13431. [Google Scholar] [CrossRef]
- Edgar, D.M.; Kilduff, T.S.; Martin, C.E.; Dement, W.C. Influence of running wheel activity on free-running sleep/wake and drinking circadian rhythms in mice. Physiol. Behav. 1991, 50, 373–378. [Google Scholar] [CrossRef]
- Weinert, D.; Schöttner, K. An inbred lineage of Djungarian hamsters with a strongly attenuated ability to synchronize. Chronobiol. Int. 2007, 24, 1065–1079. [Google Scholar] [CrossRef]
- Antle, M.C.; Sterniczuk, R.; Smith, V.M.; Hagel, K. Non-photic modulation of phase shifts to long light pulses. J. Biol. Rhythm. 2007, 22, 524–533. [Google Scholar] [CrossRef]
- Ralph, M.R.; Mrosovsky, N. Behavioral inhibition of circadian responses to light. J. Biol. Rhythm. 1992, 7, 353–359. [Google Scholar] [CrossRef]
- Steinlechner, S.; Stieglitz, A.; Ruf, T. Djungarian hamsters: A species with a labile circadian pacemaker? Arrhythmicity under a light-dark cycle induced by short light pulses. J. Biol. Rhythm. 2002, 17, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Mrosovsky, N.; Salmon, P.A.; Menaker, M.; Ralph, M.R. Nonphotic phase shifting in hamster clock mutants. J. Biol. Rhythm. 1992, 7, 41–49. [Google Scholar] [CrossRef]
- Youngstedt, S.D.; Elliott, J.A.; Kripke, D.F. Human circadian phase-response curves for exercise. J. Physiol. 2019, 597, 2253–2268. [Google Scholar] [CrossRef] [PubMed]
- Deboer, T.; Vansteensel, M.J.; Detari, L.; Meijer, J.H. Sleep states alter activity of suprachiasmatic nucleus neurons. Nat. Neurosci. 2003, 6, 1086–1090. [Google Scholar] [CrossRef]
- Maywood, E.S.; Okamura, H.; Hastings, M.H. Opposing actions of neuropeptide Y and light on the expression of circadian clock genes in the mouse suprachiasmatic nuclei. Eur. J. Neurosci. 2002, 15, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Schaap, J.; Meijer, J.H. Opposing effects of behavioural activity and light on neurons of the suprachiasmatic nucleus. Eur. J. Neurosci. 2001, 13, 1955–1962. [Google Scholar] [CrossRef]
- Song, Y.; Choi, G.; Jang, L.; Kim, S.-W.; Jung, K.-H.; Park, H. Circadian rhythm gene expression and daily melatonin levels vary in athletes and sedentary males. Biol. Rhythm Res. 2018, 49, 237–245. [Google Scholar] [CrossRef]
- Ortega, G.J.; Romanelli, L.; Golombek, D.A. Statistical and dynamical analysis of circadian rhythms. J. Theor. Biol. 1994, 169, 15–21. [Google Scholar] [CrossRef]
- Leise, T.L.; Harrington, M.E.; Molyneux, P.C.; Song, I.; Queenan, H.; Zimmerman, E.; Lall, G.S.; Biello, S.M. Voluntary exercise can strengthen the circadian system in aged mice. Age 2013, 35, 2137–2152. [Google Scholar] [CrossRef]
- Weinert, D.; Schottner, K.; Meinecke, A.C.; Hauer, J. Voluntary exercise stabilizes photic entrainment of djungarian hamsters (Phodopus sungorus) with a delayed activity onset. Chronobiol. Int. 2018, 35, 1435–1444. [Google Scholar] [CrossRef]
- Meijer, J.H.; Robbers, Y. Wheel running in the wild. Proc. R. Soc. B Biol. Sci. 2014, 281, 20140210. [Google Scholar] [CrossRef] [PubMed]
- Mrosovsky, N.; Salmon, P.A.; Vrang, N. Revolutionary science: An improved running wheel for hamsters. Chronobiol. Int. 1998, 15, 147–158. [Google Scholar] [CrossRef] [PubMed]
- Farajnia, S.; Michel, S.; Deboer, T.; van der Leest, H.T.; Houben, T.; Rohling, J.H.; Ramkisoensing, A.; Yasenkov, R.; Meijer, J.H. Evidence for neuronal desynchrony in the aged suprachiasmatic nucleus clock. J. Neurosci. 2012, 32, 5891–5899. [Google Scholar] [CrossRef] [PubMed]
- Margraf, R.R.; Puchalski, W.; Lynch., G.R. Absence of a daily neuronal rhythm in the suprachiasmatic nuclei of acircadian Djungarian hamsters. Neurosci. Lett. 1992, 142, 175–178. [Google Scholar] [CrossRef]
- Yamazaki, S.; Kerbeshian, M.C.; Hocker, C.G.; Block, G.D.; Menaker, M. Rhythmic properties of the hamster suprachiasmatic nucleus in vivo. J. Neurosci. 1998, 18, 10709–10723. [Google Scholar] [CrossRef] [Green Version]
- van Oosterhout, F.; Lucassen, E.A.; Houben, T.; van der Leest, H.T.; Antle, M.C.; Meijer, J.H. Amplitude of the SCN clock enhanced by the behavioral activity rhythm. PLoS ONE 2012, 7, e39693. [Google Scholar]
- Power, A.; Hughes, A.T.; Samuels, R.E.; Piggins, H.D. Rhythm-promoting actions of exercise in mice with deficient neuropeptide signaling. J. Biol. Rhythm. 2010, 25, 235–246. [Google Scholar] [CrossRef]
- Mrosovsky, N.; Biello, S.M. Nonphotic phase shifting in the old and the cold. Chronobiol. Int. 1994, 11, 232–252. [Google Scholar] [CrossRef]
- Hower, I.M.; Harper, S.A.; Buford, T.W. Circadian Rhythms, Exercise, and Cardiovascular Health. J. Circadian Rhythm. 2018, 16, 7. [Google Scholar] [CrossRef]
- Rubio-Sastre, P.; Gómez-Abellán, P.; Martinez-Nicolas, A.; Ordovás, J.M.; Madrid, J.A.; Garaulet, M. Evening physical activity alters wrist temperature circadian rhythmicity. Chronobiol. Int. 2014, 31, 276–282. [Google Scholar] [CrossRef]
- Duglan, D.; Lamia, K.A. Clocking In, Working Out: Circadian Regulation of Exercise Physiology. Trends Endocrinol. Metab. 2019, 30, 347–356. [Google Scholar] [CrossRef] [PubMed]
- Thomas, J.M.; Kern, P.A.; Bush, H.M.; McQuerry, K.J.; Black, W.S.; Clasey, J.L.; Pendergast, J.S. Circadian rhythm phase shifts caused by timed exercise vary with chronotype. JCI Insight 2020, 5, e134270. [Google Scholar] [CrossRef] [PubMed]
- Heden, T.D.; Kanaley, J.A. Syncing Exercise with Meals and Circadian Clocks. Exerc. Sport Sci. Rev. 2019, 47, 22–28. [Google Scholar] [CrossRef] [PubMed]
- Lewis, P.; Korf, H.W.; Kuffer, L.; Groß, J.V.; Erren, T.C. Exercise time cues (zeitgebers) for human circadian systems can foster health and improve performance: A systematic review. BMJ Open Sport Exerc. Med. 2018, 4, e000443. [Google Scholar] [CrossRef] [PubMed]
- Beyer, K.M.M.; Szabo, A.; Hoormann, K.; Stolley, M. Time spent outdoors, activity levels, and chronic disease among American adults. J. Behav. Med. 2018, 41, 494–503. [Google Scholar] [CrossRef]
- Korman, M.; Tkachev, V.; Reis, C.; Komada, Y.; Kitamura, S.; Gubin, D.; Kumar, V.; Roenneberg, T. Outdoor daylight exposure and longer sleep promote wellbeing under COVID-19 mandated restrictions. J. Sleep Res. 2022, 31, e13471. [Google Scholar] [CrossRef]
- Takahashi, M.; Haraguchi, A.; Tahara, Y.; Aoki, N.; Fukazawa, M.; Tanisawa, K.; Ito, T.; Nakaoka, T.; Higuchi, M.; Shibata, S. Positive association between physical activity and PER3 expression in older adults. Sci. Rep. 2017, 7, 39771. [Google Scholar] [CrossRef]
- Dupont Rocher, S.; Bessot, N.; Sesboüé, B.; Bulla, J.; Davenne, D. Circadian Characteristics of Older Adults and Aerobic Capacity. J. Gerontol. A Biol. Sci. Med. Sci. 2016, 71, 817–822. [Google Scholar] [CrossRef]
- Dallmann, R.; Mrosovsky, N. Scheduled wheel access during daytime: A method for studying conflicting zeitgebers. Physiol. Behav. 2006, 88, 459–465. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Honma, S.; Honma, K.-I. Two Coupled Circadian Oscillators Are Involved in Nonphotic Acceleration of Reentrainment to Shifted Light Cycles in Mice. J. Biol. Rhythm. 2018, 33, 614–625. [Google Scholar] [CrossRef]
- Barger, L.K.; Kenneth, P.; Wright, J.; Hughes, R.J.; Czeisler, C.A. Daily exercise facilitates phase delays of circadian melatonin rhythm in very dim light. Am. J. Physiol. -Regul. Integr. Comp. Physiol. 2004, 286, R1077–R1084. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, Y.; Honma, K.-i.; Hashimoto, S.; Takasu, N.; Miyazaki, T.; Honma, S. Effects of physical exercise on human circadian rhythms. Sleep Biol. Rhythm. 2006, 4, 199–206. [Google Scholar] [CrossRef]
- Vitale, J.A.; Lombardi, G.; Weydahl, A.; Banfi, G. Biological rhythms, chronodisruption and chrono-enhancement: The role of physical activity as synchronizer in correcting steroids circadian rhythm in metabolic dysfunctions and cancer. Chronobiol. Int. 2018, 35, 1185–1197. [Google Scholar] [CrossRef] [PubMed]
- Smarr, B.L.; Jennings, K.J.; Driscoll, J.R.; Kriegsfeld, L.J. A time to remember: The role of circadian clocks in learning and memory. Behav. Neurosci. 2014, 128, 283–303. [Google Scholar] [CrossRef]
- Dijk, D.J.; Neri, D.F.; Wyatt, J.K.; Ronda, J.M.; Riel, E.; Ritz-De Cecco, A.; Hughes, R.J.; Elliott, A.R.; Prisk, G.K.; West, J.B.; et al. Sleep, performance, circadian rhythms, and light-dark cycles during two space shuttle flights. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2001, 281, R1647–R1664. [Google Scholar] [CrossRef] [PubMed]
- Diekelmann, S.; Born, J. The memory function of sleep. Nat. Rev. Neurosci. 2010, 11, 114–126. [Google Scholar] [CrossRef]
- Walker, M.P. The role of slow wave sleep in memory processing. J. Clin. Sleep Med. 2009, 5, S20–S26. [Google Scholar] [CrossRef]
- Walker, M.P.; Stickgold, R. Sleep-dependent learning and memory consolidation. Neuron 2004, 44, 121–133. [Google Scholar] [CrossRef]
- Cain, S.W.; Chou, T.; Ralph, M.R. Circadian modulation of performance on an aversion-based place learning task in hamsters. Behav. Brain Res. 2004, 150, 201–205. [Google Scholar] [CrossRef]
- Catala, M.D.; Pallardo, F.; Roman, A.; Villanueva, P.; Vina Giner, J.M. Effect of pinealectomy and circadian rhythm on avoidance behavior in the male rat. Physiol. Behav. 1985, 34, 327–333. [Google Scholar] [CrossRef]
- Müller, L.; Fritzsche, P.; Weinert, D. Novel object recognition of Djungarian hamsters depends on circadian time and rhythmic phenotype. Chronobiol. Int. 2015, 32, 458–467. [Google Scholar] [CrossRef]
- Devan, B.D.; Goad, E.H.; Petri, H.L.; Antoniadis, E.A.; Hong, N.S.; Ko, C.H.; Leblanc, L.; Lebovic, S.S.; Lo, Q.N.; Ralph, M.R.; et al. Circadian phase-shifted rats show normal acquisition but impaired long-term retention of place information in the water task. Neurobiol. Learn. Mem. 2001, 75, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Ruby, N.F.; Fernandez, F.; Garrett, A.; Klima, J.; Zhang, P.; Sapolsky, R.; Heller, H.C. Spatial memory and long-term object recognition are impaired by circadian arrhythmia and restored by the GABAA antagonist pentylenetetrazole. PLoS ONE 2013, 8, e72433. [Google Scholar] [CrossRef]
- Ruby, N.F.; Hwang, C.E.; Wessells, C.; Fernandez, F.; Zhang, P.; Sapolsky, R.; Heller, H.C. Hippocampal-dependent learning requires a functional circadian system. Proc. Natl. Acad. Sci. USA 2008, 105, 15593–15598. [Google Scholar] [CrossRef] [PubMed]
- Ruby, N.F.; Patton, D.F.; Bane, S.; Looi, D.; Heller, H.C. Reentrainment Impairs Spatial Working Memory until Both Activity Onset and Offset Reentrain. J. Biol. Rhythm. 2015, 30, 408–416. [Google Scholar] [CrossRef]
- Ruby, N.F.; Barakat, M.T.; Heller, H.C. Phenotypic differences in reentrainment behavior and sensitivity to nighttime light pulses in siberian hamsters. J. Biol. Rhythm. 2004, 19, 530–541. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weinert, D.; Gubin, D. The Impact of Physical Activity on the Circadian System: Benefits for Health, Performance and Wellbeing. Appl. Sci. 2022, 12, 9220. https://doi.org/10.3390/app12189220
Weinert D, Gubin D. The Impact of Physical Activity on the Circadian System: Benefits for Health, Performance and Wellbeing. Applied Sciences. 2022; 12(18):9220. https://doi.org/10.3390/app12189220
Chicago/Turabian StyleWeinert, Dietmar, and Denis Gubin. 2022. "The Impact of Physical Activity on the Circadian System: Benefits for Health, Performance and Wellbeing" Applied Sciences 12, no. 18: 9220. https://doi.org/10.3390/app12189220
APA StyleWeinert, D., & Gubin, D. (2022). The Impact of Physical Activity on the Circadian System: Benefits for Health, Performance and Wellbeing. Applied Sciences, 12(18), 9220. https://doi.org/10.3390/app12189220