
Effect of Spiny-Cheek Crayfish (Faxonius limosus) on H2O2-Induced Oxidative Stress in Normal Fibroblast Cells

 ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Extract Preparation
2.3. Extract Characterisation
2.3.1. Bioactive Compounds
2.3.2. Antioxidative Activity In Vitro
2.4. Cell Study
2.4.1. Cell Line
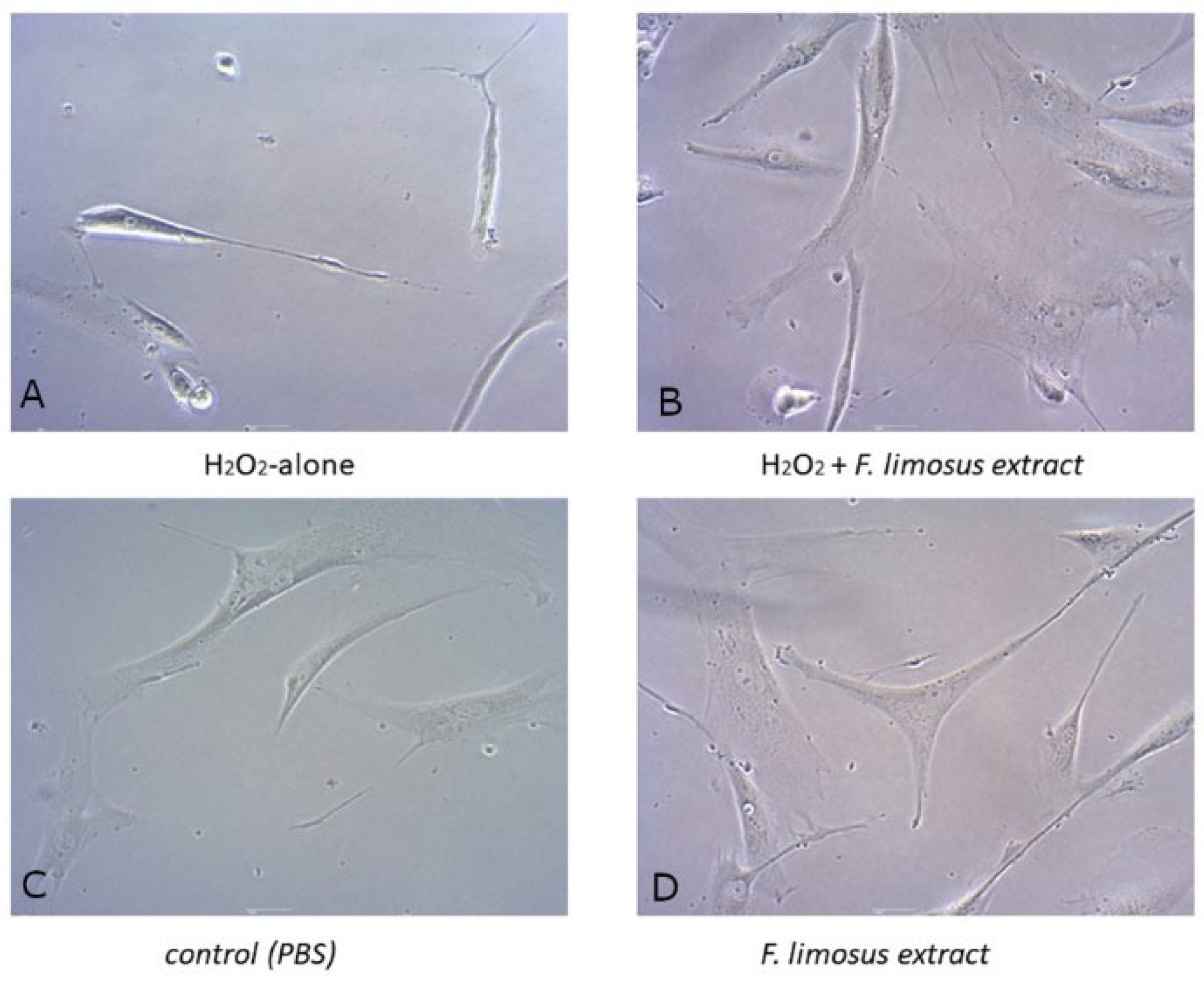
2.4.2. Light Microscopy
- PBS only control, which received neither extract nor hydrogen peroxide;
- H2O2 only control, which received no extract but had added H2O2;
- 100 µg/mL extract, which received the extract without H2O2;
- 100 µg/mL extract, which received the extract and H2O2.
2.4.3. Cytotoxicity Test
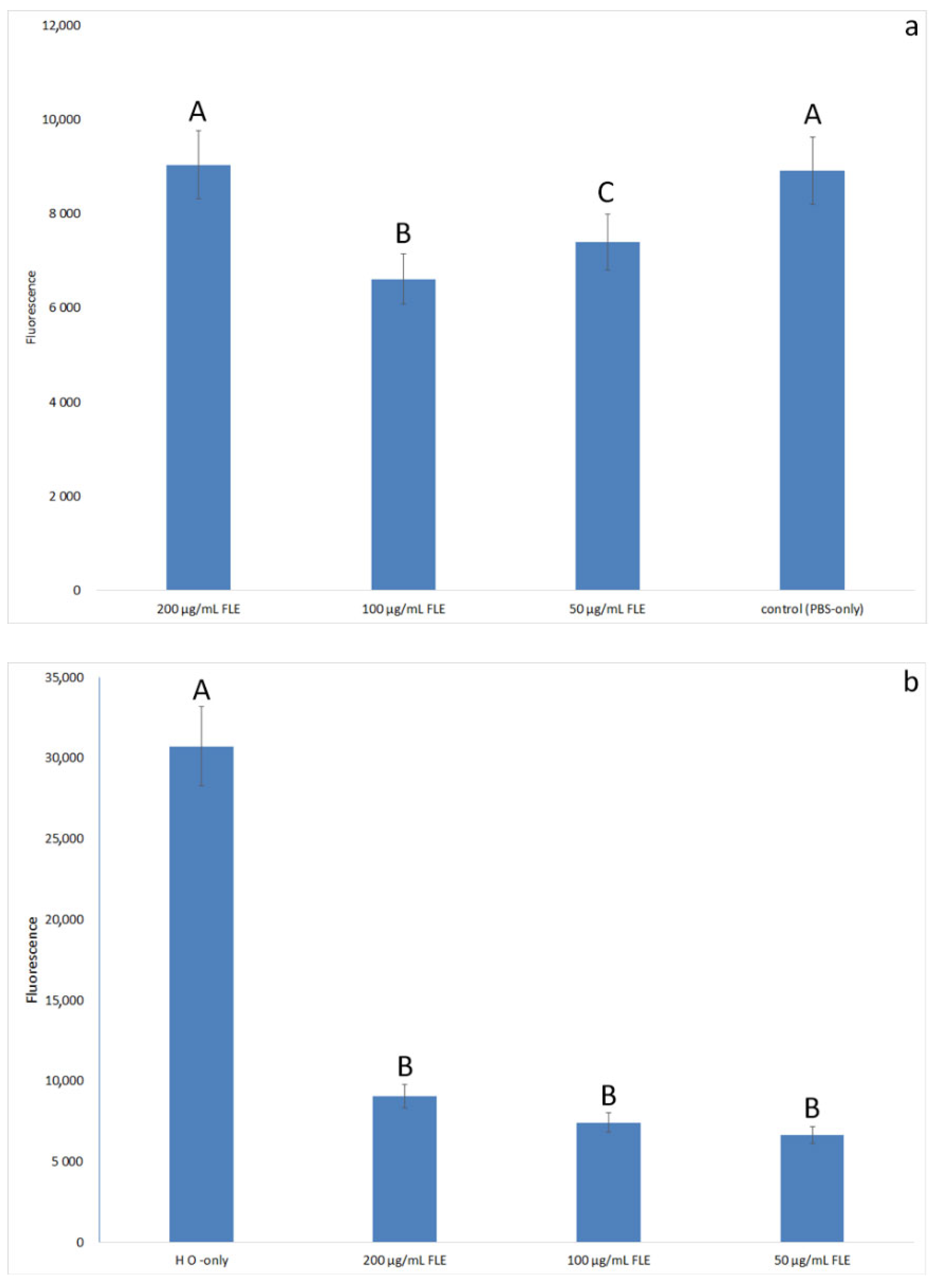
2.4.4. ROS Secretion Test
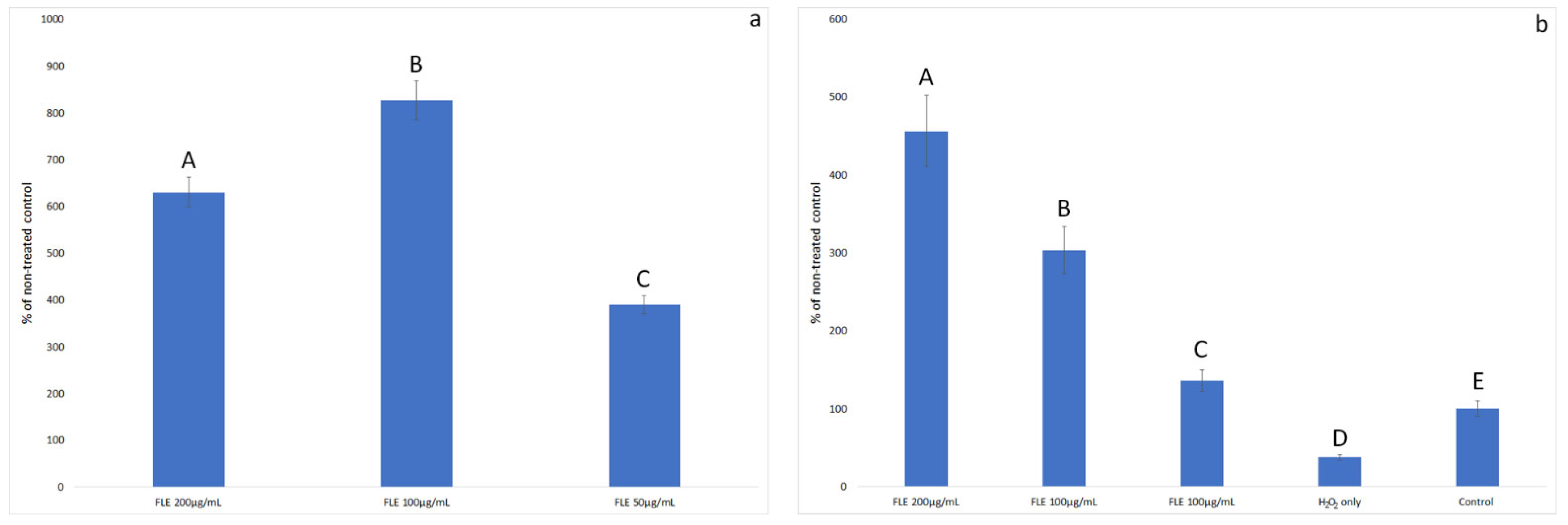
2.4.5. Proliferation Test
2.4.6. Caspase-3 Test
2.5. Statistics
3. Results
3.1. F. limosus Crayfish Extract Is a Source of Valuable Bioactive Substances
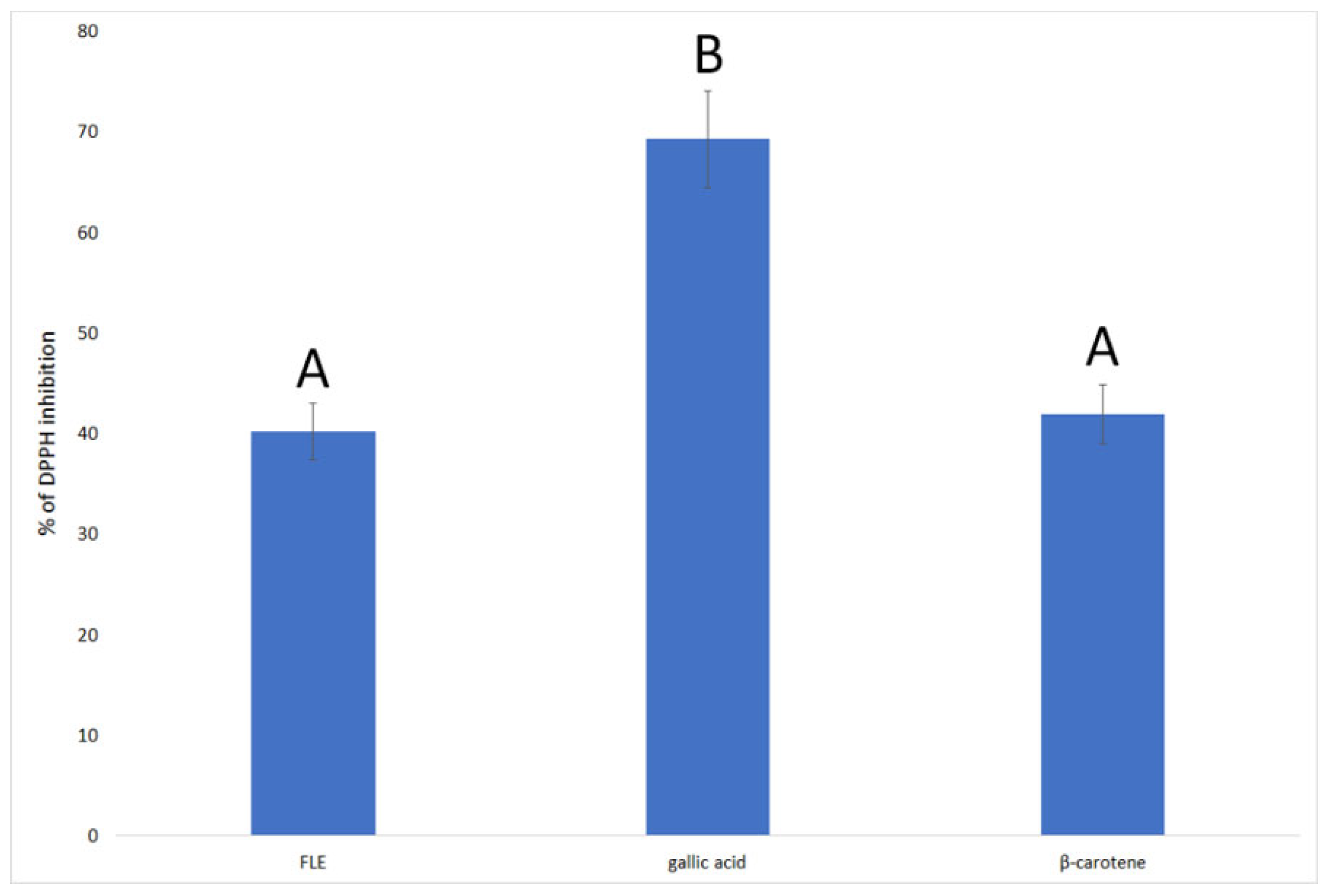
3.2. F. limosus Crayfish Extract Has Antioxidant Properties Comparable to β-Carotene
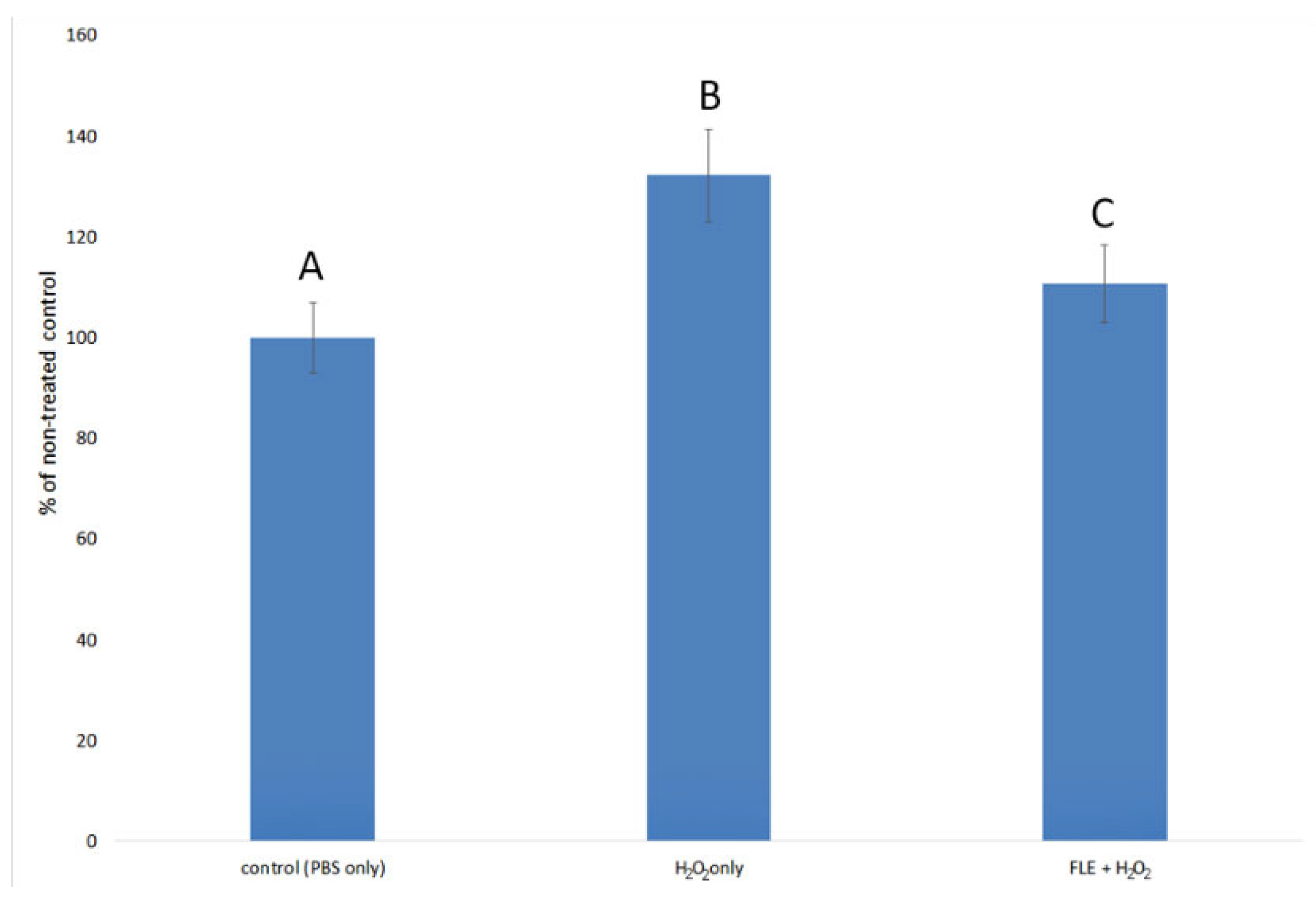
3.3. Effect of Extract on H2O2-Induced Oxidative Stress in Normal Fibroblast Cells
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Azra, M.N.; Okomoda, V.T.; Tabatabaei, M.; Hassan, M.; Ikhwanuddin, M. The Contributions of Shellfish Aquaculture to Global Food Security: Assessing Its Characteristics From a Future Food Perspective. Front. Mar. Sci. 2021, 8, 365. [Google Scholar] [CrossRef]
- Olatunde, O.O.; Benjakul, S. Antioxidants from Crustaceans: A Panacea for Lipid Oxidation in Marine-Based Foods. Food Rev. Int. 2020, 38, 1–31. [Google Scholar] [CrossRef]
- Kaldre, K.; Paaver, T.; Hurt, M.; Gross, R. Continuing expansion of non-indigenous crayfish species in Northern Europe: First established spiny-cheek crayfish Faxonius limosus (Refinesque, 1817) population in Estonia. Rec. Bioinvasions 2020, 9, 127–132. [Google Scholar] [CrossRef]
- Kouba, A.; Petrusek, A.; Kozák, P. Continental-wide distribution of crayfish species in Europe: Update and maps. Knowl. Manag. Aquat. Ecosyst. 2014, 413, 05. [Google Scholar] [CrossRef]
- Śmietana, N.; Panicz, R.; Sobczak, M.; Śmietana, P.; Nędzarek, A. Spiny-Cheek Crayfish, Faxonius limosus (Rafinesque, 1817), as an Alternative Food Source. Animals 2021, 11, 59. [Google Scholar] [CrossRef]
- Dabrowski, T.; Kolakowski, E.; Wawreszuk, H.; Choroszucha, C. Studies on Chemical Composition of American Crayfish (Orconectes limosus) Meat as Related to its Nutritive Value. J. Fish. Board Can. 2011, 23, 1653–1662. [Google Scholar] [CrossRef]
- Czeczuga, B.; Czeczuga-Semeniuk, E. Comparative studies of carotenoids in four species of crayfish. Crustaceana 1999, 72, 693–700. [Google Scholar] [CrossRef]
- Guidelines for the treatment of animals in behavioural research and teaching. Anim. Behav. 1997, 53, 229–234. [CrossRef]
- Zglińska, K.; Niemiec, T.; Łozicki, A.; Matusiewicz, M.; Szczepaniak, J.; Puppel, K.; Kutwin, M.; Jaworski, S.; Rygało-Galewska, A.; Koczoń, P. Effect of Elaeagnus umbellata (Thunb.) fruit extract on H2O2-induced oxidative and inflammatory responses in normal fibroblast cells. PeerJ 2021, 9, e10760. [Google Scholar] [CrossRef]
- Matusiewicz, M.; Bączek, K.B.; Kosieradzka, I.; Niemiec, T.; Grodzik, M.; Szczepaniak, J.; Orlińska, S.; Węglarz, Z. Effect of Juice and Extracts from Saposhnikovia divaricata Root on the Colon Cancer Cells Caco-2. Int. J. Mol. Sci. 2019, 20, 4526. [Google Scholar] [CrossRef]
- Zglińska, K.; Rygało-Galewska, A.; Bryś, J.; Koczoń, P.; Borek, K.; Roguski, M.; Niemiec, T. Elaeagnus umbellata fruit - chemical composition, bioactive compounds, and kinetic of DPPH inhibition compared to standard antioxidants. Emirates J. Food Agric. 2021, 33, 639–646. [Google Scholar] [CrossRef]
- Kim, H.; Xue, X. Detection of Total Reactive Oxygen Species in Adherent Cells by 2′,7′-Dichlorodihydrofluorescein Diacetate Staining. J. Vis. Exp. 2020, 2020, 1–5. [Google Scholar] [CrossRef]
- Safafar, H.; Van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, Phenolic Compounds and Tocopherols Contribute to the Antioxidative Properties of Some Microalgae Species Grown on Industrial Wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef]
- Winston, G.W.; Lemaire, D.G.E.; Lee, R.F. Antioxidants and total oxyradical scavenging capacity during grass shrimp, Palaemonetes pugio, embryogenesis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2004, 139, 281–288. [Google Scholar] [CrossRef]
- Moro, J.; Tomé, D.; Schmidely, P.; Demersay, T.C.; Azzout-Marniche, D. Histidine: A Systematic Review on Metabolism and Physiological Effects in Human and Different Animal Species. Nutrients 2020, 12, 1414. [Google Scholar] [CrossRef]
- Sachindra, N.M.; Bhaskar, N. In vitro antioxidant activity of liquor from fermented shrimp biowaste. Bioresour. Technol. 2008, 99, 9013–9016. [Google Scholar] [CrossRef]
- Proestos, C.; Lytoudi, K.; Mavromelanidou, O.K.; Zoumpoulakis, P.; Sinanoglou, V.J. Antioxidant Capacity of Selected Plant Extracts and Their Essential Oils. Antioxidants 2013, 2, 11–22. [Google Scholar] [CrossRef]
- Sinha, R.; Sinha, I.; Calcagnotto, A.; Trushin, N.; Haley, J.S.; Schell, T.D.; Richie, J.P. Oral supplementation with liposomal glutathione elevates body stores of glutathione and markers of immune function. Eur. J. Clin. Nutr. 2018, 72, 105–111. [Google Scholar] [CrossRef]
- Valdivia, A.; Ly, J.; Gonzalez, L.; Hussain, P.; Saing, T.; Islamoglu, H.; Pearce, D.; Ochoa, C.; Venketaraman, V. Restoring Cytokine Balance in HIV-Positive Individuals with Low CD4 T Cell Counts. AIDS Res. Hum. Retroviruses 2017, 33, 905–918. [Google Scholar] [CrossRef]
- Pereira, R.B.; Sousa, C.; Costa, A.; Andrade, P.B.; Valentão, P. Glutathione and the Antioxidant Potential of Binary Mixtures with Flavonoids: Synergisms and Antagonisms. Molecules 2013, 18, 8858–8872. [Google Scholar] [CrossRef]
- Płonka, J.; Szablińska-Piernik, J.; Buszewski, B.; Baranowska, I.; Lahuta, L.B. Analyses of Antioxidative Properties of Selected Cyclitols and Their Mixtures with Flavanones and Glutathione. Molecules 2021, 27, 158. [Google Scholar] [CrossRef]
- Zhou, Y.; Dong, Y.; Xu, Q.G.; Zhu, S.Y.; Tian, S.L.; Huo, J.J.; Hao, T.T.; Zhu, B.W. Mussel oligopeptides protect human fibroblasts from hydrogen peroxide (H2O2)-induced premature senescence. Arch. Gerontol. Geriatr. 2014, 58, 293–299. [Google Scholar] [CrossRef]
- Qian, Z.J.; Jung, W.K.; Byun, H.G.; Kim, S.K. Protective effect of an antioxidative peptide purified from gastrointestinal digests of oyster, Crassostrea gigas against free radical induced DNA damage. Bioresour. Technol. 2008, 99, 3365–3371. [Google Scholar] [CrossRef]
- Oh, Y.; Ahn, C.B.; Je, J.Y. Cytoprotective Role of Edible Seahorse (Hippocampus abdominalis)-Derived Peptides in H2O2-Induced Oxidative Stress in Human Umbilical Vein Endothelial Cells. Mar. Drugs 2021, 19, 86. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Kim, E.K.; Dong, X.; Shin, W.B.; Park, J.S.; Kim, S.J.; Go, E.A.; Byun, H.G.; Park, P.J. Antioxidant and Protective Effects of Atrina Pectinata Extract. Adv. Exp. Med. Biol. 2019, 1155, 627–641. [Google Scholar] [CrossRef]
- Fahmy, S.R.; Hamdi, S.A.H. Antioxidant effect of the Egyptian freshwater Procambarus clarkii extract in rat liver and erythrocytes. Afr. J. Pharm. Pharmacol. 2011, 5, 776–785. [Google Scholar] [CrossRef]
- Giampieri, F.; Alvarez-Suarez, J.M.; Mazzoni, L.; Forbes-Hernandez, T.Y.; Gasparrini, M.; Gonzàlez-Paramàs, A.M.; Santos-Buelga, C.; Quiles, J.L.; Bompadre, S.; Mezzetti, B.; et al. An anthocyanin-rich strawberry extract protects against oxidative stress damage and improves mitochondrial functionality in human dermal fibroblasts exposed to an oxidizing agent. Food Funct. 2014, 5, 1939–1948. [Google Scholar] [CrossRef]
- Pongcharoen, S.; Warnnissorn, P.; Leŗtkajornsin, O.; Limpeanchob, N.; Sutheerawattananonda, M. Protective effect of silk lutein on ultraviolet B-irradiated human keratinocytes. Biol. Res. 2013, 46, 39–45. [Google Scholar] [CrossRef]
- Talero, E.; García-Mauriño, S.; Ávila-Román, J.; Rodríguez-Luna, A.; Alcaide, A.; Motilva, V. Bioactive Compounds Isolated from Microalgae in Chronic Inflammation and Cancer. Mar. Drugs 2015, 13, 6152–6209. [Google Scholar] [CrossRef]
- Datta, D.; Bhinge, A.; Chandran, V. Lysine: Is it worth more? Cytotechnology 2001, 36, 3–32. [Google Scholar] [CrossRef]
- Ponrasu, T.; Jamuna, S.; Mathew, A.; Madhukumar, K.N.; Ganeshkumar, M.; Iyappan, K.; Suguna, L. Efficacy of l-proline administration on the early responses during cutaneous wound healing in rats. Amino Acids 2013, 45, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Karna, E.; Szoka, L.; Huynh, T.Y.L.; Palka, J.A. Proline-dependent regulation of collagen metabolism. Cell. Mol. Life Sci. 2020, 77, 1911–1918. [Google Scholar] [CrossRef] [PubMed]
- Addis, R.; Cruciani, S.; Santaniello, S.; Bellu, E.; Sarais, G.; Ventura, C.; Maioli, M.; Pintore, G. Fibroblast Proliferation and Migration in Wound Healing by Phytochemicals: Evidence for a Novel Synergic Outcome. Int. J. Med. Sci. 2020, 17, 1030. [Google Scholar] [CrossRef] [PubMed]
- Ponnusamy, Y.; Chear, N.J.Y.; Ramanathan, S.; Lai, C.S. Polyphenols rich fraction of Dicranopteris linearis promotes fibroblast cell migration and proliferation in vitro. J. Ethnopharmacol. 2015, 168, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Galati, G.; O’Brien, P.J. Potential toxicity of flavonoids and other dietary phenolics: Significance for their chemopreventive and anticancer properties. Free Radic. Biol. Med. 2004, 37, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Niemiec, T.; Sawosz, E.; Chwalibog, A. Effect of high doses of L-ascorbic acid on theantioxidative/oxidative state in the rats. J. Anim. Feed Sci. 2005, 14, 553–556. [Google Scholar] [CrossRef][Green Version]
- Palozza, P.; Serini, S.; Di Nicuolo, F.; Piccioni, E.; Calviello, G. Prooxidant effects of β-carotene in cultured cells. Mol. Aspects Med. 2003, 24, 353–362. [Google Scholar] [CrossRef]
- Bayliak, M.M.; Burdyliuk, N.I.; Lushchak, V.I. Effects of pH on antioxidant and prooxidant properties of common medicinal herbs. Open Life Sci. 2016, 11, 298–307. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Content |
|---|---|
| Total phenols (µg/mL 1) | 13.321 |
| GSH (nmol/mg) | 10.755 |
| β-carotene (mg/g) | 0.029 |
| Vitamin A (mg/g) | 0.085 |
| Vitamin E (mg/g) | 0.785 |
| Vitamin D (µg/g) | 0.084 |
| Vitamin K (µg/g) | 0.104 |
| Compound | mg/g Crayfish Extract | CS |
|---|---|---|
| His | 28.91 | 128.65 |
| Ile | 27.94 | 50.33 |
| Leu | 60.75 | 65.66 |
| Lys | 61.78 | 91.65 |
| TSAA | 28.77 | 83.73 |
| TAAA | 75.24 | 77.99 |
| Thr | 38.75 | 83.77 |
| Trp | 9.83 | 67.64 |
| Val | 41.77 | 65.83 |
| Pro | 29.77 | - |
| Glu | 101.72 | - |
| Crude protein | 756.60 | X |
| Group | % of Cytotoxicity | p < 0.05 |
|---|---|---|
| Control (PBS only) | 0.187 | A |
| H2O2 only | 43.925 | B |
| FLE (50 µg/mL) + H2O2 | 0.639 | A |
| FLE (100 µg/mL) + H2O2 | 0.019 | A |
| FLE (200 µg/mL) + H2O2 | 2.045 | C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zglińska, K.; Jaworski, S.; Rygało-Galewska, A.; Łozicki, A.; Roguski, M.; Matusiewicz, M.; Niemiec, T. Effect of Spiny-Cheek Crayfish (Faxonius limosus) on H2O2-Induced Oxidative Stress in Normal Fibroblast Cells. Appl. Sci. 2022, 12, 8546. https://doi.org/10.3390/app12178546
Zglińska K, Jaworski S, Rygało-Galewska A, Łozicki A, Roguski M, Matusiewicz M, Niemiec T. Effect of Spiny-Cheek Crayfish (Faxonius limosus) on H2O2-Induced Oxidative Stress in Normal Fibroblast Cells. Applied Sciences. 2022; 12(17):8546. https://doi.org/10.3390/app12178546
Chicago/Turabian StyleZglińska, Klara, Sławomir Jaworski, Anna Rygało-Galewska, Andrzej Łozicki, Mateusz Roguski, Magdalena Matusiewicz, and Tomasz Niemiec. 2022. "Effect of Spiny-Cheek Crayfish (Faxonius limosus) on H2O2-Induced Oxidative Stress in Normal Fibroblast Cells" Applied Sciences 12, no. 17: 8546. https://doi.org/10.3390/app12178546
APA StyleZglińska, K., Jaworski, S., Rygało-Galewska, A., Łozicki, A., Roguski, M., Matusiewicz, M., & Niemiec, T. (2022). Effect of Spiny-Cheek Crayfish (Faxonius limosus) on H2O2-Induced Oxidative Stress in Normal Fibroblast Cells. Applied Sciences, 12(17), 8546. https://doi.org/10.3390/app12178546