
In Vitro Antioxidant, Antibacterial and Mechanisms of Action of Ethanolic Extracts of Five Tunisian Plants against Bacteria

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Polyphenol Extraction
2.2.1. Preparation of Ethanolic Extracts (EE)
2.2.2. Total Polyphenol Contents
2.2.3. Total Flavonoid Contents
2.2.4. Condensed Tannin Content
2.3. Antioxidant Activity Evaluation
2.3.1. Evaluation of the Total Antioxidant Activity (TAA)
2.3.2. DPPH Radical Scavenging Activity
2.3.3. Determination of the Ferric-Reducing Antioxidant Power (FRAP)
2.4. Antibacterial Activity Assessment
2.5. Mode of Action of Ethanolic Extract
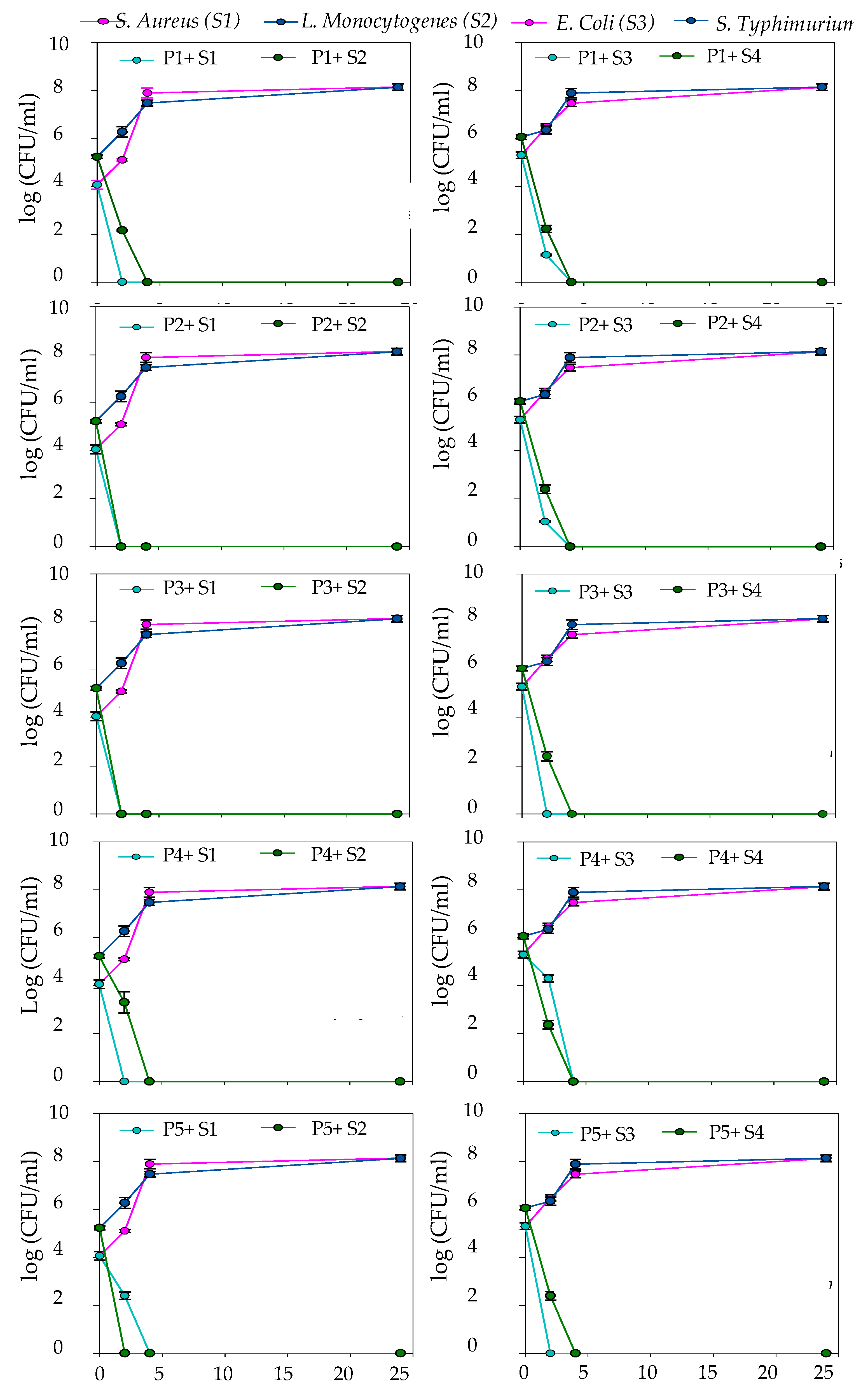
2.5.1. Time Kill Assay
2.5.2. Bacteriolysis
2.5.3. Loss of Salt Tolerance
2.6. Statistical Analyses
3. Results
3.1. Determination of Total Phenols, Flavonoids, and Tannin Contents
3.2. Biological Activities
3.2.1. Determination of Antioxidant Activity
3.2.2. Determination of Minimum Inhibitory and Bactericidal Concentrations
3.2.3. Mode of Action of Ethanolic Extract
Time Kill Assays
Determination of the Lytic Action of Ethanolic Extracts
3.2.4. Loss of Salt Tolerance
3.3. Bioactive Compounds Investigation by HPLC Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Parekh, J. In vitro screening of antibacterial activity of aqueous and alcoholic extracts of various Indian plant species against selected pathogens from Enterobacteriaceae. Afr. J. Microbiol. Res. 2007, 1, 92–99. [Google Scholar]
- Aulifa, D.L.; Fitriansyah, S.N.; Ardiansyah, S.A.; Wibowo, D.P.; Julata, Y.A.; Christy, D.S. Phytochemical screening, antibacterial activity, and mode of action on Morus nigra. Pharmacogn. J. 2018, 10, 167–171. [Google Scholar] [CrossRef]
- Saavedra-Leos, M.Z.; Leyva-Porras, C.; Toxqui-Terán, A.; Espinosa-Solis, V. Physicochemical Properties and Antioxidant Activity of Spray-Dry Broccoli (Brassica oleracea var Italica) Stalk and Floret Juice Powders. Molecules 2021, 26, 1973. [Google Scholar] [CrossRef] [PubMed]
- Gonelimali, F.D.; Lin, J.; Miao, W.; Xuan, J.; Charles, F.; Chen, M.; Hatab, S.R. Antimicrobial properties and mechanism of action of some plant extracts against food pathogens and spoilage microorganisms. Front. Microbiol. 2018, 9, 1639. [Google Scholar] [CrossRef]
- Adeniyi, A.; Asase, A.; Ekpe, P.K.; Asitoakor, B.K.; Adu-Gyamfi, A.; Avekor, P.Y. Ethnobotanical study of medicinal plants from Ghana; confirmation of ethnobotanical uses, and review of biological and toxicological studies on medicinal plants used in Apra Hills Sacred Grove. J. Herb. Med. 2018, 14, 76–87. [Google Scholar] [CrossRef]
- Perrino, E.; Tomaselli, V.; Costa, R.; Pavone, P. Conservation status of habitats (Directive 92/43 EEC) of coastal and low hill belts in a Mediterranean biodiversity hot spot (Gargano–Italy). Plant Biosyst.-Int. J. Deal. All Asp. Plant Biol. 2013, 147, 1006–1028. [Google Scholar] [CrossRef]
- Talib, W.H.; Mahasneh, A.M. Antimicrobial, cytotoxicity and phytochemical screening of Jordanian plants used in traditional medicine. Molecules 2010, 15, 1811–1824. [Google Scholar] [CrossRef]
- Elkhamlichi, A.; El Antri, A.; El Hajaji, H.; El Bali, B.; Oulyadi, H.; Lachkar, M. Phytochemical constituents from the seeds of Calycotome villosa subsp. intermedia. Arab. J. Chem. 2017, 10, S3580–S3583. [Google Scholar] [CrossRef]
- Fatma, G.; Sami, B.H.A.; Ahmed, L. Investigation of extracts from Tunisian ethnomedicinal plants as antioxidants, cytotoxins, and antimicrobials. Biomed. Environ. Sci. 2017, 30, 811–824. [Google Scholar]
- Romani, A.; Pinelli, P.; Galardi, C.; Mulinacci, N.; Tattini, M. Identification and quantification of galloyl derivatives, flavonoid glycosides and anthocyanins in leaves of Pistacia lentiscus L. Phytochem. Anal. Int. J. Plant Chem. Tech. 2002, 13, 79–86. [Google Scholar] [CrossRef]
- Azaizeh, H.; Halahleh, F.; Abbas, N.; Markovics, A.; Muklada, H.; Ungar, E.; Landau, S. Polyphenols from Pistacia lentiscus and Phillyrea latifolia impair the exsheathment of gastro-intestinal nematode larvae. Vet. Parasitol. 2013, 191, 44–50. [Google Scholar] [CrossRef]
- Amaral, G.P.; de Carvalho, N.R.; Barcelos, R.P.; Dobrachinski, F.; de Lima Portella, R.; da Silva, M.H.; Lugokenski, T.H.; Dias, G.R.M.; da Luz, S.C.A.; Boligon, A.A. Protective action of ethanolic extract of Rosmarinus officinalis L. in gastric ulcer prevention induced by ethanol in rats. Food Chem. Toxicol. 2013, 55, 48–55. [Google Scholar] [CrossRef]
- Khan, U.A.; Rahman, H.; Niaz, Z.; Qasim, M.; Khan, J.; Tayyaba; Rehman, B. Antibacterial activity of some medicinal plants against selected human pathogenic bacteria. Eur. J. Microbiol. Immunol. 2013, 3, 272–274. [Google Scholar] [CrossRef]
- Ekor, M. The growing use of herbal medicines: Issues relating to adverse reactions and challenges in monitoring safety. Front. Pharmacol. 2014, 4, 177. [Google Scholar] [CrossRef]
- Mohan, S.; Abdul, A.B.; Wahab, S.I.A.; Al-Zubairi, A.S.; Elhassan, M.M.; Yousif, M. Antibacterial and antioxidant activities of Typhonium flagelliforme (Lodd) Blume tuber. Am. J. Biochem. Biotechnol. 2008, 4, 402–407. [Google Scholar] [CrossRef][Green Version]
- Nostro, A.; Germano, M.; D’angelo, V.; Marino, A.; Cannatelli, M. Extraction methods and bioautography for evaluation of medicinal plant antimicrobial activity. Lett. Appl. Microbiol. 2000, 30, 379–384. [Google Scholar] [CrossRef]
- Saritha, K.; Rajesh, A.; Manjulatha, K.; Setty, O.H.; Yenugu, S. Mechanism of antibacterial action of the alcoholic extracts of Hemidesmus indicus (L.) R. Br. ex Schult, Leucas aspera (Wild.), Plumbago zeylanica L., and Tridax procumbens (L.) R. Br. ex Schult. Front. Microbiol. 2015, 6, 577. [Google Scholar] [CrossRef]
- Sudipta, K.; Lokesh, P.; Rashmi, W.; Vijay, R.; Ssn, K.J. Phytochemical screening and in vitro antimicrobial activity of Bougainvillea spectabilis flower extracts. Int. J. Phytomed. 2012, 4, 375. [Google Scholar]
- Christaki, E.; Bonos, E.; Giannenas, I.; Florou-Paneri, P.J. Aromatic plants as a source of bioactive compounds. Agriculture 2012, 2, 228–243. [Google Scholar] [CrossRef]
- Trigui, M.; Hsouna, A.B.; Tounsi, S.; Jaoua, S. Chemical composition and evaluation of antioxidant and antimicrobial activities of Tunisian Thymelaea hirsuta with special reference to its mode of action. Ind. Crops Prod. 2013, 41, 150–157. [Google Scholar] [CrossRef]
- Tomlinson, S.; Palombo, E.A. Characterisation of antibacterial Australian medicinal plant extracts by investigation of the mechanism of action and the effect of interfering substances. J. Basic Microbiol. Int. J. Biochem. Physiol. Genet. Morphol. Ecol. Microorg. 2005, 45, 363–370. [Google Scholar] [CrossRef]
- Dewanto, V.; Wu, X.; Adom, K.K.; Liu, R.H. Thermal processing enhances the nutritional value of tomatoes by increasing total antioxidant activity. J. Agric. Chem. 2002, 50, 3010–3014. [Google Scholar] [CrossRef]
- Broadhurst, R.B.; Jones, W.T.J. Analysis of condensed tannins using acidified vanillin. J. Sci. Food 1978, 29, 788–794. [Google Scholar] [CrossRef]
- Sun, B.; Ricardo-da-Silva, J.M.; Spranger, I.J. Critical factors of vanillin assay for catechins and proanthocyanidins. J. Agric. Chem. 1998, 46, 4267–4274. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M.J. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Burits, M.; Bucar, F.J. Antioxidant activity of Nigella sativa essential oil. Phytother. Res. 2000, 14, 323–328. [Google Scholar] [CrossRef]
- Cuendet, M.; Hostettmann, K.; Potterat, O.; Dyatmiko, W.J. Iridoid glucosides with free radical scavenging properties from Fagraea blumei. Helv. Chim. Acta 1997, 80, 1144–1152. [Google Scholar] [CrossRef]
- Oyaizu, M. Antioxidant activity of brewing products of glucosamine fractionated by organic solvents and their layer chromatography. Nippon. Skokulin Kogyo Gakkaishi 1988, 35, 771–775. [Google Scholar] [CrossRef]
- Shortridge, D.; Streit, J.M.; Mendes, R.; Castanheira, M.J. In Vitro Activity of Cefiderocol against US and European Gram-Negative Clinical Isolates Collected in 2020 as Part of the SENTRY Antimicrobial Surveillance Program. Microbiol. Spectr. 2022, 10, e02712-21. [Google Scholar] [CrossRef]
- Cosentino SC, I.G.; Tuberoso CI, G.; Pisano, B.; Satta, M.L.; Mascia, V.; Arzedi, E.; Palmas, F. In vitro antimicrobial activity and chemical composition of Sardinian thymus essential oils. Lett. Appl. Microbiol. 1999, 29, 130–135. [Google Scholar] [CrossRef]
- Carson, C.F.; Mee, B.J.; Riley, T.V. Mechanism of action of Melaleuca alternifolia (tea tree) oil on Staphylococcus aureus determined by time-kill, lysis, leakage, and salt tolerance assays and electron microscopy. Antimicrob. Agents 2002, 46, 1914–1920. [Google Scholar] [CrossRef] [PubMed]
- Escarpa, A.; González, M. Approach to the content of total extractable phenolic compounds from different food samples by comparison of chromatographic and spectrophotometric methods. Anal. Chim. Acta 2001, 427, 119–127. [Google Scholar]
- Turan, M.; Mammadov, R. Antioxidant, Cytotoxic, Larvicidal, and Anthelmintic Activity and Phytochemical Screening by HPLC of Calicotome villosa from Turkey. Pharm. Chem. J. 2020, 54, 478–483. [Google Scholar] [CrossRef]
- Cheung, S.; Tai, J. Anti-proliferative and antioxidant properties of rosemary Rosmarinus officinalis. Oncol. Rep. 2007, 17, 1525–1531. [Google Scholar] [CrossRef]
- Gu, C.; Howell, K.; Dunshea, F.R.; Suleria, H.A.J. Lc-esi-qtof/ms characterisation of phenolic acids and flavonoids in polyphenol-rich fruits and vegetables and their potential antioxidant activities. Antioxidants 2019, 8, 405. [Google Scholar] [CrossRef]
- Van Loy, M.D.; Riley, W.J.; Daisey, J.M.; Nazaroff, W.W.J.E.S. Dynamic behavior of semivolatile organic compounds in indoor air. 2. Nicotine and phenanthrene with carpet and wallboard. Environ. Sci. Technol. 2001, 35, 560–567. [Google Scholar] [CrossRef]
- Benhammou, N.; Bekkara, F.A.; Panovska, T.K.J. Antiradical capacity of the phenolic compounds of Pistacia lentiscus L. And Pistacia atlantica desf. Adv. Food Sci. 2007, 29, 155–161. [Google Scholar]
- Boughalleb, F.; Mahmoudi, M.; Abdellaoui, R.; Yahia, B.; Zaidi, S.; Nasri, N.J. Effect of long—Term storage on phenolic composition, antioxidant capacity, and protein profiles of Calicotome villosa subsp. intermedia seeds. J. Food Biochem. 2020, 44, e13093. [Google Scholar] [CrossRef]
- Mathew, S.; Abraham, T.E.; Zakaria, Z.A.J. Reactivity of phenolic compounds towards free radicals under in vitro conditions. J. Food Sci. Technol. 2015, 52, 5790–5798. [Google Scholar] [CrossRef]
- Fathollahi, M.; Aminzare, M.; Mohseni, M.; Hassanzadazar, H. Antioxidant capacity, antimicrobial activities and chemical composition of Pistacia atlantica subsp. kurdica essential oil. Vet. Res. Forum 2019, 10, 299–305. [Google Scholar]
- Pyo, Y.H.; Lee, T.C.; Logendra, L.; Rosen, R.T. Antioxidant activity and phenolic compounds of Swiss chard (Beta vulgaris subspecies cycla) extracts. Food Chem. 2004, 85, 19–26. [Google Scholar] [CrossRef]
- Mahmoudi, H.; Aouadhi, C.; Kaddour, R.; Gruber, M.; Zargouni, H.; Zaouali, W.; Ben Hamida, N.; Ben Nasri, M.; Ouerghi, Z.; Hosni, K. Comparison of antioxidant and antimicrobial activities of two cultivated Cistus species from Tunisia. Biosci. J. 2016, 32, 226–237. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total phenolic content, flavonoid content and antioxidant potential of wild vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef]
- Muflihah, Y.M.; Gollavelli, G.; Ling, Y.-C.J.A. Correlation Study of Antioxidant Activity with Phenolic and Flavonoid Compounds in 12 Indonesian Indigenous Herbs. Antioxidants 2021, 10, 1530. [Google Scholar] [CrossRef]
- Olajuyigbe, O.O.; Afolayan, A.J. In vitro antibacterial and time-kill assessment of crude methanolic stem bark extract of Acacia mearnsii De Wild against bacteria in shigellosis. Molecules 2012, 17, 2103–2118. [Google Scholar] [CrossRef]
- Burt, S. Essential oils: Their antibacterial properties and potential applications in foods—A review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef]
- Oonmetta-aree, J.; Suzuki, T.; Gasaluck, P.; Eumkeb, G. Antimicrobial properties and action of galangal (Alpinia galanga Linn.) on Staphylococcus aureus. LWT-Food Sci. 2006, 39, 1214–1220. [Google Scholar] [CrossRef]
- Kollef, M.H.; Golan, Y.; Micek, S.T.; Shorr, A.F.; Restrepo, M.I.J.C.i.d. Appraising contemporary strategies to combat multidrug resistant gram-negative bacterial infections–proceedings and data from the Gram-Negative Resistance Summit. Clin. Infect. Dis. 2011, 53, S33–S55. [Google Scholar] [CrossRef]
- Beveridge, T.J. Structures of gram-negative cell walls and their derived membrane vesicles. J. Bacteriol. 1999, 181, 4725–4733. [Google Scholar] [CrossRef]
- Mastino, P.M.; Marchetti, M.; Costa, J.; Juliano, C.; Usai, M. Analytical Profiling of Phenolic Compounds in Extracts of Three Cistus Species from Sardinia and Their Potential Antimicrobial and Antioxidant Activity. Chem. Biodivers. 2021, 18, e2100053. [Google Scholar] [CrossRef]
- Shimoda, M.; Ohki, K.; Shimamoto, Y.; Kohashi, O. Morphology of defensin-treated Staphylococcus aureus. Infect. Immun. 1995, 63, 2886–2891. [Google Scholar] [CrossRef]
- Riahi, L.; Elferchichi, M.; Ghazghazi, H.; Jebali, J.; Ziadi, S.; Aouadhi, C.; Chograni, H.; Zaouali, Y.; Zoghlami, N.; Mliki, A. Phytochemistry, antioxidant and antimicrobial activities of the essential oils of Mentha rotundifolia L. in Tunisia. Ind. Crops Prod. 2013, 49, 883–889. [Google Scholar] [CrossRef]
- Gilbert, P.; Collier, P.J.; Brown, M. Influence of growth rate on susceptibility to antimicrobial agents: Biofilms, cell cycle, dormancy, and stringent response. Antimicrob. Agents 1990, 34, 1865–1868. [Google Scholar] [CrossRef]
- Horne, D.; Holm, M.; Oberg, C.; Chao, S.; Young, D.G. Antimicrobial effects of essential oils on Streptococcus pneumoniae. J. Essent. Oil Res. 2001, 13, 387–392. [Google Scholar] [CrossRef]
- Jenie, B.S.L.; Priosoeryanto, B.P.; Syarief, R.; Rekso, G.T. Mode of action Temu kunci (Kaempferia pandurata) essential oil on E. coli K1. 1 cell determined by leakage of material cell and salt tolerance assays. HAYATI J. Biosci. 2008, 15, 56–60. [Google Scholar]
- Cushnie, T.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Hamburger, M.; Hostettmann, K. 7. Bioactivity in plants: The link between phytochemistry and medicine. Phytochemistry 1991, 30, 3864–3874. [Google Scholar] [CrossRef]
- Wagner, H.; Ulrich-Merzenich, G. Synergy research: Approaching a new generation of phytopharmaceuticals. Phytomedicine 2009, 16, 97–110. [Google Scholar] [CrossRef]
- Ikeuchi, T.; Sisodia, S.S. The Notch ligands, Delta1 and Jagged2, are substrates for presenilin-dependent “γ-secretase” cleavage. J. Biol. Chem. 2003, 278, 7751–7754. [Google Scholar] [CrossRef]
- Brown, A.; Turner, H. Membrane stability and salt tolerance in gram-negative bacteria. Nature 1963, 199, 301–302. [Google Scholar] [CrossRef]
- Zhu, X.; Dong, N.; Wang, Z.; Ma, Z.; Zhang, L.; Ma, Q.; Shan, A. Design of imperfectly amphipathic α-helical antimicrobial peptides with enhanced cell selectivity. Acta Biomater. 2014, 10, 244–257. [Google Scholar] [CrossRef] [PubMed]
- Tariq, S.; Wani, S.; Rasool, W.; Shafi, K.; Bhat, M.A.; Prabhakar, A.; Shalla, A.H.; Rather, M.A. A comprehensive review of the antibacterial, antifungal and antiviral potential of essential oils and their chemical constituents against drug-resistant microbial pathogens. Microb. Pathog. 2019, 134, 103580. [Google Scholar] [CrossRef] [PubMed]
- Iandolo, J.J.; Ordal, Z.J. Repair of thermal injury of Staphylococcus aureus. J. Bacteriol. 1966, 91, 134–142. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| TAC (GAE/gDW) | DPPH (IC50 mg/mL) | FRAP (EC50 mg/mL) | |
|---|---|---|---|
| Pistacia lentiscus | 13.50 c ± 1.50 | 5.38 e ± 0.44 | 13.39 e ± 1.29 |
| Rosmarinus officinalis | 12.10 d ± 0.53 | 12.25 c ± 0.24 | 20.82 b ± 0.52 |
| Erica multiflora | 13.45 c ± 2.48 | 10.85 d ± 0.99 | 17.89 d ± 0.66 |
| Calicotome villosa | 15.33 b ± 0.39 | 19.60 a ± 0.39 | 24.2 a ± 0.11 |
| Phillyrea latifolia | 20.16 a ± 0.32 | 15.59 b ± 0.66 | 19.35 c ± 0.26 |
| Inhibition Zone Diameter (mm) | MIC (mg/mL) | BMC (mg/mL) | ||
|---|---|---|---|---|
| EE | Gentamicin (10 mg/disc) | |||
| Staphylococcus aureus | ||||
| Pistacia lentiscus | 15 c ± 0.8 | 24 a ± 0.1 | 12 | 25 |
| Rosmarinus officinalis | 14.5 ± 0.2 cd | 3.84 | 12 | |
| Erica multiflora | 16 b ± 0.1 | 0.04 | 3.84 | |
| Calicotome villosa | 13 d ± 0.3 | 25 | 25 | |
| Phillyrea latifolia | 16 b ± 0.5 | 3.84 | 12 | |
| Escherichia coli | ||||
| Pistacia lentiscus | 15 c ± 0.7 | 25 a ± 0.08 | 12 | 25 |
| Rosmarinus officinalis | 14 d ± 0.00 | 3.84 | 12 | |
| Erica multiflora | 16 b ± 0.3 | 0.04 | 3.84 | |
| Calicotome villosa | 15 c ± 0.5 | 25 | 25 | |
| Phillyrea latifolia | 13 e ± 0.5 | 3.84 | 12 | |
| Listeria monocytogenes | ||||
| Pistacia lentiscus | 13 d ± 0.4 | 25 a ± 0.1 | 0.04 | 3.84 |
| Rosmarinus officinalis | 14 c ± 0.5 | 3.84 | 3.84 | |
| Erica multiflora | 15 b ± 0.1 | 3.84 | 3.84 | |
| Calicotome villosa | 14 c ± 0.3 | 3.84 | 12 | |
| Phillyrea latifolia | 15 b ± 0.5 | 3.84 | 12 | |
| Salmonella typhimurium | ||||
| Pistacia lentiscus | 15 b ± 0.2 | 25 a ± 0.1 | 3.84 | 12 |
| Rosmarinus officinalis | 13 c ± 0.4 | 12 | 25 | |
| Erica multiflora | 14 c ± 0.7 | 3.84 | 12 | |
| Calicotome villosa | 14 c ± 0.2 | 3.84 | 12 | |
| Phillyrea latifolia | 14 c ± 0.5 | 3.84 | 12 | |
| Treatment Agents (MICs) and Strains | OD 620 at (T) min/OD 620 at 0 min (%) | |||
|---|---|---|---|---|
| Immediately | After 60 min | After 120 min | After 24 h | |
| Control: S. aureus | 103.1 a ± 6.17 | 94.7 a ± 4.28 | 91.7 a ± 1.31 | 102.5 a ± 4.23 |
| Control: E. coli | 98.2 a ± 0.66 | 99.1 a ± 0.04 | 95.6 a ± 1.70 | 98.4 a ± 0.78 |
| Control: L. monocytogenes | 98.22 a ± 0.66 | 99.21 a ± 0.8 | 98.35 a ± 0.90 | 99.40 a ± 0.41 |
| Control: S. typhimurium | 100.02 a ± 3.01 | 99.01 a ± 1.00 | 98.76 a ± 0.55 | 101.02 a ± 3.46 |
| P. lentiscus + S. aureus | 96.56 a ± 01.13 | 83.49 b ± 0.90 | 59.48 c ± 0.82 | 25.21 d ± 1.75 |
| P. lentiscus + E. coli | 98.43 a ± 0.51 | 67.61 b ± 0.90 | 51.03 c ± 1.05 | 27.61 d ± 1.78 |
| P. lentiscus + L. monocytogenes | 98.36 a ± 0.39 | 66.59 b ± 0.90 | 47.54 c ± 1.88 | 22.87 d ± 2.13 |
| P. lentiscus + S. typhimurium | 97.58 a ± 0.64 | 70.21b ± 0.33 | 56.31 c ± 1.04 | 22.22 d ± 1.57 |
| R. offinalis + S. aureus | 98.21 a ± 1.01 | 67.10 b ± 0.01 | 59.42 bc ± 0.74 | 31.14 c ± 1.22 |
| R. offinalis + E. coli | 98.77 a ± 0.68 | 80.37 b ± 0.85 | 49.17 c ± 1.50 | 35.4 d ± 0.75 |
| R. offinalis + L. monocytogenes | 98.43 a ± 0.74 | 80.74 b ± 0.70 | 50.52 c ± 0.08 | 25.6 d ± 1.37 |
| R. offinalis + S. typhimurium | 98.43 a ± 1.05 | 73.25 b ± 2.29 | 57.73 c ± 0.63 | 35.10 d ± 0.78 |
| E. multiflora + S. aureus | 97.49 a ± 0.14 | 66.77 b ± 1.67 | 57.98 c ± 2.22 | 31.53 d ± 2.33 |
| E. multiflora + E. coli | 97.96 a ± 0.50 | 67.48 b ± 0.22 | 53.54 c ± 2.87 | 25.48 d ± 3.56 |
| E. multiflora + L. monocytogenes | 97.63 a ± 0.31 | 56.22 b ± 1.49 | 55.55 b ± 1.03 | 23.81 c ± 0.54 |
| E. multiflora + S. typhimurium | 97.67 a ± 0.18 | 63.99 b ± 0.61 | 58.51 b ± 2.5 | 26.45 c ± 0.99 |
| C. villosa + S. aureus | 97.55 a ± 0.44 | 77.19 b ± 3.34 | 54.04 c ± 1.17 | 31.27 d ± 0.31 |
| C. villosa + E. coli | 98.82 a ± 0.91 | 67.32 b ± 1.89 | 52.46 c ± 4.34 | 30.26 d ± 0.94 |
| C. villosa + L. monocytogenes | 98.43 a ± 0.51 | 72.55 b ± 4.61 | 51.49 c ± 0.90 | 28.05 d ± 1.01 |
| C. villosa + S. typhimurium | 97.55 a ± 1.51 | 83.27 b ± 1.07 | 58.89 c ± 0.66 | 41.27 d ± 0.85 |
| P. latifolia + S. aureus | 96.41 a ± 0.72 | 82.70 b ± 5.17 | 58.67 c ± 0.48 | 26.00 d ± 2.16 |
| P. latifolia + E. coli | 98.00 a ± 0.39 | 61.00 b ± 6.27 | 54.20 c ± 2.50 | 24.92 d ± 5.73 |
| P. latifolia + L. monocytogenes | 97.63 a ± 0.31 | 65.09 b ± 0.31 | 56.69 c ± 5.63 | 27.75 d ± 0.48 |
| P. latifolia + S. typhimurium | 97.26 a ± 0.82 | 64.97 b ± 0.42 | 55.88 c ± 1.11 | 25.01 d ± 0.51 |
| Growth of Strains (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | P. lentiscus | R. offinalis | E. mutiflora | C. villosa | P. latifolia | |||||||
| Strains | 2.5 | 5 | 2.5 | 5 | 2.5 | 5 | 2.5 | 5 | 2.5 | 5 | 2.5 | 5 |
| S. aureus | 100 a | 100 a | 0.24 b | 0.06 d | 0.22 b | 0 | 0.23 b | 0 | 0.25 b | 0.14 c | 0.23 b | 0.18 b |
| E. coli | 100 a | 100 a | 0.11 c | 0 | 0.23 b | 0 | 0.23 b | 0 | 0.25 b | 0.1 b | 0.23 b | 0 |
| L. monocytogenes | 100 a | 100 a | 0.13 c | 0.03 cd | 0.17 bc | 0 | 0.14 c | 0.06 b | 0.13 c | 0.04 c | 0.22 b | 0.04 c |
| S. typhimurium | 100 a | 100 a | 0.22 b | 0.06 d | 0.10 d | 0.08 d | 0.13 c | 0.08 d | 0.25 b | 0.19 b | 0.19 b | 0.10 c |
| Species | Major Compound | Concentrations (mg/g) | References |
|---|---|---|---|
| Pistacia lentiscus | Chlorogenic acid | 17.4 ± 1.9 | [10,11] |
| 3,4,5 Tri-O-galloyquinic acid | 15.9 ± 0.2 | ||
| Rutin | 13.6 ± 1.5 | ||
| Phillyrea latifolia | Tyrosol | 78.2 ± 18.4 | [11] |
| Quercetin-7-O-rutinoside | 42.5 ± 5.1 | ||
| Oleuropein | 167.0 ± 7.7 | ||
| Erica multiflora | Quercetin | 43.57 ± 0.18 | [9] |
| Rutin | 42.05 ± 2.67 | ||
| Catechin | 27.29 ± 2.73 | ||
| Rosmarinus officinalis | Rosmarinic acid | 38.5 ± 0.04 | [12,34] |
| Carnosic acid | 26.4 ± 0.15 | ||
| Chlorogenic acid | 11.2 ± 0.17 | ||
| Calicotome villosa | vanillic acid | 1.4 | [33] |
| 4-hydroxybenzoic acid | 0.39 | ||
| chlorogenic acid | 0.39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nefzi, K.; Ben Jemaa, M.; Baraket, M.; Dakhlaoui, S.; Msaada, K.; Nasr, Z. In Vitro Antioxidant, Antibacterial and Mechanisms of Action of Ethanolic Extracts of Five Tunisian Plants against Bacteria. Appl. Sci. 2022, 12, 5038. https://doi.org/10.3390/app12105038
Nefzi K, Ben Jemaa M, Baraket M, Dakhlaoui S, Msaada K, Nasr Z. In Vitro Antioxidant, Antibacterial and Mechanisms of Action of Ethanolic Extracts of Five Tunisian Plants against Bacteria. Applied Sciences. 2022; 12(10):5038. https://doi.org/10.3390/app12105038
Chicago/Turabian StyleNefzi, Khaoula, Mariem Ben Jemaa, Mokhtar Baraket, Sarra Dakhlaoui, Kamel Msaada, and Zouheir Nasr. 2022. "In Vitro Antioxidant, Antibacterial and Mechanisms of Action of Ethanolic Extracts of Five Tunisian Plants against Bacteria" Applied Sciences 12, no. 10: 5038. https://doi.org/10.3390/app12105038
APA StyleNefzi, K., Ben Jemaa, M., Baraket, M., Dakhlaoui, S., Msaada, K., & Nasr, Z. (2022). In Vitro Antioxidant, Antibacterial and Mechanisms of Action of Ethanolic Extracts of Five Tunisian Plants against Bacteria. Applied Sciences, 12(10), 5038. https://doi.org/10.3390/app12105038