
Quantifying Nutrient Content in the Leaves of Cowpea Using Remote Sensing

 ,
,  , , , , , , and
, , , , , , and 
Abstract
1. Introduction
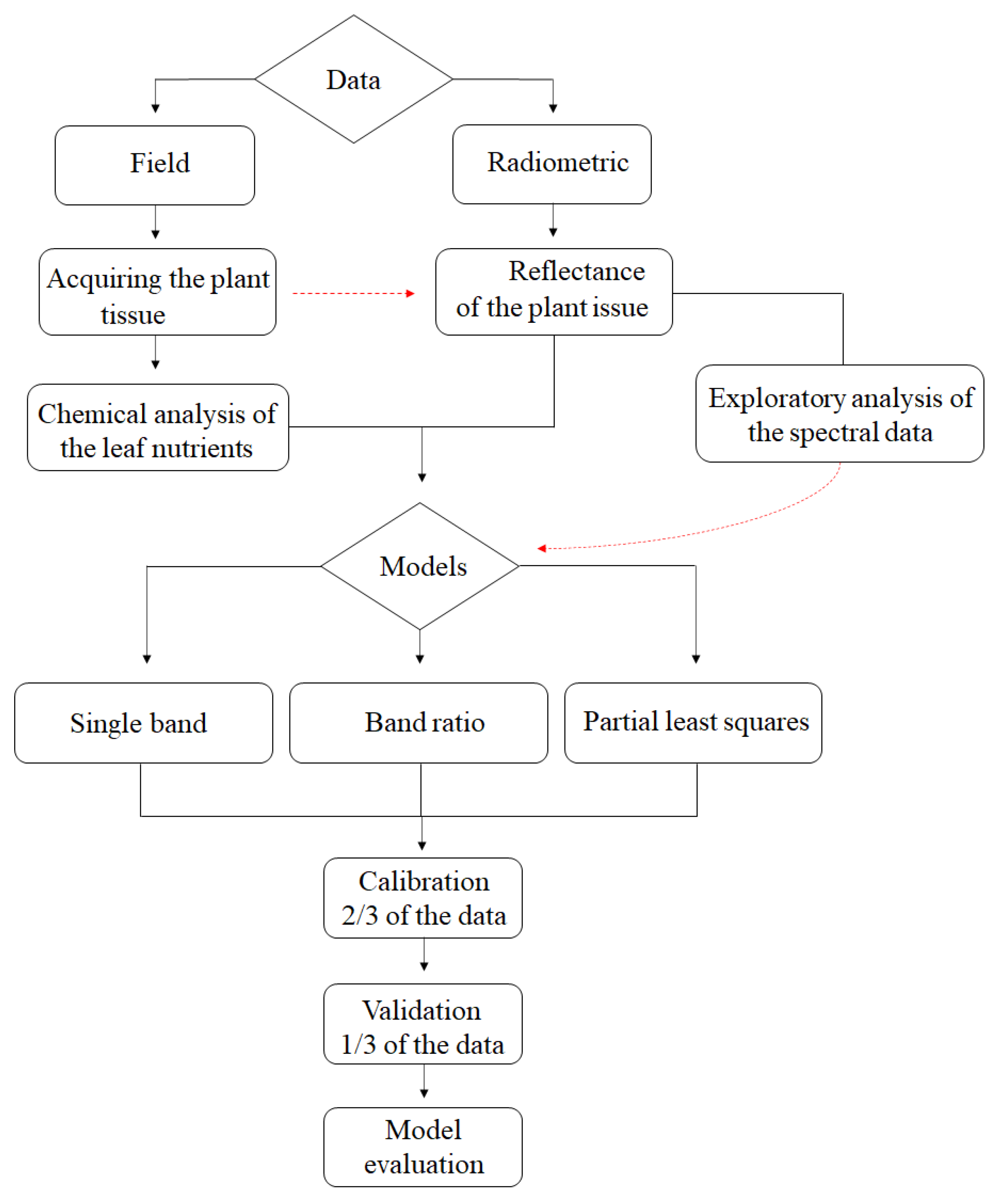
2. Materials and Methods
- FRBλ = bidirectional reflectance factor (dimensionless)
- La,λ = spectral radiance of the target (W cm−2 sr−1 μm−1)
- Lr,λ = spectral radiance of the reference plate (W cm−2 sr−1 μm−1).
- Yi = measured values
- Y = predicted values
- n = sample size
3. Results and Discussion
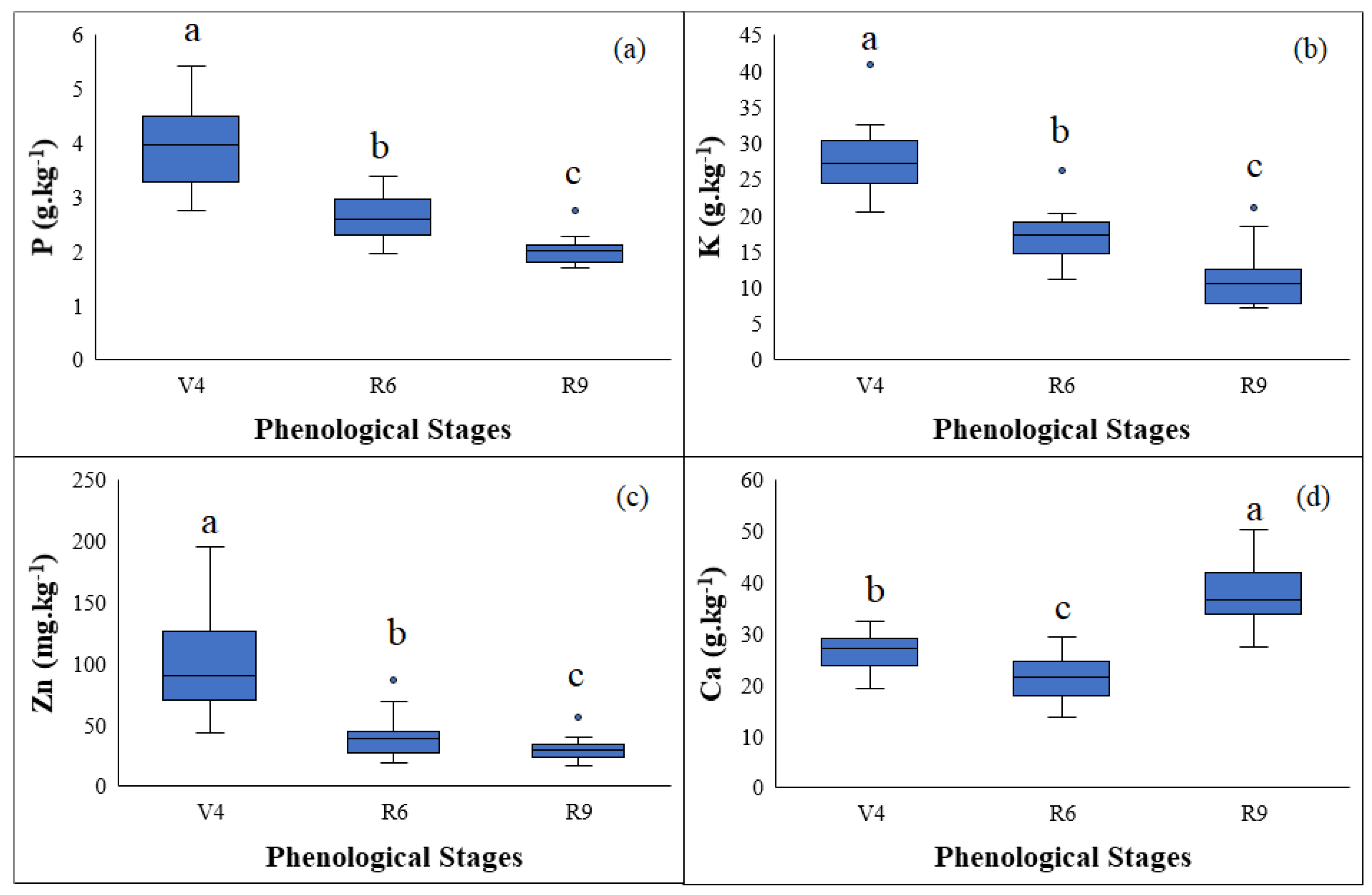
3.1. Nutrient Content
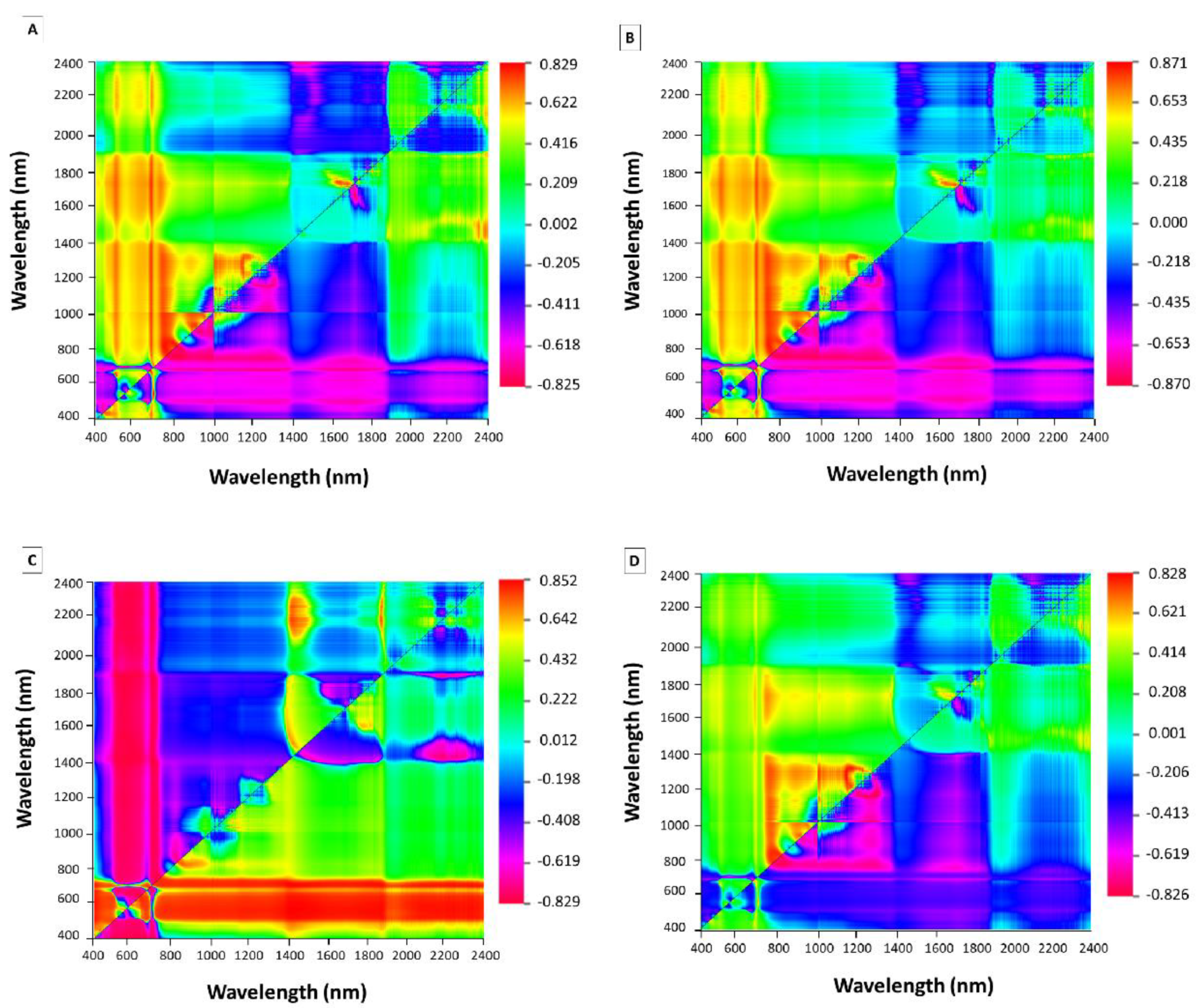
3.2. Spectral Reflectance
3.3. Model Calibration
3.3.1. Single Band
3.3.2. Band Ratio
3.3.3. Partial Least Squares Regression (PLSR)
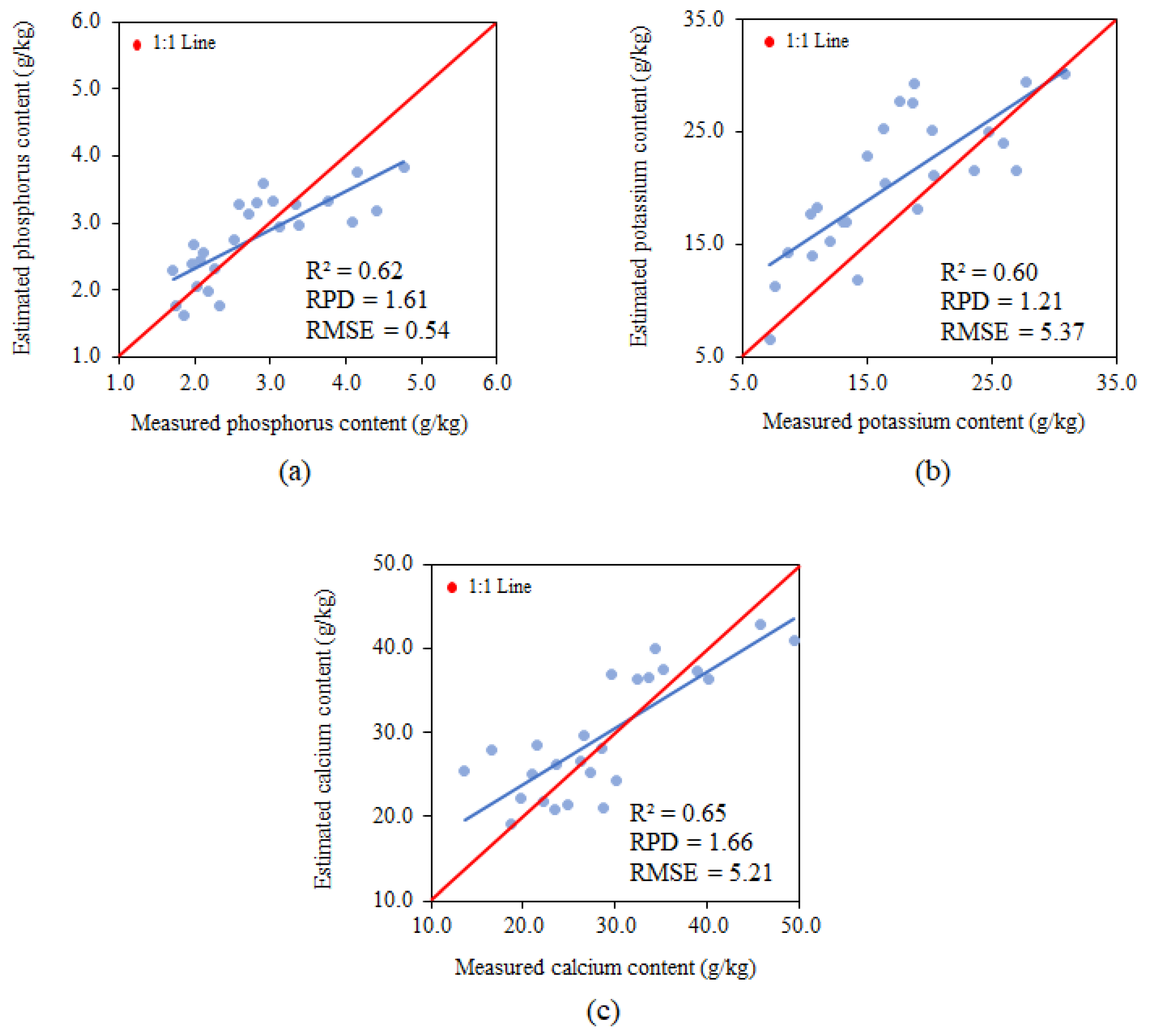
3.4. Model Validation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muñoz-Amatriaín, M.; Mirebrahim, H.; Xu, P.; Wanamaker, S.I.; Luo, M.; AlHakami, H.; Alpert, M.; Atokple, I.; Batieno, B.J.; Boukar, O.; et al. Genome resources for climate-resilient cowpea, an essential crop for food security. Plant J. 2017, 89, 1042–1054. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, M.; Lino-Neto, T.; Rosa, E.; Carnide, V. Cowpea: A legume crop for a challenging environment. J. Sci. Food Agric. 2017, 97, 4273–4284. [Google Scholar] [CrossRef] [PubMed]
- Lonardi, S.; Muñoz-Amatriaín, M.; Liang, Q.; Shu, S.; Wanamaker, S.I.; Lo, S.; Tanskanen, J.; Schulman, A.H.; Zhu, T.; Luo, M.; et al. The genome of cowpea (Vigna unguiculata L. Walp.). Plant J. 2019, 98, 767–782. [Google Scholar] [CrossRef]
- Freire Filho, F.R. Feijão-Caupi No Brasil: Produção, Melhoramento Genético, Avanços e Desafios; Embrapa Meio-Norte: Teresina, Brazil, 2011; 84p. [Google Scholar]
- Andrade Júnior, A.S.D.E. Cultivo do Feijão-Caupi (Vigna unguiculata (L.) Walp.); Embrapa Mid-North: Brasília, Brazil, 2002. [Google Scholar]
- Bertini, C.H.; Do Vale, J.C.; Borém, A. Feijão-caupi do Plantio à Colheita; Editora UFV: Viçosa, Brazil, 2017. [Google Scholar]
- Campanharo, M.; Monnerat, P.H.; Espindula, M.C.; Rabello, D.S. Doses De Níquel Em Feijão Caupi Cultivado Em Dois Solos. Rev. Caatinga 2013, 26, 9. [Google Scholar]
- Li, Q.; Jin, C.; Wang, G.; Ji, J.; Guan, C.; Li, X. Enhancement of endogenous SA accumulation improves poor-nutrition stress tolerance in transgenic tobacco plants overexpressing a SA-binding protein gene. Plant Sci. 2020, 292, 110384. [Google Scholar] [CrossRef]
- Malmir, M.; Tahmasbian, I.; Xu, Z.; Farrar, M.B.; Bai, S.H. Prediction of macronutrients in plant leaves using chemometric analysis and wavelength selection. J. Soils Sediments 2020, 20, 249–259. [Google Scholar] [CrossRef]
- Osco, L.P.; Ramos, A.P.M.; Moriya, É.A.S.; de Souza, M.; Junior, J.M.; Matsubara, E.; Imai, N.N.; Creste, J.E. Improvement of leaf nitrogen content inference in Valencia-orange trees applying spectral analysis algorithms in UAV mounted-sensor images. Int. J. Appl. Earth Obs. Geoinf. 2019, 83, 101907. [Google Scholar] [CrossRef]
- Abdulridha, J.; Batuman, O.; Ampatzidis, Y. UAV-Based Remote Sensing Technique to Detect Citrus Canker Disease Utilizing Hyperspectral Imaging and Machine Learning. Remote Sens. 2019, 11, 1373. [Google Scholar] [CrossRef]
- Calou, V.B.C.; Teixeira, A.D.S.; Moreira, L.C.J.; Lima, C.S.; de Oliveira, J.B.; de Oliveira, M.R.R. The use of UAVs in monitoring yellow sigatoka in banana. Biosyst. Eng. 2020, 193, 115–125. [Google Scholar] [CrossRef]
- Osco, L.P.; Ramos, A.P.M.; Pinheiro, M.M.F.; Moriya, É.A.S.; Imai, N.N.; Estrabis, N.; Ianczyk, F.; De Araújo, F.F.; Liesenberg, V.; Jorge, L.A.D.C.; et al. A Machine Learning Framework to Predict Nutrient Content in Valencia-Orange Leaf Hyperspectral Measurements. Remote Sens. 2020, 12, 906. [Google Scholar] [CrossRef]
- Peng, Z.; Guan, Z.; Liao, Y.; Lian, S. Estimating total leaf chlorophyll content of Gannan navel orange leaves using hyper-spectral data based on partial least squares regression. IEEE Access 2019, 7, 155540–155551. [Google Scholar] [CrossRef]
- Zheng, H.; Li, W.; Jiang, J.; Liu, Y.; Cheng, T.; Tian, Y.; Zhu, Y.; Cao, W.; Zhang, Y.; Yao, X. A Comparative Assessment of Different Modeling Algorithms for Estimating Leaf Nitrogen Content in Winter Wheat Using Multispectral Images from an Unmanned Aerial Vehicle. Remote Sens. 2018, 10, 2026. [Google Scholar] [CrossRef]
- O’Connell, J.; Byrd, K.; Kelly, M. Remotely-Sensed Indicators of N-Related Biomass Allocation in Schoenoplectus acutus. PLoS ONE 2014, 9, e90870. [Google Scholar] [CrossRef]
- Oliveira, M.; Queiroz, T.R.G.; Teixeira, A.D.S.; Moreira, L.C.J.; Leão, R.A.D.O. Reflectance spectrometry applied to the analysis of nitrogen and potassium deficiency in cotton. Rev. Ciência Agronômica 2020, 51. [Google Scholar] [CrossRef]
- Chen, Z.; Wang, X. Model for estimation of total nitrogen content in sandalwood leaves based on nonlinear mixed effects and dummy variables using multispectral images. Chemom. Intell. Lab. Syst. 2019, 195, 103874. [Google Scholar] [CrossRef]
- Fletcher, R.S.; Reddy, K. Random forest and leaf multispectral reflectance data to differentiate three soybean varieties from two pigweeds. Comput. Electron. Agric. 2016, 128, 199–206. [Google Scholar] [CrossRef]
- Rokhafrouz, M.; Latifi, H.; Abkar, A.A.; Wojciechowski, T.; Czechlowski, M.; Naieni, A.S.; Maghsoudi, Y.; Niedbała, G. Simplified and Hybrid Remote Sensing-Based Delineation of Management Zones for Nitrogen Variable Rate Application in Wheat. Agriculture 2021, 11, 1104. [Google Scholar] [CrossRef]
- Pullanagari, R.; Kereszturi, G.; Yule, I. Mapping of macro and micro nutrients of mixed pastures using airborne AisaFENIX hyperspectral imagery. ISPRS J. Photogramm. Remote Sens. 2016, 117, 1–10. [Google Scholar] [CrossRef]
- Wen, P.-F.; He, J.; Ning, F.; Wang, R.; Zhang, Y.-H.; Li, J. Estimating leaf nitrogen concentration considering unsynchronized maize growth stages with canopy hyperspectral technique. Ecol. Indic. 2019, 107, 105590. [Google Scholar] [CrossRef]
- Yu, X.; Lu, H.; Liu, Q. Deep-learning-based regression model and hyperspectral imaging for rapid detection of nitrogen concentration in oilseed rape (Brassica napus L.) leaf. Chemom. Intell. Lab. Syst. 2018, 172, 188–193. [Google Scholar] [CrossRef]
- De Oliveira, D.M.; Fontes, L.M.; Pasquini, C. Comparing laser induced breakdown spectroscopy, near infrared spectroscopy, and their integration for simultaneous multi-elemental determination of micro- and macronutrients in vegetable samples. Anal. Chim. Acta 2019, 1062, 28–36. [Google Scholar] [CrossRef]
- Ling, B.; Goodin, D.G.; Raynor, E.; Joern, A. Hyperspectral Analysis of Leaf Pigments and Nutritional Elements in Tallgrass Prairie Vegetation. Front. Plant Sci. 2019, 10, 142. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Rahman, E.M.; Mutanga, O.; Odindi, J.; Adam, E.; Odindo, A.; Ismail, R. Estimating Swiss chard foliar macro- and micronutrient concentrations under different irrigation water sources using ground-based hyperspectral data and four partial least squares (PLS)-based (PLS1, PLS2, SPLS1 and SPLS2) regression algorithms. Comput. Electron. Agric. 2017, 132, 21–33. [Google Scholar] [CrossRef]
- Mao, H.; Gao, H.; Zhang, X.; Kumi, F. Nondestructive measurement of total nitrogen in lettuce by integrating spectroscopy and computer vision. Sci. Hortic. 2015, 184, 1–7. [Google Scholar] [CrossRef]
- Gama, T.; Wallace, H.; Trueman, S.; Tahmasbian, I.; Bai, S. Hyperspectral imaging for non-destructive prediction of total nitrogen concentration in almond kernels. Acta Hortic. 2018, 1219, 259–264. [Google Scholar] [CrossRef]
- González-Martín, I.; Hernández-Hierro, J.M.; González-Cabrera, J.M. Use of NIRS technology with a remote reflectance fibre-optic probe for predicting mineral composition (Ca, K, P, Fe, Mn, Na, Zn), protein and moisture in alfalfa. Anal. Bioanal. Chem. 2007, 387, 2199–2205. [Google Scholar] [CrossRef]
- Galvez-Sola, L.; Garciasanchez, F.J.; Pérez-Pérez, J.G.; Gimeno, V.; Navarro, J.M.; Moral, R.; Nicolas, J.J.M.; Nieves, M. Rapid estimation of nutritional elements on citrus leaves by near infrared reflectance spectroscopy. Front. Plant Sci. 2015, 6, 571. [Google Scholar] [CrossRef]
- Halgerson, J.; Sheaffer, C.; Martin, N.; Peterson, P.; Weston, S. Nearinfrared reflectance spectroscopy prediction of leaf and mineral concentrations in alfalfa. Agron. J. 2004, 96, 344–351. [Google Scholar]
- Yarce, C.J.; Rojas, G. Near infrared spectroscopy for the analysis of macro and micro nutrients in sugarcane leaves. Sugar Ind. 2012, 137, 707–710. [Google Scholar] [CrossRef]
- Köppen, W. Climatologia: Con un Estudio de los Climas de la Tierra; Fondo de Cultura Economica: Mexico City, Mexico, 1948; 478p. [Google Scholar]
- Cardoso, M.J.; Bastos, E.A.; Andrade Junior, A.S.; De Athayde Sobrinho, C. Feijão-Caupi: O Produtor Pergunta, a Embrapa Responde; Embrapa: Brasília, Brazil, 2017; 244p. [Google Scholar]
- Aquino, A.B.; Aquino, B.F.; Hernandez, F.F.F.; Holanda, F.J.M.; Freire, J.M.; Crisóstomo, L.A.; Costa, R.I.; Unchôa, S.C.P.; Fernandes, V.L.B. Recomendações de Adubação e Calagem para o Estado do Ceará; UFC: Fortaleza, Brazil, 1993; 247p. [Google Scholar]
- Furtini Neto, A.E.; Vale, F.R.; Resende, A.V.; Guilherme, L.R.G.; Guedes, G.A.A. Fertilidade do Solo; UFLA: Lavras, Brazil, 2001; 261p. [Google Scholar]
- SILVA, C.A. Uso de resíduos orgânicos na agricultura. In Fundamentos da Matéria Orgânica do solo: Ecossistemas Tropicais e Subtropicais, 2nd ed.; Métropole: Porto Alegre, Brazil, 2008; pp. 597–624. [Google Scholar]
- Magalhães, A.C.M. de Adubação orgânica Com Base na taxa de Mineralizaçãode Nutrientes do Composto Orgânico. Master’s Thesis, Universidade Federal do Ceará, Fortaleza, Brazil, 2018. [Google Scholar]
- Dourado Neto, D.; Fancelli, A.L. Produção de Feijão; Agropecuária: Guaíba, Brazil, 2000; 385p. [Google Scholar]
- Campelo, D.H. Uso do Sensoriamento Remoto Para Diagnóstico Nutricional na Cultura do Milho Irrigado. Ph.D. Thesis, Universidade Federal do Ceará, Fortaleza, Brazil, 2018; 182p. [Google Scholar]
- Silva, F.C. da (Org.). Manual de Análises Químicas de Solos, Plantas e Fertilizante; Embrapa Comunicação para Transferência de Tecnologia: Brasília, Brazil, 1999. [Google Scholar]
- Ogashawara, I.; Curtarelli, M.P.; Souza, A.F.; Augusto-Silva, P.B.; Alcântara, E.H.; Stech, J.L. Interactive Correlation Environment (ICE)—A Statistical Web Tool for Data Collinearity Analysis. Remote Sens. 2014, 6, 3059–3074. [Google Scholar] [CrossRef]
- Lopes, F.B.; Barbosa, C.C.F.; Novo, E.M.L.D.M.; de Carvalho, L.A.S.; de Andrade, E.M.; Teixeira, A.D.S. Modelling chlorophyll-a concentrations in a continental aquatic ecosystem of the Brazilian semi-arid region based on remote sensing. Revista Ciência Agronômica 2021, 52, 1–12. [Google Scholar] [CrossRef]
- Wilcox, G.E.; Fageria, N.K. Deficiências Nutricionais do feijão, sua Identificação e Correção; Boletim Técnico, 5; Embrapa/CNPAF: Goiânia, Brazil, 1976; 22p. [Google Scholar]
- Martinez, H.E.P.; Menezes, J.F.S.; De Souza, R.B.; Venegas, V.H.A.; Guimarães, P.T.G. Faixas críticas de concentrações de nutrientes e avaliação do estado nutricional de cafeeiros em quatro regiões de Minas Gerais. Pesqui. Agropecuária Bras. 2003, 38, 703–713. [Google Scholar] [CrossRef]
- Marschner, P. Marschner’s Mineral Nutrititon of Higher Plants, 3rd ed.; Academic Press: Cambridge, MA, USA, 2012; 649p. [Google Scholar]
- Büll, L.T.; Novello, A.; Corrêa, J.; Boas, R. Doses de fósforo e zinco na cultura do alho em condições de casa de vegetação; Bragantia: Campinas, Brazil, 2008; Volume 67, pp. 941–949. [Google Scholar]
- Zucareli, C. Adubação fosfatada, produção e desempenho em campo de sementes de feijoeiro CV. Carioca Precoce e IAC Carioca Tybatã. Master’s Thesis, Universidade Estadual Paulista, Botucatu, Brazil, 2005. Júlio de Mesquita Filho. [Google Scholar]
- Cavalcante, L.F.; Santos, C.J.O.; De Holanda, J.S.; De Lima Neto, A.J.; De Souto, A.G.L.; Dantas, T.A.G. Produção de maracujazeiro Amarelo no solo Com Calcário e Potássio sob Irrigação com água Salina; Irriga: Botucatu, Brazil, 2018; Volume 23, pp. 727–740. [Google Scholar]
- Malavolta, E. Manual de Nutrição Mineral de Plantas; Editora Agronômica Ceres: São Paulo, Brazil, 2006; p. 638. [Google Scholar]
- Pedrosa, M.V.B. Qualidade Fisiológica de Sementes de Feijão (Stageolus vulgaris L.) em Função da Maturação e Adubação Com enxofre, Nitrogênio e Zinco. Master’s Thesis, Universidade Federal do Espírito Santo, Vitória, Brazil, 2017; p. 87. [Google Scholar]
- Ponzoni, F.J.; Shimbukuro, Y.E.; Kuplich, T.M. Sensoriamento Remoto da Vegetação. São José dos Campos, 2nd ed.; Oficina de Textos: Sao paulo, Brazil, 2012; 176p. [Google Scholar]
- Moreira, M.A. Fundamentos do Sensoriamento Remoto e Metodologias de Aplicação, 4th ed.; Viçosa, M.G., Ed.; UFV: Abbotsford, BC, Canada, 2011; 422p. [Google Scholar]
- Gasparotto, A.C.; Nanni, M.R.; Miotto, L.S.; Guirado, G.C.; Silva Junior, C.A.; Silva, A.A.; Cezar, E.; Romagnoli, F. Comportamento espectral de milho submetido a diferentes doses de nitrogênio. XVII Simpósio Brasileiro de Sensoriamento Remoto-SBSR, João Pessoa-PB, Brasil. 2015. Available online: http://marte2.sid.inpe.br/col/sid.inpe.br/marte2/2015/05.31.21.54/doc/@sumario.htm (accessed on 31 October 2021).
- Liu, L.; Song, B.; Zhang, S.; Liu, X. A Novel Principal Component Analysis Method for the Reconstruction of Leaf Reflectance Spectra and Retrieval of Leaf Biochemical Contents. Remote Sens. 2017, 9, 1113. [Google Scholar] [CrossRef]
- Carvalho, A. Estudo de Características foliares de Espécies de Lenhosas de cerrado e Sua relação com os Espectros de Reflectância. Ph.D. Thesis, Universidade de Brasília, Brasília, Brazil, 2005. [Google Scholar]
- Tierny, J.; Vandeborre, J.; Daoudi, M. The Visual Computer. Int. J. Comput. Graphicsv 2008, 24, 155–172. [Google Scholar]
- Chicati, M.S. Resposta Espectral da Cultura do Feijão e sua Relação com Parâmetros Biofísicos em Diferentes Doses de Nitrogênio/Mônica Sacioto Chicati. Master’s Thesis, Universidade Estadual de Maringá, Maringá, Brazil, 2015. [Google Scholar]
- Jensen, J.R. Sensoriamento Remoto do Ambiente: Uma Perspectiva em Recursos Terrestres; Parêntese: São José dos Campos, Brazil, 2011; 582p. [Google Scholar]
- Papa, R.D.A. Comportamento Espectro-Temporal Da Cultura Do Feijão, Por Meio De Dados Obtidos Por Espectroradiometria, Câmera Digital E Imagem Aster; Novas Edições Acadêmicas: Brasília, Brazil, 2009; 146p. [Google Scholar]
- Moreira, L.C.J.; Teixeira, A.D.S.; Galvao, L. Laboratory Salinization of Brazilian Alluvial Soils and the Spectral Effects of Gypsum. Remote Sens. 2014, 6, 2647–2663. [Google Scholar] [CrossRef]
- Silva Junior, M.B. Fertilizantes Foliares no Manejo da Mancha de Phoma de Cafeeiro. Master’s Thesis, Universidade Federal de Lavras, Lavras, Brazil, 2013. [Google Scholar]
- Taiz, L.; Zeiger, E. Fisiologia Vegetal; Artmed: Porto Alegre, Brazil, 2004; pp. 449–484. [Google Scholar]
- Stein, B.R.; Thomas, V.A.; Lorentz, L.J.; Strahm, B.D. Predicting macronutrient concentrations from loblolly pine leaf reflectance across local and regional scales. GIScience Remote Sens. 2014, 51, 269–287. [Google Scholar] [CrossRef]
- Gong, X.; Hong, M.; Wang, Y.; Zhou, M.; Cai, J.; Liu, C.; Gong, S.; Hong, F. Cerium Relieves the Inhibition of Photosynthesis of Maize Caused by Manganese Deficiency; Biological Trace Element Research: Clifton, FL, USA, 2011; Volume 141, pp. 305–316. [Google Scholar]
- Barbosa, M.; Silva, M.; Willadino, L.; Ulisses, C.; Camara, T. Geração e Desintoxicação Enzimática de Espécies Reativas de Oxigênio em Plantas; Ciência Rural: Santa Maria, FL, USA, 2014; Volume 44, pp. 453–460. [Google Scholar]
- Shao, Y.; He, Y. Visible/Near Infrared Spectroscopy and Chemometrics for the Prediction of Trace Element (Fe and Zn) Levels in Rice Leaf. Sensors 2013, 13, 1872–1883. [Google Scholar] [CrossRef] [PubMed]
- Curran, P.J.; Dungan, J.L.; Macler, F.A.; Plummer, S.E. The effect of a red leaf pigment on the relationship between red edge and chlorophyll concentration. Remote Sens. Environ. 1991, 35, 69–76. [Google Scholar] [CrossRef]
- Chu, X.; Guo, Y.; He, J.; Yao, X.; Zhu, Y.; Cao, W.; Cheng, T.; Tian, Y. Comparison of Different Hyperspectral Vegetation Indices for Estimating Canopy Leaf Nitrogen Accumulation in Rice. Agron. J. 2014, 106, 1911–1920. [Google Scholar] [CrossRef]
- Tian, Y.C.; Yao, X.; Cao, W.X.; Hannaway, D.B.; Zhu, Y. Assessing Newly Developed and Published Vegetation Indices for Estimating Rice Leaf Nitrogen Concentration with Ground- and Space-Based Hyperspectral Reflectance; Field Crops Research: Amsterdam, The Netherlands, 2011; Volume 120, pp. 299–310. [Google Scholar]
- Abdel-Rahman, E.M.; Ahmed, F.B.; Berg, M.V.D. Estimation of sugarcane leaf nitrogen concentration using in situ spectroscopy. Int. J. Appl. Earth Obs. Geoinf. Enschede 2010, 12, 52–57. [Google Scholar] [CrossRef]
- Zhou, X.; Huang, W.; Kong, W.; Ye, H.; Luo, J.; Chen, P. Remote estimation of canopy nitrogen content in winter wheat using airborne hyperspectral reflectance measurements. Adv. Space Res. 2016, 58, 1627–1637. [Google Scholar] [CrossRef]
- Mee, C.Y.; Bala, S.K.; Mohd, A.H. Detecting and Monitoring Plant Nutrient Stress Using Remote Sensing Approaches: A Review. Asian J. Plant Sci. 2016, 16, 1–8. [Google Scholar] [CrossRef]
- Prananto, J.A.; Minasny, B.; Weaver, T. Near Infrared (NIR) Spectroscopy as a Rapid and Cost-Effective Method for Nutrient Analysis of Plant Leaf Tissues, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020; Volume 164, ISBN 9780128207710. [Google Scholar]
- Clark, D.H.; Mayland, H.F.; Lamb, R.C. Mineral Analysis of Forages with near Infrared Reflectance Spectroscopy 1. Agron. J. 1987, 79, 485–490. [Google Scholar] [CrossRef]
- De Aldana, B.V.; Criado, B.G.; Ciudad, A.G.; Corona, M.P. Estimation of mineral content in natural grasslands by near infrared reflectance spectroscopy. Commun. Soil Sci. Plant Anal. 1995, 26, 1383–1396. [Google Scholar] [CrossRef][Green Version]
- Ge, Y.; Atefi, A.; Zhang, H.; Miao, C.; Ramamurthy, R.K.; Sigmon, B.; Yang, J.; Schnable, J.C. High-throughput analysis of leaf physiological and chemical traits with VIS–NIR–SWIR spectroscopy: A case study with a maize diversity panel. Plant Methods 2019, 15, 1–12. [Google Scholar] [CrossRef]
- Liao, H.; Wu, J.; Chen, W.; Guo, W.; Shi, C. Rapid diagnosis of nutrient elements in fingered citron leaf using near infrared reflectance spectroscopy. J. Plant Nutr. 2012, 35, 1725–1734. [Google Scholar] [CrossRef]
- Comino, F.; Ayora-Cañada, M.; Aranda, V.; Díaz, A.; Domínguez-Vidal, A. Near-infrared spectroscopy and X-ray fluorescence data fusion for olive leaf analysis and crop nutritional status determination. Talanta 2018, 188, 676–684. [Google Scholar] [CrossRef]
- Santoso, H.; Tani, H.; Wang, X.; Segah, H. Predicting oil palm leaf nutrient contents in kalimantan, indonesia by measuring reflectance with a spectroradiometer. Int. J. Remote Sens. 2018, 40, 7581–7602. [Google Scholar] [CrossRef]
- Petisco, C.; García-Criado, B.; de Aldana, B.R.V.; Zabalgogeazcoa, I.; Mediavilla, S.; García-Ciudad, A. Use of near-infrared reflectance spectroscopy in predicting nitrogen, phosphorus and calcium contents in heterogeneous woody plant species. Anal. Bioanal. Chem. 2005, 382, 458–465. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Attribute | Value |
|---|---|
| Coarse sand (g.kg−1) | 523.00 |
| Fine sand (g.kg−1) | 370.00 |
| Silt (g.kg−1) | 52.00 |
| Clay (g.kg−1) | 55.00 |
| Bulk density (g.cm−3) | 1.46 |
| Particle density (g.cm−3) | 2.68 |
| pH (water) | 6.92 |
| Calcium (mmolc.dm−3) | 16.00 |
| Magnesium (mmolc.dm−3) | 13.00 |
| Sodium (mmolc.dm−3) | 3.00 |
| Potassium (mmolc.dm−3) | 1.00 |
| H + Al (mmolc.dm−3) | 18.20 |
| Organic carbon (g.kg−1) | 7.02 |
| Total nitrogen (g.kg−1) | 0.68 |
| Organic matter (g.kg−1) | 12.10 |
| Available phosphorus (mg.dm−3) | 23.00 |
| C:N ratio | 10:1 |
| Nutrient | Wavelength (nm) | Equation | R2 |
|---|---|---|---|
| P | 684 | y = −89.297x + 8.158 | 0.62 |
| K | 684 | y = −915.51x + 74.546 | 0.63 |
| Ca | 720 | y = 208.12x − 41.659 | 0.67 |
| Nutrient | Band Ratio (nm) | Equation | R2 |
|---|---|---|---|
| P | 826/750 | P = 34.63x − 32.35 | 0.66 |
| K | 744/816 | K = −256.09x + 265.54 | 0.70 |
| Ca | 720/844 | Ca = 89.675x − 33.934 | 0.63 |
| Zn | 1252/1148 | Zn = 6818.2x − 6643 | 0.66 |
| Phenological Stage | Nutrient | Band | n | Adjusted R2 |
|---|---|---|---|---|
| V4 | Ca | 413, 625, 1301, 1405, 1683, 1714, 1726, 1727, 1883, 1904 | 16 | 0.98 |
| Zn | 634, 1364, 1981, 1994 | 16 | 0.73 | |
| R6 | Zn | 923, 925, 944, 951, 1864, 1890 | 17 | 0.90 |
| R9 | K | 630, 1405 | 17 | 0.43 |
| Ca | 613, 631, 1350 | 17 | 0.59 | |
| All (V4, R6, R9) | P | 504, 651, 685, 1649, 1715 | 50 | 0.77 |
| K | 504, 651, 685, 1715, 1735 | 50 | 0.84 | |
| Ca | 705, 1890, 2377 | 50 | 0.73 | |
| Zn | 751, 1715, 2434 | 50 | 0.57 |
| Phenological Stage | Nutrient | Equation |
|---|---|---|
| V4 | Ca | Ca = 41.380740 − 10.38684 λ1683 nm − 3.63733 λ1405 nm + 1.344476 λ413 nm + 4.155283 λ1883 nm − 9.864286 λ1726 nm − 7.38664·λ1714 nm − 7.014894 λ1031 nm + 5.896256 λ1904 nm − 10.20575 λ1727 nm + 2.19506·λ625 nm |
| Zn | Zn = −607.617 − 10181.2 λ634 nm + 3809.383 λ1364 nm + 2204.323 λ1994 nm − 3493.65 λ1981 nm | |
| R6 | Zn | Zn = −119.164400 + 70.25752 λ944 nm + 71.05573 λ951 nm + 71.56557 λ1890 nm + 88.90969 λ1864 nm + 68.28416 λ925 nm + 67.71706 λ923 nm |
| R9 | K | K = −1.65418 − 80.7834 λ630 nm + 116.8171 λ1405 nm |
| Ca | Ca = 106.346000 − 252.393 λ1350 nm + 3637.852 λ631 nm − 3048.85 λ613 nm | |
| All the bands together (V4. R6. R9) | P | P = 6.162487 − 124.4690 λ685 nm + 181.6616 λ651 nm − 87.4141 λ504 nm + 299.9536 λ1715 nm − 293.5019 λ1649 nm |
| K | K = 8.754299 − 427.4241 λ685 nm + 2012.5704 λ651 nm − 1940.1794 λ504 nm + 2668.3838 λ1715 nm − 2624.0833λ1735 nm | |
| Ca | Ca = 16.96427 + 193.3209 λ705 nm − 670.447 λ2377 nm + 458.5386 λ1890 nm | |
| Zn | Zn = −92.4856 + 1803.0171 λ1715 nm − 2073.2190 λ2434 nm − 519.7374 λ751 nm |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amaral, J.B.C.; Lopes, F.B.; Magalhães, A.C.M.d.; Kujawa, S.; Taniguchi, C.A.K.; Teixeira, A.d.S.; Lacerda, C.F.d.; Queiroz, T.R.G.; Andrade, E.M.d.; Araújo, I.C.d.S.; et al. Quantifying Nutrient Content in the Leaves of Cowpea Using Remote Sensing. Appl. Sci. 2022, 12, 458. https://doi.org/10.3390/app12010458
Amaral JBC, Lopes FB, Magalhães ACMd, Kujawa S, Taniguchi CAK, Teixeira AdS, Lacerda CFd, Queiroz TRG, Andrade EMd, Araújo ICdS, et al. Quantifying Nutrient Content in the Leaves of Cowpea Using Remote Sensing. Applied Sciences. 2022; 12(1):458. https://doi.org/10.3390/app12010458
Chicago/Turabian StyleAmaral, Julyanne Braga Cruz, Fernando Bezerra Lopes, Ana Caroline Messias de Magalhães, Sebastian Kujawa, Carlos Alberto Kenji Taniguchi, Adunias dos Santos Teixeira, Claudivan Feitosa de Lacerda, Thales Rafael Guimarães Queiroz, Eunice Maia de Andrade, Isabel Cristina da Silva Araújo, and et al. 2022. "Quantifying Nutrient Content in the Leaves of Cowpea Using Remote Sensing" Applied Sciences 12, no. 1: 458. https://doi.org/10.3390/app12010458
APA StyleAmaral, J. B. C., Lopes, F. B., Magalhães, A. C. M. d., Kujawa, S., Taniguchi, C. A. K., Teixeira, A. d. S., Lacerda, C. F. d., Queiroz, T. R. G., Andrade, E. M. d., Araújo, I. C. d. S., & Niedbała, G. (2022). Quantifying Nutrient Content in the Leaves of Cowpea Using Remote Sensing. Applied Sciences, 12(1), 458. https://doi.org/10.3390/app12010458