
Echinacea purpurea Alleviates Cyclophosphamide-Induced Immunosuppression in Mice

 ,
, 
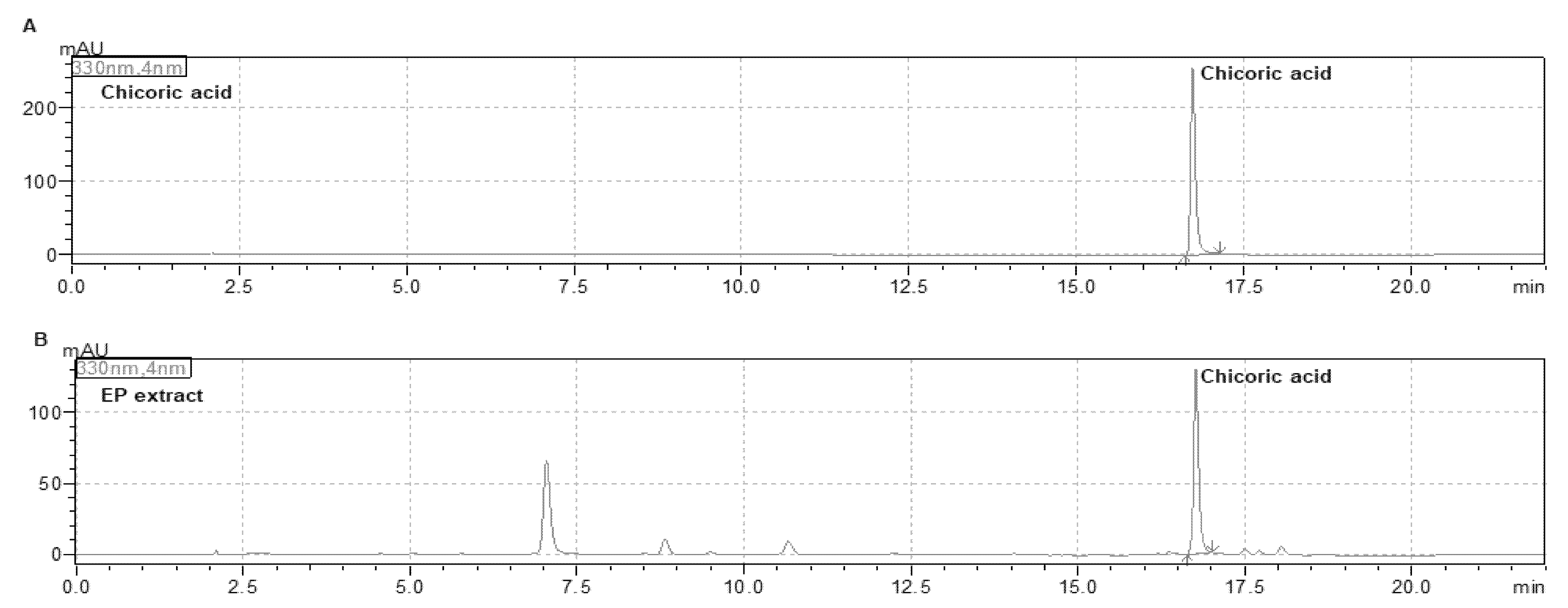{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Animals
2.3. Preparation of EP Extract and High-Performance Liquid Chromatography (HPLC) Analysis
2.4. CP-Induced Immunosuppression in Mice
2.5. Immune Organ Indices
2.6. Preparation of Spleen Cell Suspension
2.7. Lymphocyte Proliferation Assay
2.8. Splenic Natural Killer (NK) Cell Activity
2.9. Splenic T-Lymphocyte Subpopulation Assay
2.10. Determination of Serum Cytokines
2.11. RNA Extraction and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.12. Statistical Analysis
3. Results
3.1. Fingerprint of EP Extract
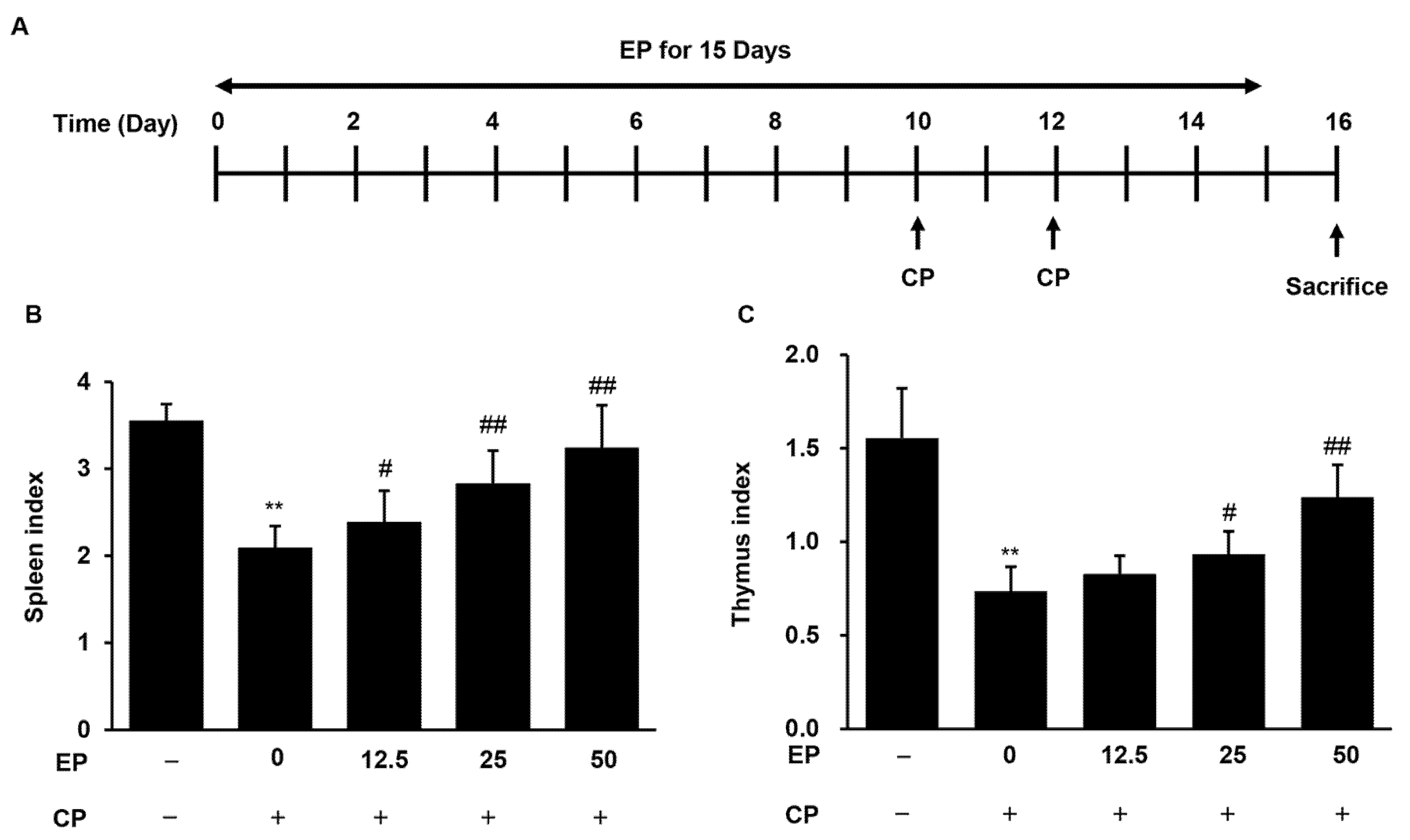
3.2. Effects of EP on Spleen and Thymus Indexes in the CP-Induced Immunosuppression Model
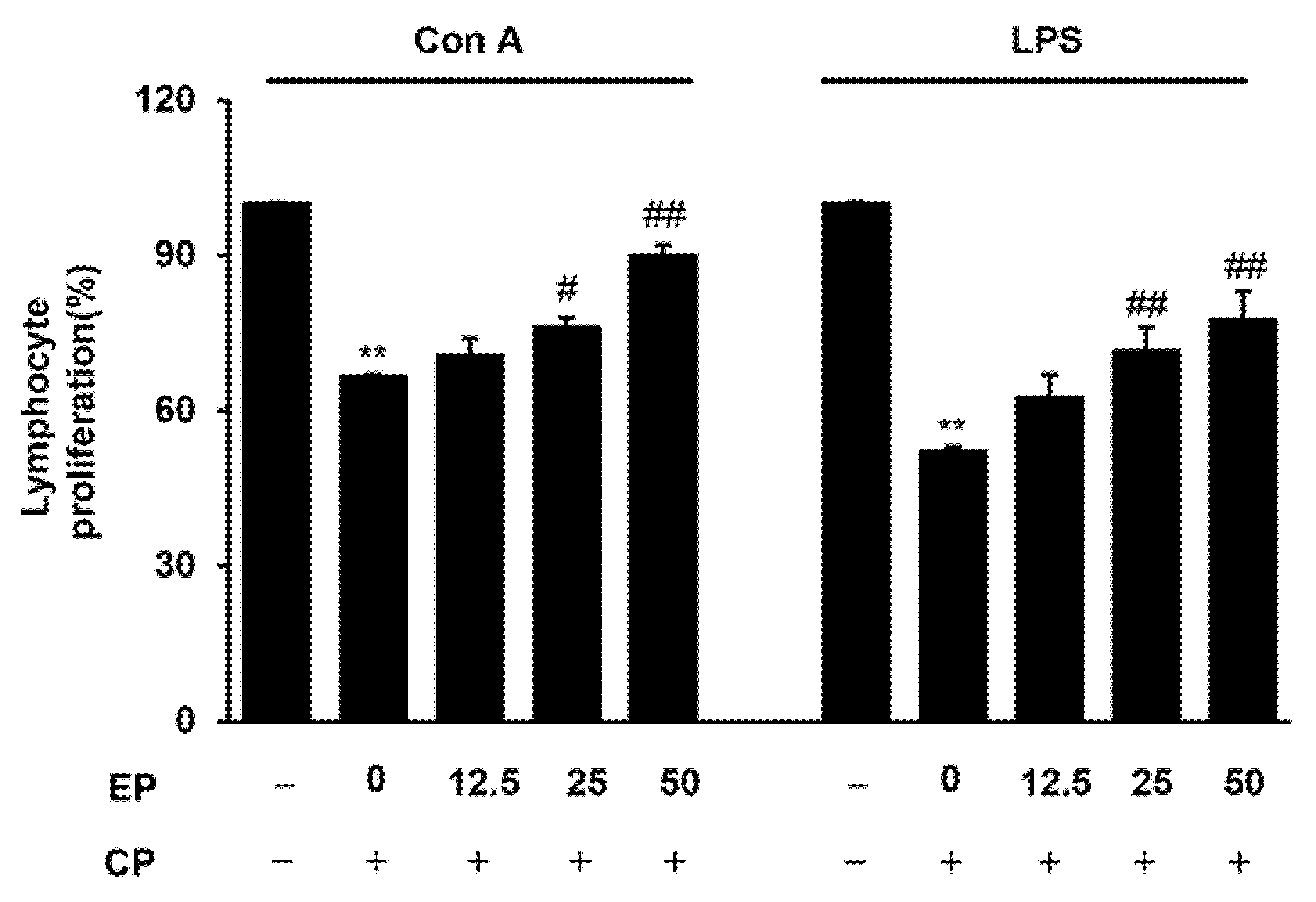
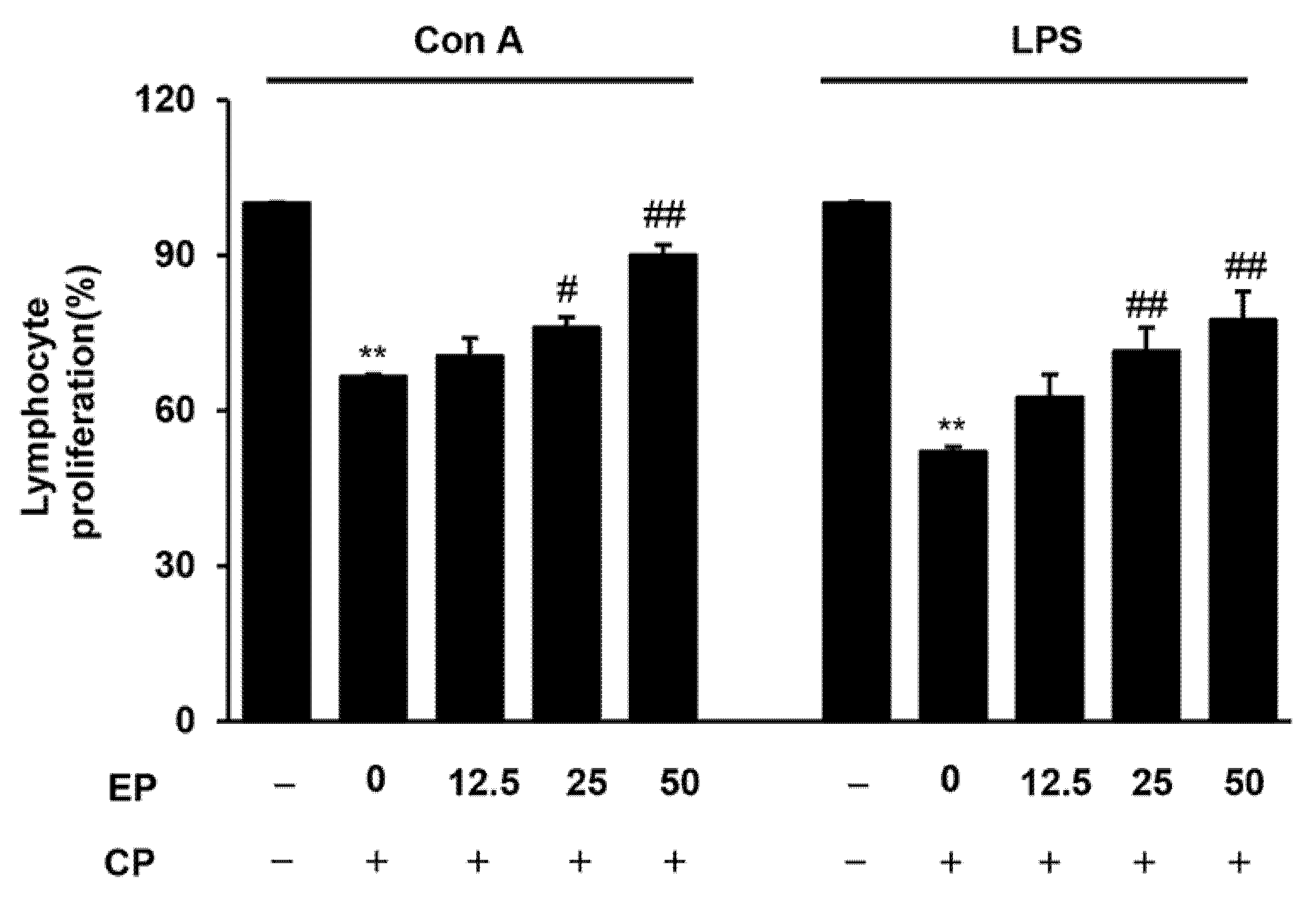
3.3. Effect of EP on Splenic Lymphocyte Proliferation in the CP-Induced Immunosuppression Model
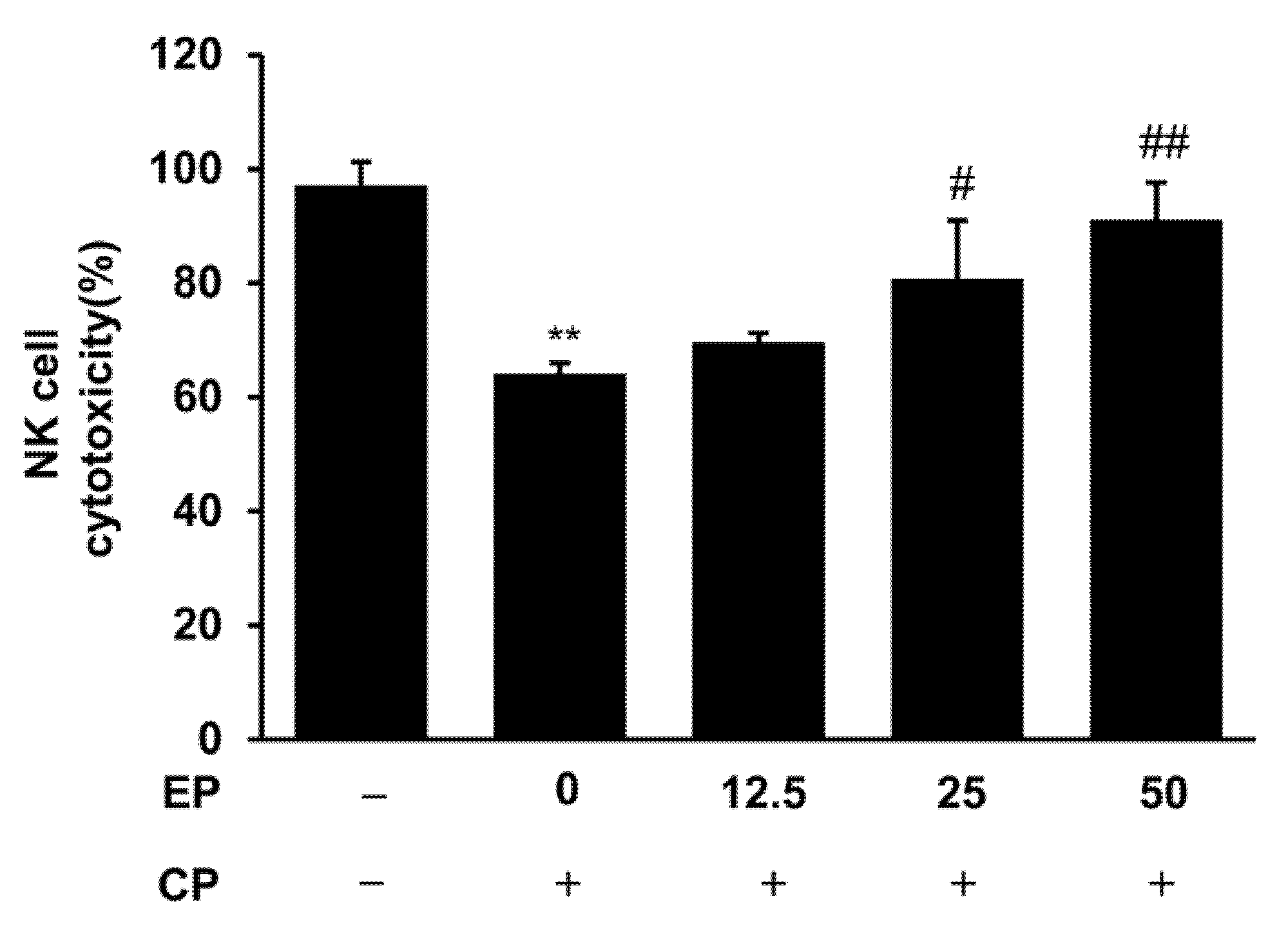
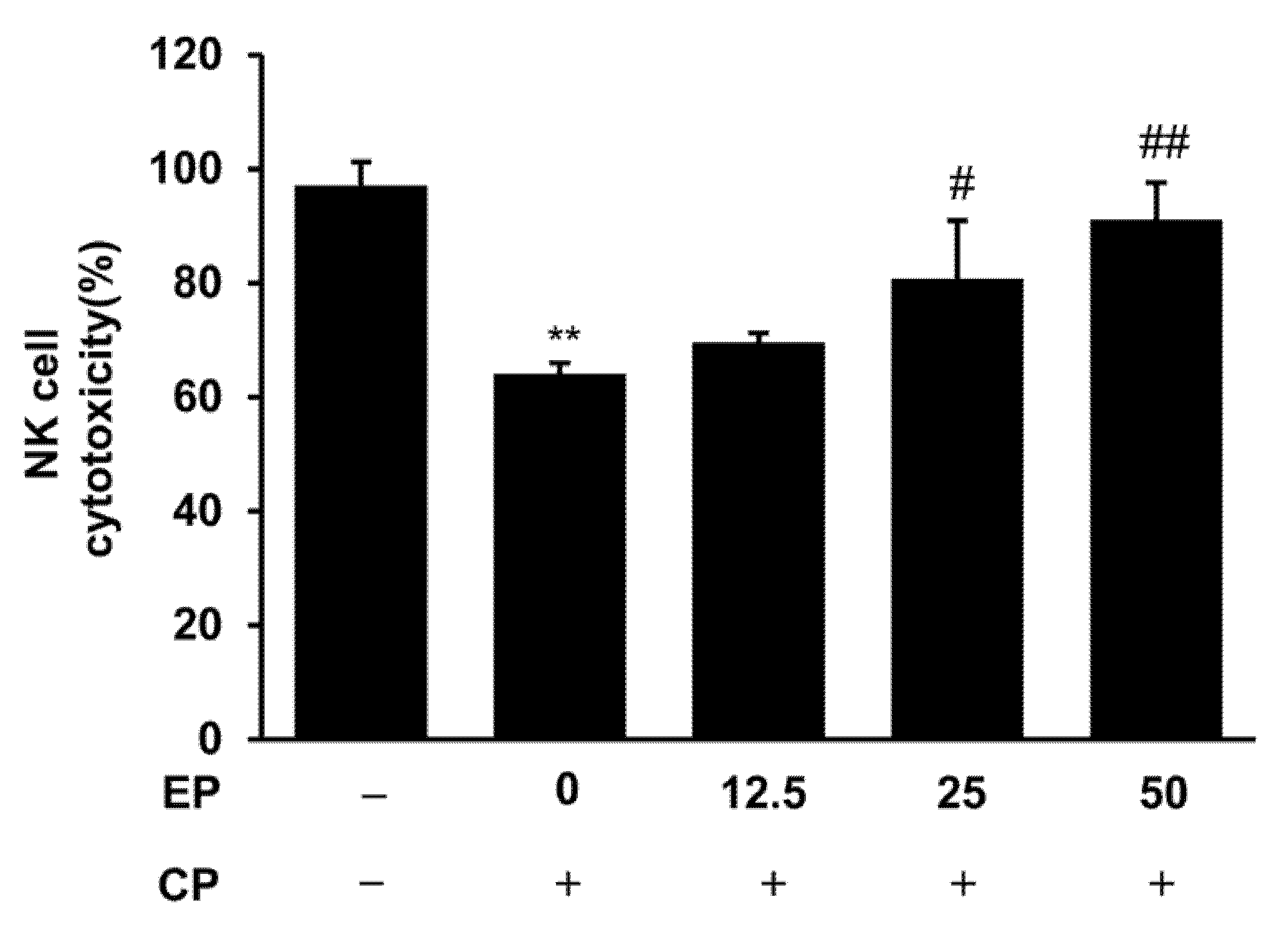
3.4. Effect of EP on Splenic NK Cell Activity in the CP-Induced Immunosuppression Model
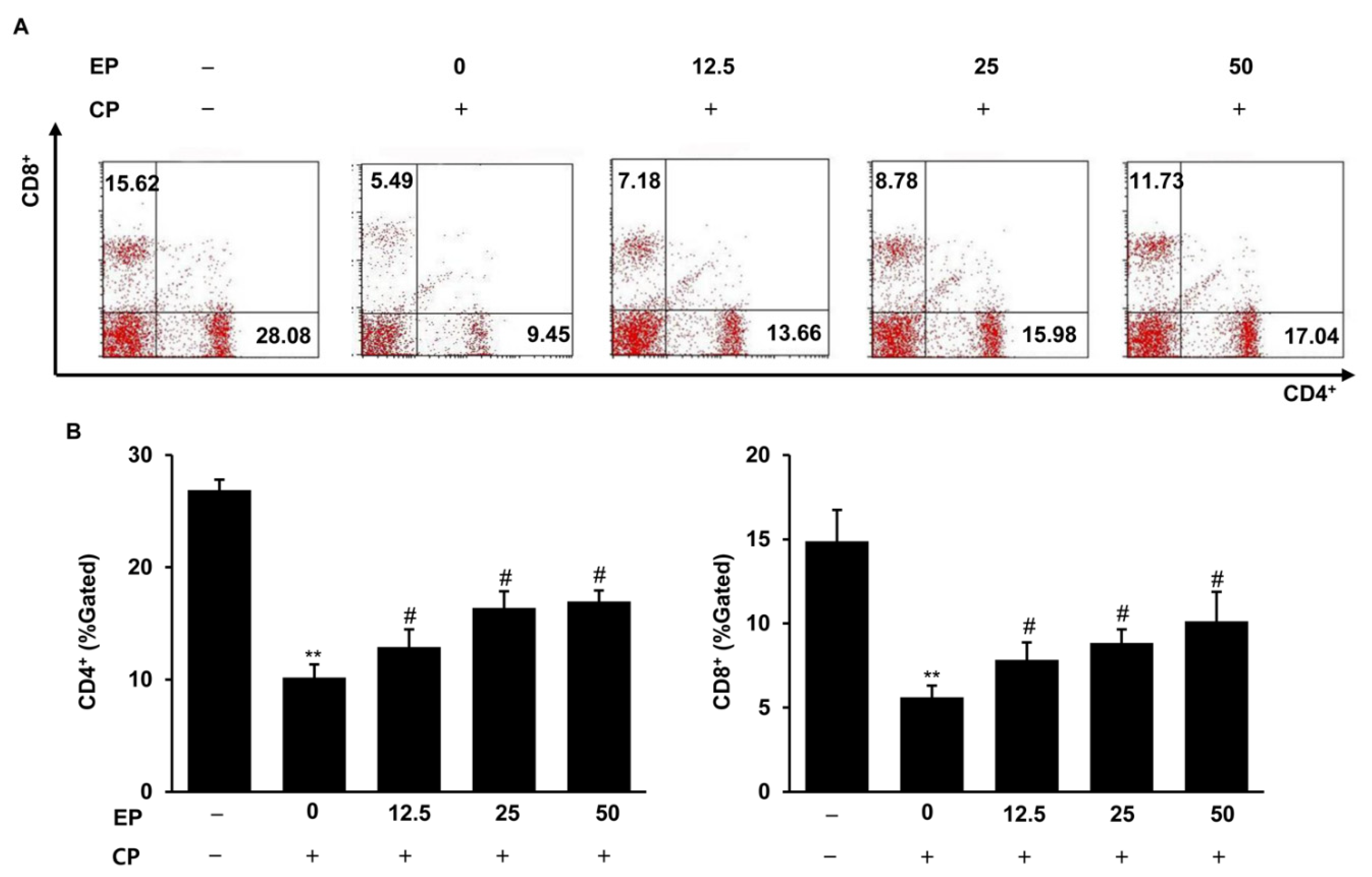
3.5. Effect of EP on Splenic T-Lymphocyte Subpopulations in the CP-Induced Immunosuppression Model
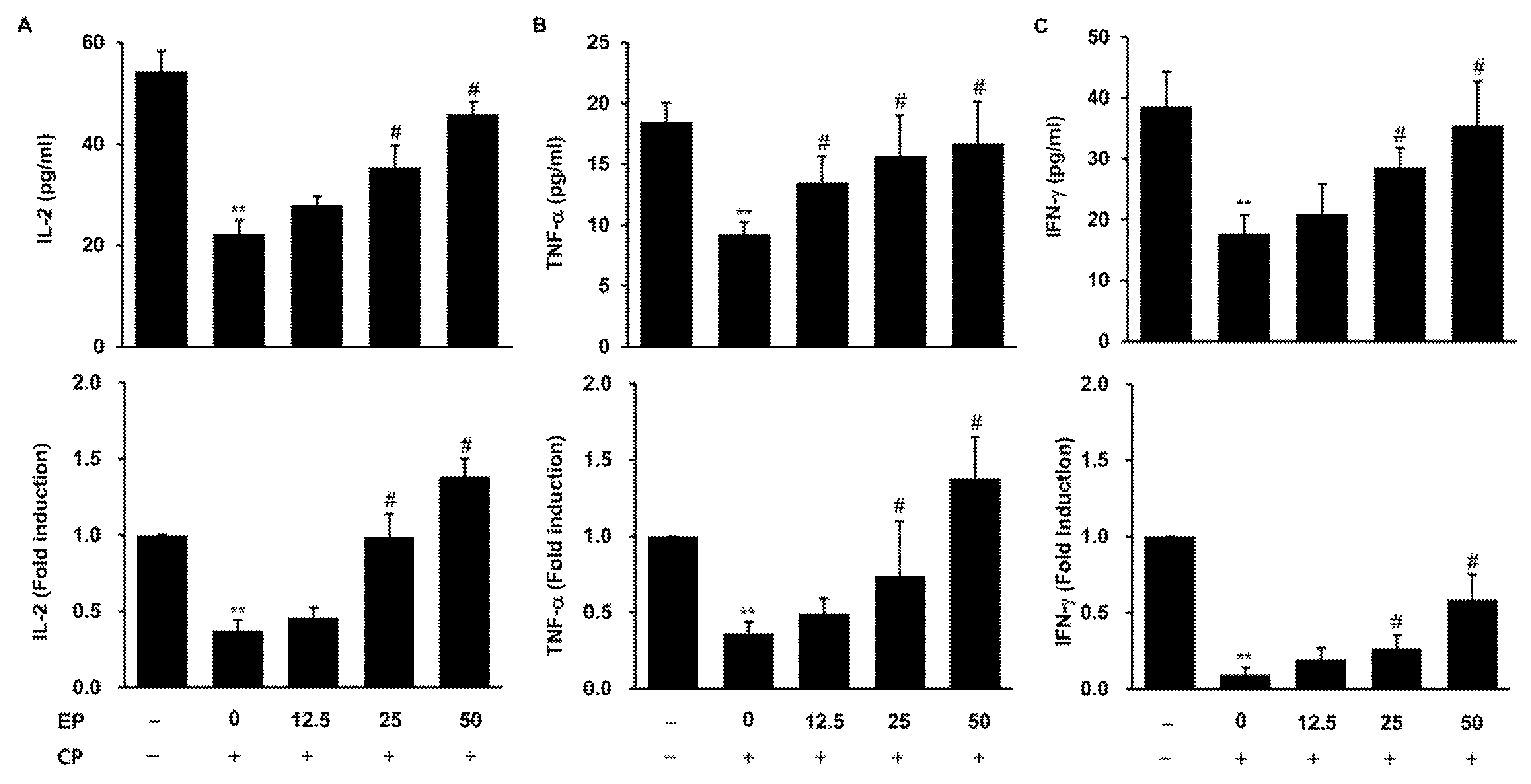
3.6. Effect of EP on Production of Immuno-Cytokines in the CP-Induced Immunosuppression Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Viladomiu, M.; Hontecillas, R.; Bassaganya-Riera, J. Modulation of inflammation and immunity by dietary conjugated linoleic acid. Eur. J. Pharmacol. 2016, 785, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Balogh, P.; Horvath, G.; Szakal, A.K. Immunoarchitecture of distinct reticular fibroblastic domains in the white pulp of mouse spleen. J. Histochem. Cytochem. 2004, 52, 1287–1298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolte, M.A.; Hamann, A.; Kraal, G.; Mebius, R.E. The strict regulation of lymphocyte migration to splenic white pulp does not involve common homing receptors. Immunology 2002, 106, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Lori, A.; Perrotta, M.; Lembo, G.; Carnevale, D. The spleen: A hub connecting nervous and immune systems in cardiovascular and metabolic diseases. Int. J. Mol. Sci. 2017, 18, 1216. [Google Scholar] [CrossRef] [PubMed]
- Mebius, R.E.; Kraal, G. Structure and function of the spleen. Nat. Rev. Immunol. 2005, 5, 606–616. [Google Scholar] [CrossRef] [PubMed]
- Im, S.A.; Kim, K.; Lee, C.K. Immunomodulatory activity of polysaccharides isolated from Salicornia herbacea. Int. Immunopharmacol. 2006, 6, 1451–1458. [Google Scholar] [CrossRef]
- Spiering, M.J. Primer on the immune system. Alcohol. Res. 2015, 37, 171–175. [Google Scholar]
- Pass, G.J.; Carrie, D.; Boylan, M.; Lorimore, S.; Wright, E.; Houston, B.; Henderson, C.J.; Wolf, C.R. Role of hepatic cytochrome p450s in the pharmacokinetics and toxicity of cyclophosphamide: Studies with the hepatic cytochrome p450 reductase null mouse. Cancer Res. 2005, 65, 4211–4217. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.N.; Jena, G.B. Intervention of astaxanthin against cyclophosphamide-induced oxidative stress and DNA damage: A study in mice. Chem. Biol. Interact. 2009, 80, 398–406. [Google Scholar] [CrossRef]
- Pratheeshkumar, P.; Kuttan, G. Ameliorative action of Vernonia cinerea L. on cyclophosphamide-induced immunosuppression and oxidative stress in mice. Inflammopharmacology 2010, 18, 197–207. [Google Scholar] [CrossRef]
- Rabinovitch, A.; Sorensen, O.; Suarez-Pinzon, W.L.; Power, R.F.; Rajotte, R.V.; Bleackley, R.C. Analysis of cytokine mRNA expression in syngeneic islet grafts of NOD mice: Interleukin 2 and interferon gamma mRNA expression correlate with graft rejection and interleukin 10 with graft survival. Diabetologia 1994, 37, 833–837. [Google Scholar] [CrossRef]
- Yu, Q.; Nie, S.P.; Wang, J.Q.; Huang, D.F.; Li, W.J.; Xie, M.Y. Molecular mechanism underlying chemoprotective effects of Ganoderma atrum polysaccharide in cyclophosphamide-induced immunosuppressed mice. J. Funct. Foods 2015, 15, 52–60. [Google Scholar] [CrossRef]
- Qi, Q.; Dong, Z.; Sun, Y.; Li, S.; Zhao, Z. Protective Effect of Bergenin against Cyclophosphamide-Induced Immunosuppression by Immunomodulatory Effect and Antioxidation in Balb/c Mice. Molecules 2018, 23, 2668. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.E.; Monmai, C.; Rod-In, W.; Jang, A.Y.; You, S.G.; Lee, S.M.; Park, W.J. Immune Enhancement Effects of Codium fragile Anionic Macromolecules Combined with Red Ginseng Extract in Immune-Suppressed Mice. J. Microbiol. Biotechnol. 2019, 17, 1213–1216. [Google Scholar] [CrossRef]
- Gao, H.Y.; Li, G.Y.; Huang, J.; Han, Y.; Sun, F.Z.; Du, X.W.; An, L.J.; Wang, H.Y.; Wang, J.H. Protective effects of Zhuyeqing liquor on the immune function of normal and immunosuppressed mice in vivo. BMC Complement. Altern. Med. 2013, 13, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markman, M. Chemotherapy-associated neurotoxicity: An important side effect-impacting on quality, rather than quantity, of life. J. Cancer Res. Clin. Oncol. 1996, 122, 511–512. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Silván, B.; Entrialgo-Cadierno, R.; Villar, C.J.; Capasso, R.; Uranga, J.A.; Lombó, F.; Abalo, R. Antiproliferative and palliative activity of flavonoids in colorectal cancer. Biomed. Pharmacother. 2021, 143, 112241. [Google Scholar] [CrossRef]
- Küpeli Akkol, E.; Genç, Y.; Karpuz, B.; Sobarzo-Sánchez, E.; Capasso, R. Coumarins and Coumarin-Related Compounds in Pharmacotherapy of Cancer. Cancers 2020, 12, 1959. [Google Scholar] [CrossRef] [PubMed]
- Barnes, J.; Anderson, L.A.; Gibbons, S.; Phillipson, J.D. Echinacea species (Echinacea angustifolia (DC.) Hell., Echinacea pallida (Nutt.) Nutt., Echinacea purpurea (L.) Moench): A review of their chemistry, pharmacology and clinical properties. J. Pharm. Pharmacol. 2005, 57, 929–954. [Google Scholar] [CrossRef] [Green Version]
- Hueza, I.M.; Gotardo, A.T.; da Silva Mattos, M.I.; Górniak, S.L. Immunomodulatory effect of Cynara scolymus (artichoke) in rats. Phytother. Res. 2019, 33, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Karimi, M.H.; Ebrahimnezhad, S.; Namayandeh, M.; Amirghofran, Z. The effects of cichorium intybus extract on the maturation and activity of dendritic cells. DARU J. Pharm. Sci. 2014, 22, 28. [Google Scholar] [CrossRef] [Green Version]
- Hohmann, J.; R’edei, D.; Forgo, P.; Szab´o, P.; Freund, T.F.; Haller, J.; Bojnik, E.; Benyhe, S. Alkamides and a neolignan from Echinacea purpurea roots and the interaction of alkamides with G-protein-coupled cannabinoid receptors. Phytochemistry 2011, 72, 1848–1853. [Google Scholar] [CrossRef]
- Woelkart, K.; Bauer, R. The role of alkamides as an active principle of echinacea. Planta Med. 2007, 73, 615–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, M.; Anderson, S.A.; Schoop, R.; Hudson, J.B. Induction of multiple pro-inflammatory cytokines by respiratory viruses and reversal by standardized Echinacea, a potent antiviral herbal extract. Antivir. Res. 2009, 83, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Todd, D.A.; Gulledge, T.V.; Britton, E.R.; Oberhofer, M.; Leyte-Lugo, M.; Moody, A.N.; Shymanovich, T.; Grubbs, L.F.; Juzumaite, M.; Tyler, N.G.; et al. Ethanolic Echinacea purpurea extracts contain a mixture of cytokine-suppressive and cytokine-inducing compounds, including some that originate from endophytic bacteria. PLoS ONE 2015, 10, e0124276. [Google Scholar]
- Lee, H.H.; Cho, Y.; Kim, G.H.; Cho, H. Undaria pinnatifida Fucoidan-Rich Extract Recovers Immunity of Immunosuppressed Mice. J. Microbiol. Biotechnol. 2020, 30, 439–447. [Google Scholar] [CrossRef] [PubMed]
- Abouelella, A.M.; Shahein, Y.E.; Tawfik, S.S.; Zahran, A.M. Phytotherapeutic effects of Echinacea purpurea in gamma-irradiated mice. J. Vet. Sci. 2007, 8, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Raso, G.M.; Pacilio, M.; Di Carlo, G.; Esposito, E.; Pinto, L.; Meli, R. In-vivo and in-vitro anti-inflammatory effect of Echinacea purpurea and Hypericum perforatum. J. Pharm. Pharmacol. 2002, 54, 1379–1383. [Google Scholar] [CrossRef]
- Gao, T.; Bi, H.; Ma, S.; Lu, J. The antitumor and immunostimulating activities of water soluble polysaccharides from Radix Aconiti, Radix Aconiti Lateralis and Radix Aconiti Kusnezoffii. Nat. Prod. Commun. 2010, 5, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.R.; Lee, H.S.; Cho, S.Y.; Kim, Y.S.; Shin, K.S. Antimetastatic effect of polysaccharide isolated from Colocasia esculenta is exerted through immunostimulation. Int. J. Mol. Med. 2013, 31, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Qi, Z.; Chen, L.; Li, Z.; Shao, Z.; Qi, Y.; Gao, K.; Liu, S.; Sun, Y.; Li, P.; Liu, J. Immunomodulatory effects of (24R)-Pseudo-Ginsenoside HQ and (24S)-Pseudo- Ginsenoside HQ on cyclophosphamide-induced immunosuppression and their anti-tumor effects study. Int. J. Mol. Sci. 2019, 20, 836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Conlon, M.; Ren, W.; Chen, B.B.; Baczek, T. Natural products as targeted modulators of the immune system. J. Immunol. Res. 2018, 2018, 7862782. [Google Scholar] [CrossRef] [PubMed]
- Asherson, R.A.; Gunter, K.; Daya, D.; Shoenfeld, Y. Multiple Autoimmune Diseases in a Young Woman: Tuberculosis and Splenectomy as Possible Triggering Factors? Another Example of the “Mosaic” of Autoimmunity. J. Rheumatol. 2008, 35, 1224–1227. [Google Scholar] [PubMed]
- Cheng, Z.Y.; Sun, X.; Liu, P.; Lin, B.; Li, L.Z.; Yao, G.D.; Huang, X.X.; Song, S.J. Sesquiterpenes from Echinacea purpurea and their anti-inflammatory activities. Phytochemistry 2020, 179, 112503. [Google Scholar] [CrossRef]
- Zorig, A.; Toko, R.; Sukhbold, E.; Takasugi, M.; Arai, H. Echinacea purpurea water extracts suppress the release of chemical mediators from mast cells. Biosci. Biotechnol. Biochem. 2021, 85, 931–940. [Google Scholar] [CrossRef]
- Wang, H.; Wang, M.; Chen, J.; Tang, Y.; Dou, J.; Yu, J.; Xi, T.; Zhou, C. A polysaccharide from Strongylocentrotus nudus eggs protects against myelosuppression and immunosuppression in cyclophosphamide-treated mice. Int. Immunopharmacol. 2011, 11, 1946–1953. [Google Scholar] [CrossRef] [PubMed]
- Tanahashi, T.; Sekiguchi, N.; Matsuda, K.; Matsumoto, A.; Ito, T.; Nakazawa, H.; Ishida, F. A screening method with lymphocyte percentage and proportion of granular lymphocytes in the peripheral blood for large granular lymphocyte (LGL) leukemia. Int. J. Hematol. 2017, 105, 87–91. [Google Scholar] [CrossRef]
- Cui, H.L.; Chen, Y.; Wang, S.S.; Kai, G.Q.; Fang, Y.M. Isolation, partial characterisation and immunomodulatory activities of polysaccharide from Morchella esculenta. J. Sci. Food Agric. 2011, 91, 2180–2185. [Google Scholar] [CrossRef]
- Meng, Y.; Li, B.; Jin, D.; Zhan, M.; Lu, J.; Huo, G. Immunomodulatory activity of Lactobacillus plantarum KLDS1.0318 in cyclophosphamide-treated mice. Food Nutr. Res. 2018, 62, 1269. [Google Scholar] [CrossRef]
- Kawabata, T.T.; White, K.L. Suppression of the vitro humoral immune response of mouse splenocytes by benzo(a)pyrene metabolites and inhibition of benzo(a)pyrene-induced immunosuppression by alpha-naphthoflavone. Cancer Res. 1987, 47, 2317–2322. [Google Scholar]
- Wang, J.; Tong, X.; Li, P.; Cao, H.; Su, W. Immuno-enhancement effects of shenqi fuzheng injection on cyclophosphamide-induced immunosuppression in BALB/c mice. J. Ethnopharmacol. 2014, 139, 788–795. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Lu, Y.; Wang, D.; Liu, J.; Song, X.; Zhang, W.; Zhao, X.; Nguyen, T.L.; Hu, Y. Effect of epimedium polysaccharide-propolis flavone immunopotentiator on immunosuppression induced by cyclophosphamide in chickens. Cell. Immunol. 2013, 281, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Duggina, P.; Kalla, C.M.; Varikasuvu, S.R.; Bukke, S.; Tartte, V. Protective effect of centella triterpene saponins against cyclophosphamide-induced immune and hepatic system dysfunction in rats: Its possible mechanisms of action. J. Physiol. Biochem. 2015, 71, 435–454. [Google Scholar] [CrossRef]
- Kos, F.J.; Engleman, E.G. Immune regulation: A critical link between NK cells and CTLs. Immunol. Today 1996, 17, 174–176. [Google Scholar] [CrossRef]
- Constant, S.L.; Bottomly, K. Induction of Th1 and Th2 CD4+ T cell responses: The alternative approaches. Annu. Rev. Immunol. 1997, 15, 297–322. [Google Scholar] [CrossRef]
- Zhu, X.L.; Chen, A.F.; Lin, Z.B. Ganoderma lucidum polysaccharides enhance the function of immunological effector cells in immunosuppressed mice. J. Ethnopharmacol. 2007, 111, 219–226. [Google Scholar] [CrossRef]
- Mosmann, T.R.; Coffman, R.L. TH1 and TH2 cells: Different patterns of lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 1989, 7, 145–173. [Google Scholar] [CrossRef] [PubMed]
- Erenler, R.; Telci, I.; Ulutas, M.; Demirtas, I.; Gul, F.; Elmastas, M.; Kayir, O. Chemical constituents, quantitative analysis and antioxidant activities of Echinacea purpurea (L.) Moench and Echinacea pallida (Nutt.) Nutt. J. Food Biochem. 2015, 39, 622–630. [Google Scholar] [CrossRef]
- Chiellini, C.; Maida, I.; Maggini, V.; Bosi, E.; Mocali, S.; Emiliani, G.; Perrin, E.; Firenzuoli, F.; Mengoni, A.; Fani, R. Preliminary data on antibacterial activity of Echinacea purpurea-associated bacterial communities against Burkholderia cepacia complex strains, opportunistic pathogens of Cystic Fibrosis patients. Microbiol. Res. 2017, 196, 34–43. [Google Scholar] [CrossRef]
- Park, S.; Lee, M.S.; Jung, S.; Lee, S.; Kwon, O.; Kreuter, M.H.; Perrinjaquet-Moccetti, T.; Min, B.; Yun, S.H.; Kim, Y. Echinacea purpurea Protects Against Restraint Stress-Induced Immunosuppression in BALB/c Mice. J. Med. Food 2018, 21, 261–268. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.-R.; Kim, Y.-S.; Lee, D.-R.; Choi, B.-K.; Kwon, K.-B.; Bae, G.-S. Echinacea purpurea Alleviates Cyclophosphamide-Induced Immunosuppression in Mice. Appl. Sci. 2022, 12, 105. https://doi.org/10.3390/app12010105
Kim H-R, Kim Y-S, Lee D-R, Choi B-K, Kwon K-B, Bae G-S. Echinacea purpurea Alleviates Cyclophosphamide-Induced Immunosuppression in Mice. Applied Sciences. 2022; 12(1):105. https://doi.org/10.3390/app12010105
Chicago/Turabian StyleKim, Ha-Rim, Ye-Seul Kim, Dong-Ryung Lee, Bong-Keun Choi, Kang-Beom Kwon, and Gi-Sang Bae. 2022. "Echinacea purpurea Alleviates Cyclophosphamide-Induced Immunosuppression in Mice" Applied Sciences 12, no. 1: 105. https://doi.org/10.3390/app12010105
APA StyleKim, H.-R., Kim, Y.-S., Lee, D.-R., Choi, B.-K., Kwon, K.-B., & Bae, G.-S. (2022). Echinacea purpurea Alleviates Cyclophosphamide-Induced Immunosuppression in Mice. Applied Sciences, 12(1), 105. https://doi.org/10.3390/app12010105