Gallic Acid Induces S and G2 Phase Arrest and Apoptosis in Human Ovarian Cancer Cells In Vitro

 , ,
, ,  ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Reagents
2.2. Assessment of Cell Viability
2.3. Hoechst 33342 Staining
2.4. Apoptosis Analysis by Flow Cytometry
2.5. Detection of Caspase-3/7 Enzyme Activities
2.6. Western Blot
2.7. Cell Cycle Evaluation Using Flow Cytometry
2.8. Statistical Analysis
3. Results
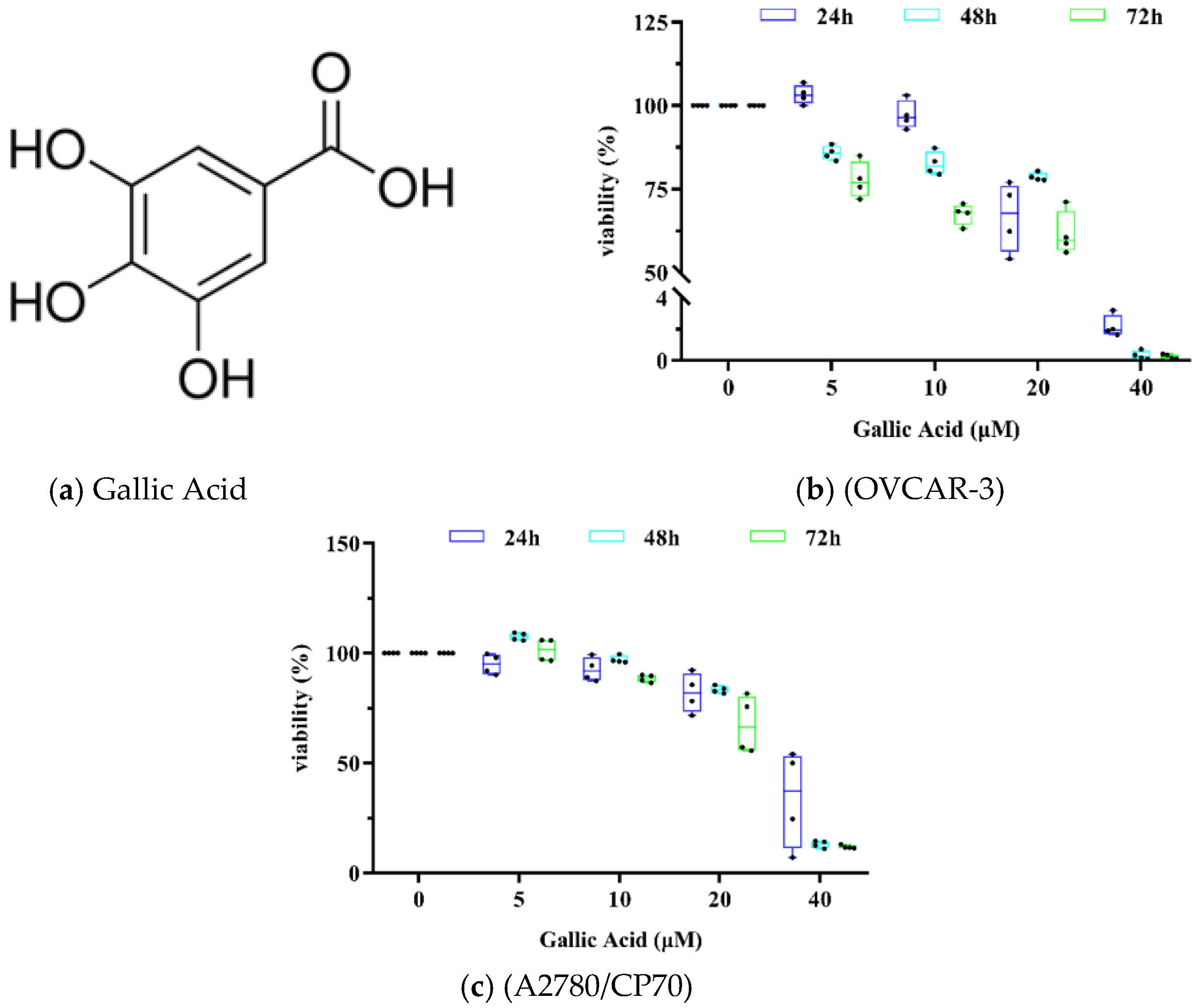
3.1. GA Inhibition of OC Cell Viability Is Dose Dependent
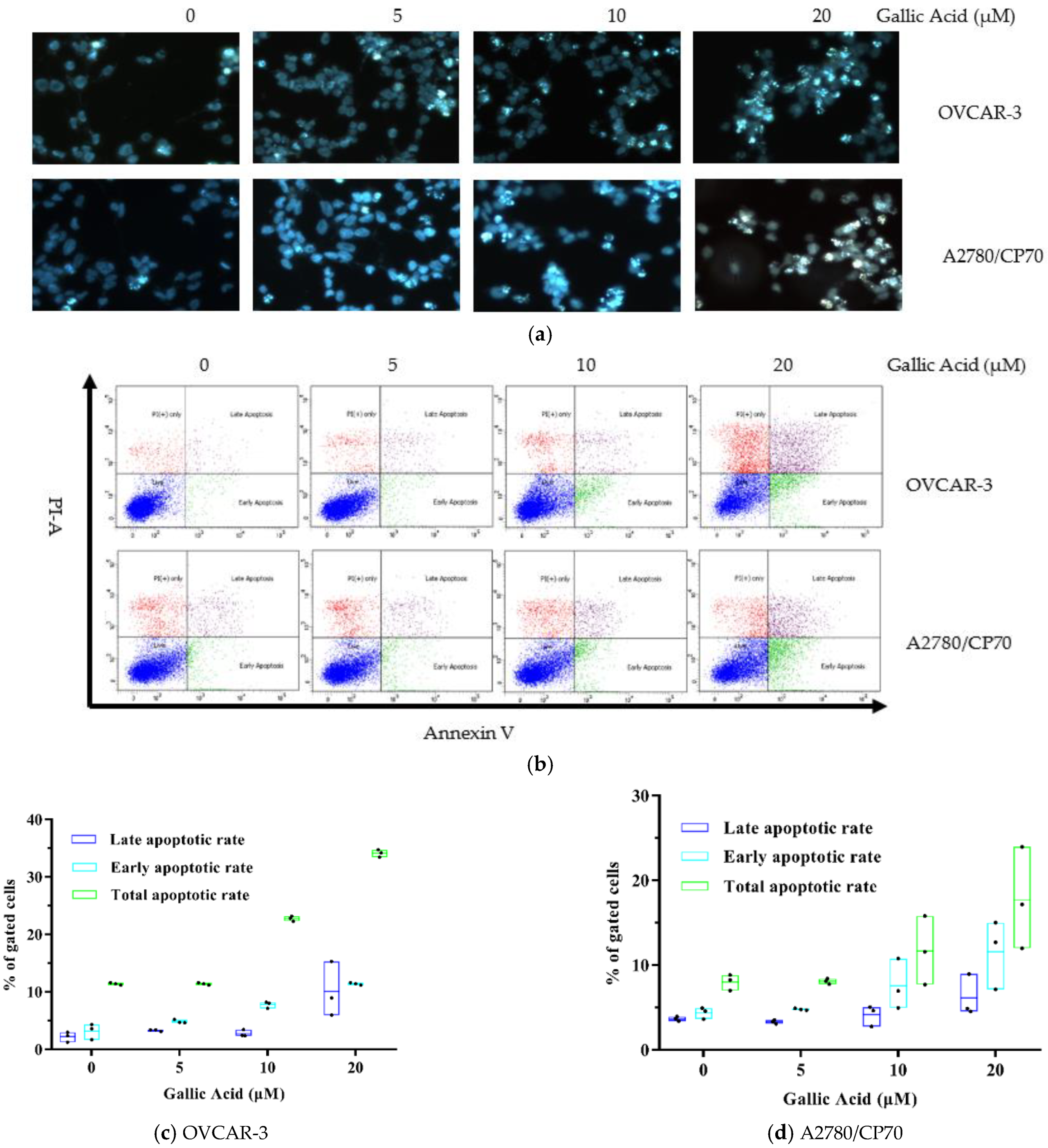
3.2. GA Promotes Cell Apoptosis in Both OC Cell Lines
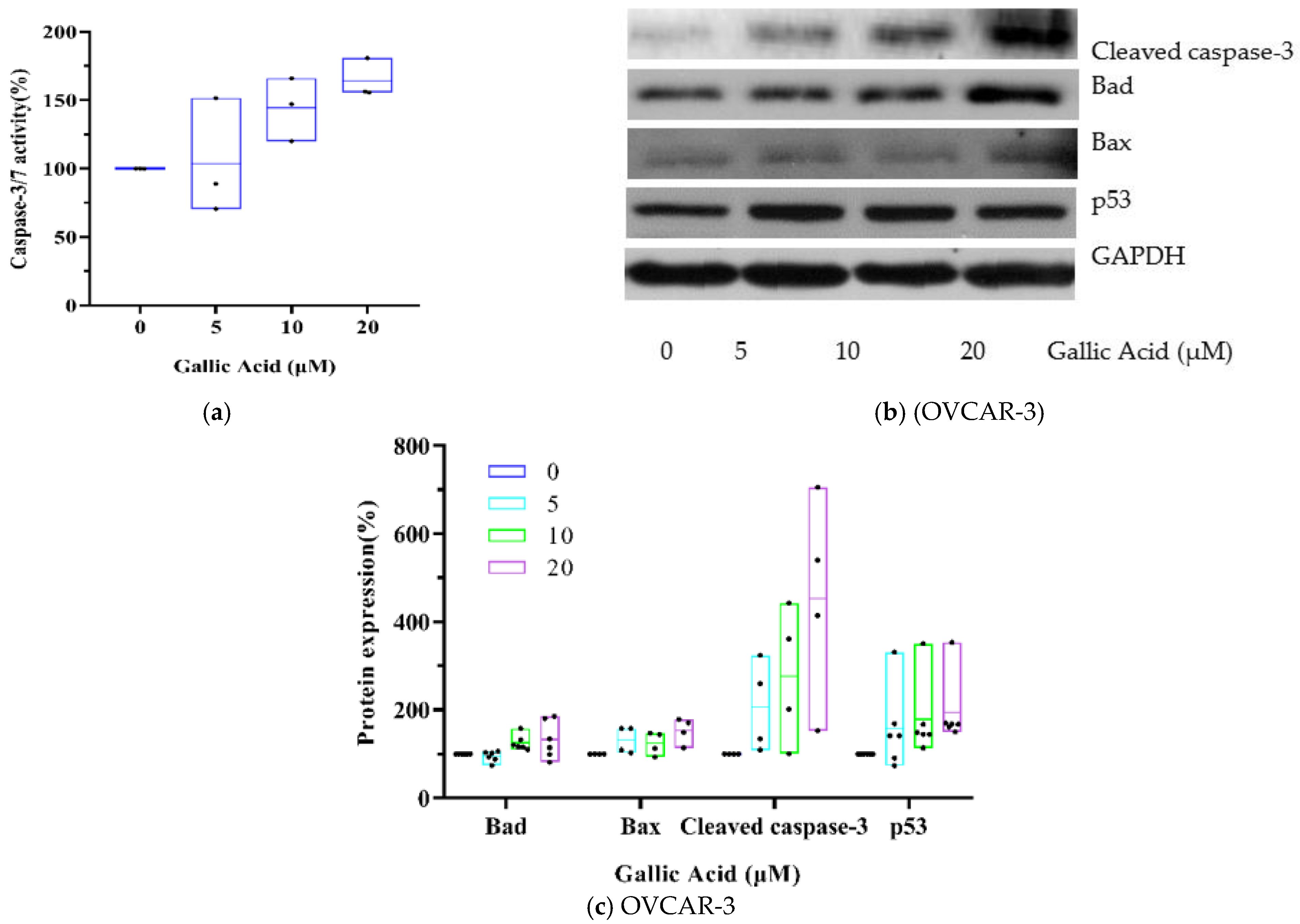
3.3. GA Activates Caspase-3 in OVCAR-3s via the p53-Dependent Intrinsic Pathway
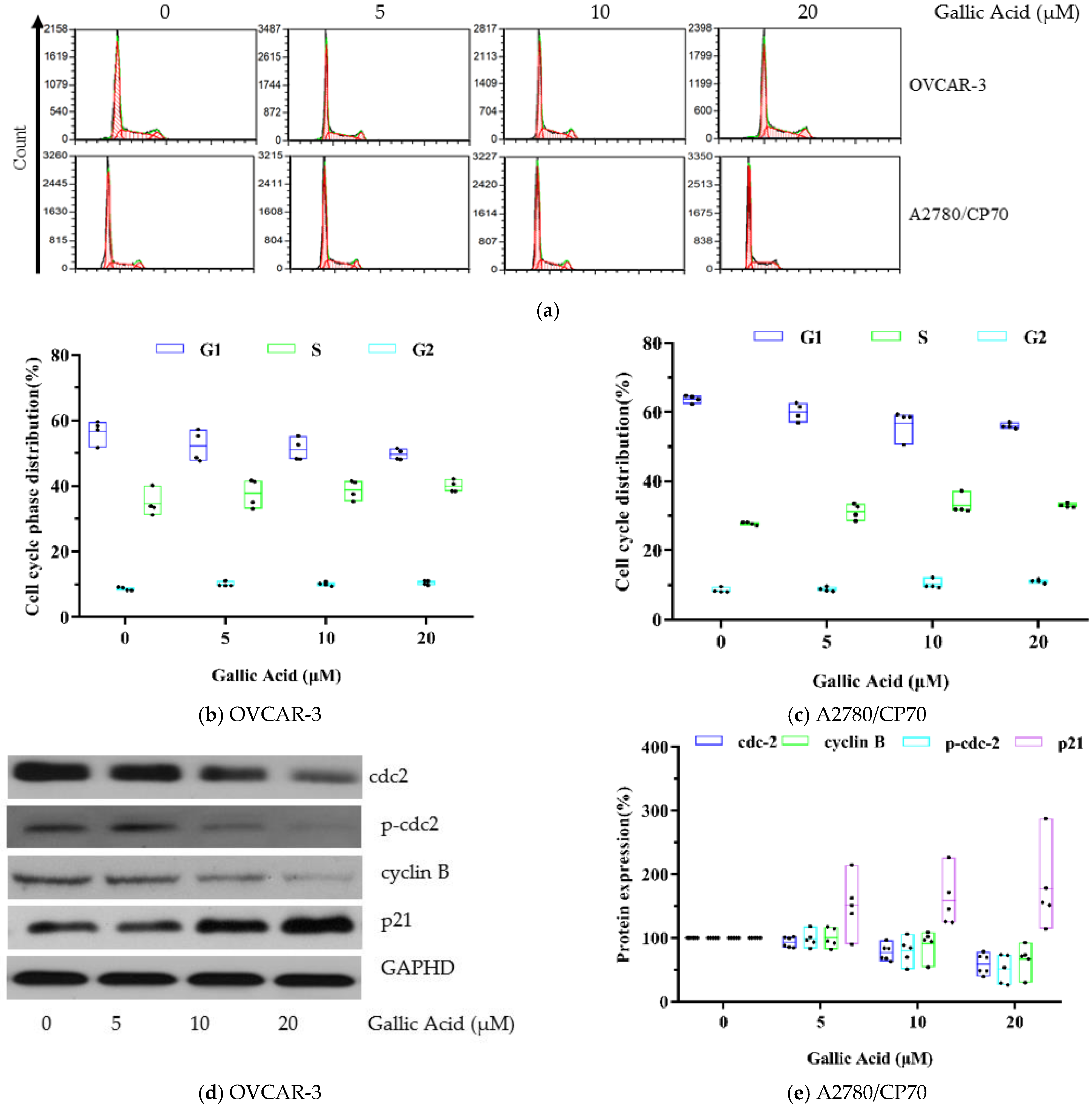
3.4. GA Causes S and G2 Phase Arrest in Both OC Cell Lines
3.5. GA Modulates Cell Cycle-Related Proteins Levels in OVCAR-3s
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef]
- Lheureux, S.; Msc, M.B.; Oza, A.M. Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA Cancer J. Clin. 2019, 69, 280–304. [Google Scholar] [CrossRef]
- Sznurkowski, J.J.; Bodnar, L.; Bidziński, M.; Bieńkiewicz, A.; Blecharz, P.; Jach, R.; Knapp, P.; Kojs, Z.; Kotarski, J.; Markowska, J.; et al. Recommendations of the Polish Gynecological Oncology Society for the diagnosis andtreatment of vulvar cancer. Curr. Gynecol. Oncol. 2017, 15, 45–53. [Google Scholar] [CrossRef]
- Kim, J.; Park, E.Y.; Kim, O.; Schilder, J.M.; Coffey, D.M.; Cho, C.-H.; Bast, J.R.C. Cell Origins of High-Grade Serous Ovarian Cancer. Cancers 2018, 10, 433. [Google Scholar] [CrossRef]
- Lavoué, V.; Thédrez, A.; Levêque, J.; Foucher, F.; Henno, S.; Jauffret, V.; Belaud-Rotureau, M.-A.; Catros, V.; Cabillic, F. Immunity of human epithelial ovarian carcinoma: The paradigm of immune suppression in cancer. J. Transl. Med. 2013, 11, 147. [Google Scholar] [CrossRef]
- Coleman, R.L.; Brady, M.F.; Herzog, T.J.; Sabbatini, P.; Armstrong, D.K.; Walker, J.L.; Kim, B.-G.; Fujiwara, K.; Tewari, K.S.; O’Malley, D.M.; et al. Bevacizumab and paclitaxel–carboplatin chemotherapy and secondary cytoreduction in recurrent, platinum-sensitive ovarian cancer (NRG Oncology/Gynecologic Oncology Group study GOG-0213): A multicentre, open-label, randomised, phase 3 trial. Lancet Oncol. 2017, 18, 779–791. [Google Scholar] [CrossRef]
- Li, S.-S.; Ma, J.; Wong, A.S.T. Chemoresistance in ovarian cancer: Exploiting cancer stem cell metabolism. J. Gynecol. Oncol. 2018, 29, e32. [Google Scholar] [CrossRef]
- Lee, H.-L.; Lin, C.-S.; Kao, S.-H.; Chou, M.-C. Gallic acid induces G1 phase arrest and apoptosis of triple-negative breast cancer cell MDA-MB-231 via p38 mitogen-activated protein kinase/p21/p27 axis. Anti Cancer Drugs 2017, 28, 1150–1156. [Google Scholar] [CrossRef]
- Kang, D.Y.; Sp, N.; Jo, E.S.; Rugamba, A.; Hong, D.Y.; Lee, H.G.; Yoo, J.-S.; Liu, Q.; Jang, K.-J.; Yang, Y.M. The Inhibitory Mechanisms of Tumor PD-L1 Expression by Natural Bioactive Gallic Acid in Non-Small-Cell Lung Cancer (NSCLC) Cells. Cancers 2020, 12, 727. [Google Scholar] [CrossRef]
- Lin, M.-L.; Chen, S.-S. Activation of Casein Kinase II by Gallic Acid Induces BIK–BAX/BAK-Mediated ER Ca++-ROS-Dependent Apoptosis of Human Oral Cancer Cells. Front. Physiol. 2017, 8, 8. [Google Scholar] [CrossRef]
- Jang, Y.-G.; Ko, E.-B.; Choi, K.-C. Gallic acid, a phenolic acid, hinders the progression of prostate cancer by inhibition of histone deacetylase 1 and 2 expression. J. Nutr. Biochem. 2020, 84, 108444. [Google Scholar] [CrossRef]
- Aborehab, N.M.; Osama, N. Effect of Gallic acid in potentiating chemotherapeutic effect of Paclitaxel in HeLa cervical cancer cells. Cancer Cell Int. 2019, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Li, B.; Rankin, G.O.; Rojanasakul, Y.; Chen, Y.C. Selecting bioactive phenolic compounds as potential agents to inhibit proliferation and VEGF expression in human ovarian cancer cells. Oncol. Lett. 2015, 9, 1444–1450. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Chen, A.Y.; Rojanasakul, Y.; Rankin, G.O.; Chen, Y.C. Gallic acid, a phenolic compound, exerts anti-angiogenic effects via the PTEN/AKT/HIF-1α/VEGF signaling pathway in ovarian cancer cells. Oncol. Rep. 2015, 35, 291–297. [Google Scholar] [CrossRef] [PubMed]
- Varela-Rodríguez, L.; Sánchez-Ramírez, B.; Hernández-Ramírez, V.I.; Varela-Rodríguez, H.; Castellanos-Mijangos, R.D.; González-Horta, C.; Chávez-Munguía, B.; Talamás-Rohana, P. Effect of Gallic acid and Myricetin on ovarian cancer models: A possible alternative antitumoral treatment. BMC Complement. Med. Ther. 2020, 20, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gao, Y.; Rankin, G.O.; Rojanasakul, Y.; Cutler, S.J.; Tu, Y.; Chen, Y.C. Chaetoglobosin K induces apoptosis and G2 cell cycle arrest through p53-dependent pathway in cisplatin-resistant ovarian cancer cells. Cancer Lett. 2015, 356, 418–433. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, N.; Rankin, G.O.; Li, B.; Rojanasakul, Y.; Tu, Y.; Chen, Y.C. Anti-proliferative effect and cell cycle arrest induced by saponins extracted from tea (Camellia sinensis) flower in human ovarian cancer cells. J. Funct. Foods 2017, 37, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.-F.; Li, X.-X.; Zhao, Y.-H.; Kang, J.-K.; Li, J.-Y.; Tian, W.; Wang, C.-M.; Hou, H.-X.; Li, D.-R. Confirming whether novel rhein derivative 4a induces paraptosis-like cell death by endoplasmic reticulum stress in ovarian cancer cells. Eur. J. Pharmacol. 2020, 886, 173526. [Google Scholar] [CrossRef]
- Garcia-Arguello, S.F.; Lopez-Lorenzo, B.; Cornelissen, B.; Smith, G. Development of F-18 ICMT-11 for Imaging Caspase-3/7 Activity during Therapy-Induced Apoptosis. Cancers 2020, 12, 2191. [Google Scholar] [CrossRef]
- Aborehab, N.M.; Elnagar, M.R.; Waly, N.E. Gallic acid potentiates the apoptotic effect of paclitaxel and carboplatin via overexpression of Bax and P53 on the MCF-7 human breast cancer cell line. J. Biochem. Mol. Toxicol. 2021, 35, e22638. [Google Scholar] [CrossRef]
- Zeng, M.; Su, Y.; Li, K.; Jin, D.; Li, Q.; Li, Y.; Zhou, B. Gallic Acid Inhibits Bladder Cancer T24 Cell Progression Through Mitochondrial Dysfunction and PI3K/Akt/NF-κB Signaling Suppression. Front. Pharmacol. 2020, 11, 1222. [Google Scholar] [CrossRef]
- Gao, L.; Wang, Z.; Lu, D.; Huang, J.; Liu, J.; Hong, L. Paeonol induces cytoprotective autophagy via blocking the Akt/mTOR pathway in ovarian cancer cells. Cell Death Dis. 2019, 10, 1–13. [Google Scholar] [CrossRef]
- Anaya-Eugenio, G.D.; Tan, C.Y.; Rakotondraibe, L.H.; De Blanco, E.C.C. Tumor suppressor p53 independent apoptosis in HT-29 cells by auransterol from Penicillium aurantiacobrunneum. Biomed. Pharmacother. 2020, 127, 110124. [Google Scholar] [CrossRef]
- Sharma, H.; Stephen, N.M.; Gopal, S.S.; Rudresh, D.U.; Kavalappa, Y.P.; Shivarudrappa, A.H.; Gavirangappa, H.; Ponesakki, G. Phenolic Extract of Seagrass, Halophila ovalis Activates Intrinsic Pathway of Apoptosis in Human Breast Cancer (MCF-7) Cells. Nutr. Cancer 2021, 73, 307–317. [Google Scholar] [CrossRef]
- Naoi, M.; Wu, Y.; Shamoto-Nagai, M.; Maruyama, W. Mitochondria in Neuroprotection by Phytochemicals: Bioactive Polyphenols Modulate Mitochondrial Apoptosis System, Function and Structure. Int. J. Mol. Sci. 2019, 20, 2451. [Google Scholar] [CrossRef]
- Sourani, Z.; Pourgheysari, B.; Beshkar, P.; Shirzad, H.; Shirzad, M. Gallic Acid Inhibits Proliferation and Induces Apoptosis in Lymphoblastic Leukemia Cell Line (C121). Iran. J. Med. Sci. 2016, 41, 525–530. [Google Scholar]
- Moghtaderi, H.; Sepehri, H.; Delphi, L.; Attari, F. Gallic acid and curcumin induce cytotoxicity and apoptosis in human breast cancer cell MDA-MB-231. BioImpacts 2018, 8, 185–194. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef]
- Greenway, F.L.; Roberts, A.T.; Martin, C.K.; Liu, Z.; Amen, R.J.; Woltering, E.A.; Rood, J.C.; Caruso, M.K.; Yu, Y.; Xie, H. The Safety and Efficacy of a Dietary Herbal Supplement and Gallic Acid for Weight Loss. J. Med. Food 2007, 10, 184–188. [Google Scholar]
- Pitica, I.M.; Matei, L.; Dragu, L.D.; Alexandru, G.; Suciu, A.; Alexiu, I.D.; Neagu, A.I.; Chivu-Economescu, M.; Crisan, I.; Cristina, M.; et al. Tamarix gallica extract induces apoptosis via an intrinsic mechanism. Rom. Biotechnol. Lett. 2020, 25, 1701–1708. [Google Scholar]
- He, Z.; Wu, S.; Lin, J.; Booth, A.; Rankin, G.O.; Martinez, I.; Chen, Y.C. Polyphenols Extracted from Chinese Hickory (Carya Cathayensis) Promote Apoptosis and Inhibit Proliferation through the p53-Dependent Intrinsic and HIF-1α-VEGF Pathways in Ovarian Cancer Cells. Appl. Sci. 2020, 10, 8615. [Google Scholar] [CrossRef]
- Sankhe, N.M.; Durgashivaprasad, E.; Kutty, N.G.; Rao, J.V.; Narayanan, K.; Kumar, N.; Jain, P.; Udupa, N.; Raj, P.V. Novel 2,5-disubstituted-1,3,4-oxadiazole derivatives induce apoptosis in HepG2 cells through p53 mediated intrinsic pathway. Arab. J. Chem. 2019, 12, 2548–2555. [Google Scholar] [CrossRef]
- Luo, H.; Rankin, G.O.; Li, Z.; DePriest, L.; Chen, Y.C. Kaempferol induces apoptosis in ovarian cancer cells through activating p53 in the intrinsic pathway. Food Chem. 2011, 128, 513–519. [Google Scholar] [CrossRef]
- Lomax, M.E.; Barnes, D.M.; Hupp, T.R.; Picksley, S.M.; Camplejohn, R.S. Characterization of p53 Oligomerization Domain Mutations Isolated from Li–Fraumeni and Li–Fraumeni like Family Members. Oncogene 1998, 17, 643–649. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, S.; Wei, C.; Rankin, G.O.; Rojanasakul, Y.; Ren, N.; Ye, X.; Chen, Y.C. Dietary compound proanthocyanidins from Chinese bayberry (Myrica rubra Sieb. et Zucc.) leaves inhibit angiogenesis and regulate cell cycle of cisplatin-resistant ovarian cancer cells via targeting Akt pathway. J. Funct. Foods 2018, 40, 573–581. [Google Scholar] [CrossRef]
- Gao, Y.; Yin, J.; Tu, Y.; Chen, Y.C. Theaflavin-3,3′-Digallate Suppresses Human Ovarian Carcinoma OVCAR-3 Cells by Regulating the Checkpoint Kinase 2 and p27 kip1 Pathways. Molecules 2019, 24, 673. [Google Scholar] [CrossRef]
- Liao, W.-L.; Lin, J.-Y.; Shieh, J.-C.; Yeh, H.-F.; Hsieh, Y.-H.; Cheng, Y.-C.; Lee, H.-J.; Shen, C.-Y.; Cheng, C.-W. Induction of G2/M Phase Arrest by Diosgenin via Activation of Chk1 Kinase and Cdc25C Regulatory Pathways to Promote Apoptosis in Human Breast Cancer Cells. Int. J. Mol. Sci. 2019, 21, 172. [Google Scholar] [CrossRef]
- Hung, K.-C.; Wang, S.-G.; Lin, M.-L.; Chen, S.-S. Citrate-Induced p85α–PTEN Complex Formation Causes G2/M Phase Arrest in Human Pharyngeal Squamous Carcinoma Cell Lines. Int. J. Mol. Sci. 2019, 20, 2105. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Liu, X.; Wu, F.; Wu, S.; Rankin, G.O.; Martinez, I.; Rojanasakul, Y.; Chen, Y.C. Gallic Acid Induces S and G2 Phase Arrest and Apoptosis in Human Ovarian Cancer Cells In Vitro. Appl. Sci. 2021, 11, 3807. https://doi.org/10.3390/app11093807
He Z, Liu X, Wu F, Wu S, Rankin GO, Martinez I, Rojanasakul Y, Chen YC. Gallic Acid Induces S and G2 Phase Arrest and Apoptosis in Human Ovarian Cancer Cells In Vitro. Applied Sciences. 2021; 11(9):3807. https://doi.org/10.3390/app11093807
Chicago/Turabian StyleHe, Zhiping, Xingquan Liu, Fenghua Wu, Shaozhen Wu, Gary O’Neal Rankin, Ivan Martinez, Yon Rojanasakul, and Yi Charlie Chen. 2021. "Gallic Acid Induces S and G2 Phase Arrest and Apoptosis in Human Ovarian Cancer Cells In Vitro" Applied Sciences 11, no. 9: 3807. https://doi.org/10.3390/app11093807
APA StyleHe, Z., Liu, X., Wu, F., Wu, S., Rankin, G. O., Martinez, I., Rojanasakul, Y., & Chen, Y. C. (2021). Gallic Acid Induces S and G2 Phase Arrest and Apoptosis in Human Ovarian Cancer Cells In Vitro. Applied Sciences, 11(9), 3807. https://doi.org/10.3390/app11093807