
Isoprene Production from Municipal Wastewater Biosolids by Engineered Archaeon Methanosarcina acetivorans


Abstract
Featured Application
Abstract
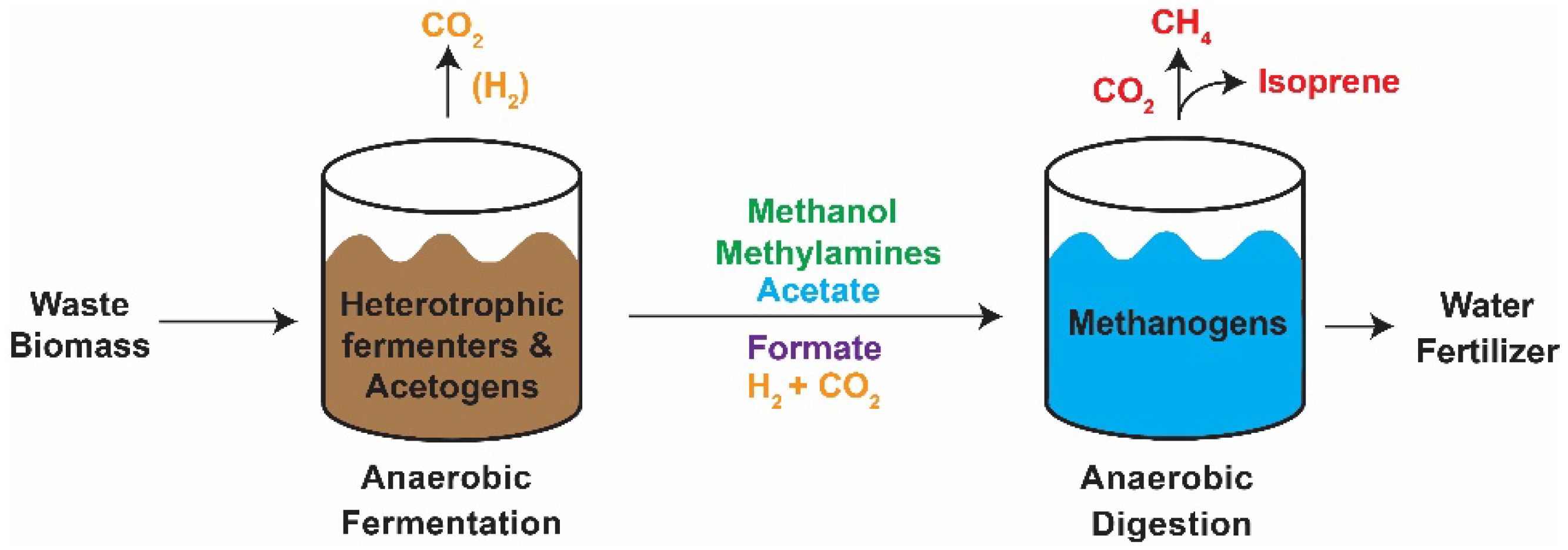
1. Introduction
2. Materials and Methods
2.1. Anaerobic Techniques
2.2. Synthetic Wastewater
2.3. Methane Production Assay
2.4. Isoprene Production Assay
2.5. Municipal Wastewater Handling
3. Results
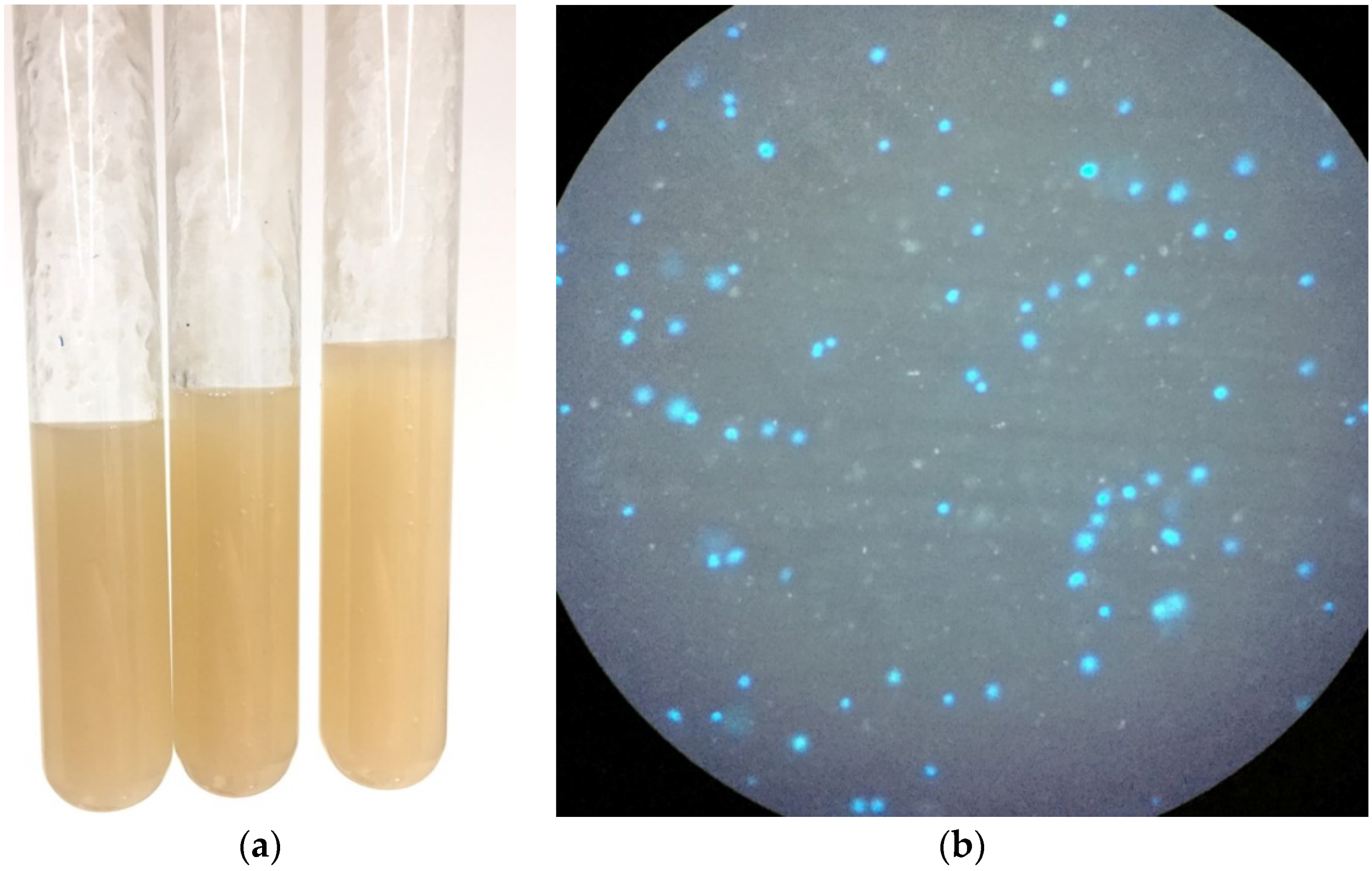
3.1. Adapting Methanosarcina acetivorans to Growth in Wastewater
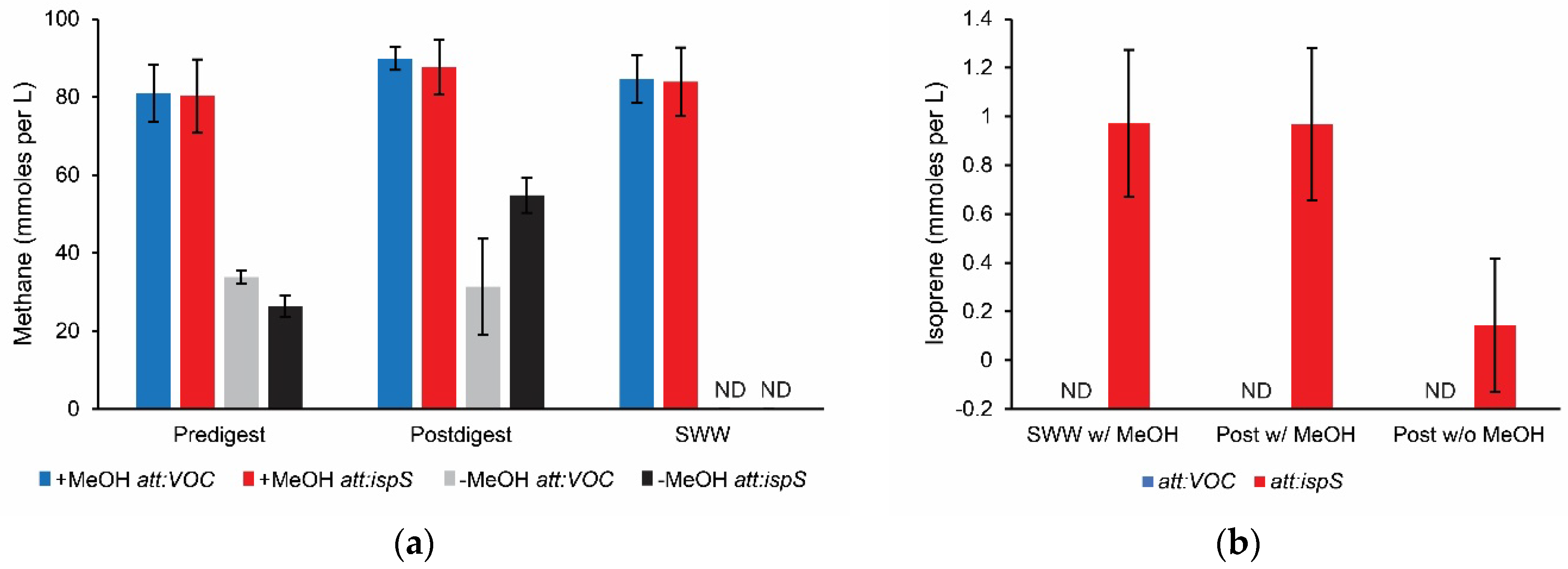
3.2. Measurement of Methane and Isoprene Production on Wastewater
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zinder, S.H. Physiological ecology of methanogens. In Methanogenesis; Springer: Boston, MA, USA, 1993; pp. 128–206. [Google Scholar] [CrossRef]
- Thauer, R.K.; Kaster, A.-K.; Seedorf, H.; Buckel, W.; Hedderich, R. Methanogenic archaea: Ecologically relevant differences in energy conservation. Nat. Rev. Microbiol. 2008, 6, 579–591. [Google Scholar] [CrossRef]
- Conrad, R. Microbial ecology of methanogens and methanotrophs. Adv. Agron. 2007, 96, 1–63. [Google Scholar]
- Lyu, Z.; Shao, N.; Akinyemi, T.; Whitman, W.B. Methanogenesis. Curr. Biol. 2018, 28, R727–R732. [Google Scholar] [CrossRef] [PubMed]
- Schiraldi, C.; Giuliano, M.; De Rosa, M. Perspectives on biotechnological applications of archaea. Archaea 2002, 1, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Tabatabaei, M.; Rahim, R.A.; Abdullah, N.; Wright, A.-D.G.; Shirai, Y.; Sakai, K.; Sulaiman, A.; Hassan, M.A. Importance of the methanogenic archaea populations in anaerobic wastewater treatments. Process Biochem. 2010, 45, 1214–1225. [Google Scholar] [CrossRef]
- Chen, S.; Cheng, H.; Liu, J.; Hazen, T.C.; Huang, V.; He, Q. Unexpected competitiveness of Methanosaeta populations at elevated acetate concentrations in methanogenic treatment of animal wastewater. Appl. Microbiol. Biotechnol. 2017, 101, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Ferry, J.G.; Maranas, C.D.; Wood, T.K. Methane-to-Acetate Pathway for Producing Liquid Biofuels and Biorenewables. U.S. Patent 20150147791A1, 28 May 2015. [Google Scholar]
- Aldridge, J.; Carr, S.; Weber, K.A.; Buan, N.R. Anaerobic production of isoprene by engineered Methanosarcina spp. archaea. Appl. Environ. Microbiol. 2021. [Google Scholar] [CrossRef]
- Elalami, D.; Carrere, H.; Monlau, F.; Abdelouahdi, K.; Oukarroum, A.; Barakat, A. Pretreatment and co-digestion of wastewater sludge for biogas production: Recent research advances and trends. Renew. Sustain. Energy Rev. 2019, 114, 109287. [Google Scholar] [CrossRef]
- Vítězová, M.; Kohoutová, A.; Vítěz, T.; Hanišáková, N.; Kushkevych, I. Methanogenic Microorganisms in Industrial Wastewater Anaerobic Treatment. Processes 2020, 8, 1546. [Google Scholar] [CrossRef]
- Lettinga, G. Anaerobic digestion and wastewater treatment systems. Antonie Leeuwenhoek 1995, 67, 3–28. [Google Scholar] [CrossRef]
- Sekiguchi, Y.; Kamagata, Y.; Harada, H. Recent advances in methane fermentation technology. Curr. Opin. Biotechnol. 2001, 12, 277–282. [Google Scholar] [CrossRef]
- McCarty, P.L.; Bae, J.; Kim, J. Domestic wastewater treatment as a net energy producer—Can this be achieved? Environ. Sci. Technol. 2011. [Google Scholar] [CrossRef] [PubMed]
- Van Lier, J.B.; Mahmoud, N.; Zeeman, G. Anaerobic wastewater treatment. In Biological Wastewater Treatment: Principles, Modelling and Design; IWA Publishing: London, UK, 2008; pp. 415–456. [Google Scholar]
- Dionisi, D. Biological Wastewater Treatment Processes: Mass and Heat Balances; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Galagan, J.E.; Nusbaum, C.; Roy, A.; Endrizzi, M.G.; Macdonald, P.; FitzHugh, W.; Calvo, S.; Engels, R.; Smirnov, S.; Atnoor, D. The genome of M. acetivorans reveals extensive metabolic and physiological diversity. Genome Res. 2002, 12, 532–542. [Google Scholar] [CrossRef] [PubMed]
- Ferry, J.G. Methanosarcina acetivorans: A Model for Mechanistic Understanding of Aceticlastic and Reverse Methanogenesis. Front. Microbiol. 2020, 11, 1806. [Google Scholar] [CrossRef]
- Sowers, K.R.; Boone, J.E.; Gunsalus, R.P. Disaggregation of Methanosarcina spp. and Growth as Single Cells at Elevated Osmolarity. Appl. Environ. Microbiol. 1993, 59, 3832–3839. [Google Scholar] [CrossRef]
- Metcalf, W.W.; Zhang, J.K.; Apolinario, E.; Sowers, K.R.; Wolfe, R.S. A genetic system for Archaea of the genus Methanosarcina: Liposome-mediated transformation and construction of shuttle vectors. Proc. Natl. Acad. Sci. USA 1997, 94, 2626–2631. [Google Scholar] [CrossRef]
- Zhang, J.K.; White, A.K.; Kuettner, H.C.; Boccazzi, P.; Metcalf, W.W. Directed mutagenesis and plasmid-based complementation in the methanogenic archaeon Methanosarcina acetivorans C2A demonstrated by genetic analysis of proline biosynthesis. J. Bacteriol. 2002, 184, 1449–1454. [Google Scholar] [CrossRef]
- Aldridge, J.; Carr, S.; Weber, K.A.; Buan, N.R. Production of Bioisoprene from Wastewater; NTRY6R14/4822; The Water Research Foundation: Denver, CO, USA, 2018. [Google Scholar]
- OECD. Test No. 314: Simulation Tests to Assess the Biodegradability of Chemicals Discharged in Wastewater; OECD: Paris, France, 2008. [Google Scholar]
- Aldridge, J.T.; Catlett, J.L.; Smith, M.L.; Buan, N.R. Methods for Detecting Microbial Methane Production and Consumption by Gas Chromatography. Bio Protoc. 2016, 6, e1779. [Google Scholar] [CrossRef]
- Pade, N.; Erdmann, S.; Enke, H.; Dethloff, F.; Duhring, U.; Georg, J.; Wambutt, J.; Kopka, J.; Hess, W.R.; Zimmermann, R.; et al. Insights into isoprene production using the cyanobacterium Synechocystis sp. PCC 6803. Biotechnol. Biofuels 2016, 9, 89. [Google Scholar] [CrossRef]
- Yang, C.; Gao, X.; Jiang, Y.; Sun, B.; Gao, F.; Yang, S. Synergy between methylerythritol phosphate pathway and mevalonate pathway for isoprene production in Escherichia coli. Metab. Eng. 2016, 37, 79–91. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Zhou, P.; Su, B.; Su, S.; Ye, L.; Yu, H. Enhanced Isoprene Production by Reconstruction of Metabolic Balance between Strengthened Precursor Supply and Improved Isoprene Synthase in Saccharomyces cerevisiae. ACS Synth. Biol. 2018, 7, 2308–2316. [Google Scholar] [CrossRef]
- Cabezas, A.; de Araujo, J.C.; Callejas, C.; Galès, A.; Hamelin, J.; Marone, A.; Sousa, D.Z.; Trably, E.; Etchebehere, C. How to use molecular biology tools for the study of the anaerobic digestion process? Rev. Environ. Sci. Bio/Technol. 2015, 14, 555–593. [Google Scholar] [CrossRef]
- Weiss, A.; Jérôme, V.; Freitag, R.; Mayer, H.K. Diversity of the resident microbiota in a thermophilic municipal biogas plant. Appl. Microbiol. Biotechnol. 2008, 81, 163–173. [Google Scholar] [CrossRef]
- Karakashev, D.; Batstone, D.J.; Angelidaki, I. Influence of environmental conditions on methanogenic compositions in anaerobic biogas reactors. Appl. Environ. Microbiol. 2005, 71, 331–338. [Google Scholar] [CrossRef]
- Ozgun, H.; Tao, Y.; Ersahin, M.E.; Zhou, Z.; Gimenez, J.B.; Spanjers, H.; van Lier, J.B. Impact of temperature on feed-flow characteristics and filtration performance of an upflow anaerobic sludge blanket coupled ultrafiltration membrane treating municipal wastewater. Water Res. 2015, 83, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Catlett, J.L.; Catazaro, J.; Cashman, M.; Carr, S.; Powers, R.; Cohen, M.B.; Buan, N.R. Metabolic Feedback Inhibition Influences Metabolite Secretion by the Human Gut Symbiont Bacteroides thetaiotaomicron. mSystems 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- McInerney, M.J.; Sieber, J.R.; Gunsalus, R.P. Microbial syntrophy: Ecosystem-level biochemical cooperation. Microbe Mag. 2011, 6, 479–485. [Google Scholar] [CrossRef][Green Version]
- Sieber, J.R.; McInerney, M.J.; Gunsalus, R.P. Genomic insights into syntrophy: The paradigm for anaerobic metabolic cooperation. Annu. Rev. Microbiol. 2012, 66, 429–452. [Google Scholar] [CrossRef]
- Worm, P.; Koehorst, J.J.; Visser, M.; Sedano-Núñez, V.T.; Schaap, P.J.; Plugge, C.M.; Sousa, D.Z.; Stams, A.J. A genomic view on syntrophic versus non-syntrophic lifestyle in anaerobic fatty acid degrading communities. Biochim. Biophys. Acta BBA Bioenerg. 2014, 1837, 2004–2016. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Synthetic Waste Water | Predigest Effluent | Postdigest Effluent | |||||
|---|---|---|---|---|---|---|---|
| CH4 a Production | |||||||
| Substrate | Strain b | mmol L−1 | Std Dev | mmol L−1 | Std Dev | mmol L−1 | Std Dev |
| MeOH c | att:VOC | 84.64 | 6.19 | 80.98 | 7.34 | 89.87 | 2.94 |
| att:ispS | 83.89 | 8.32 | 80.22 | 9.42 | 87.63 | 6.98 | |
| None | att:VOC | NT | 33.85 | 1.64 | 31.36 | 12.30 | |
| att:ispS | NT | 26.36 | 2.74 | 54.82 | 4.57 | ||
| Heat killed | att:VOC | ND | ND | ND | |||
| att:ispS | ND | ND | ND | ||||
| Isoprene a Production | |||||||
| Substrate | Strain | mmol L−1 | Std Dev | mmol L−1 | Std Dev | mmol L−1 | Std Dev |
| MeOH c | att:VOC | ND | ND | ND | |||
| att:ispS | 0.972 | 0.301 | ND | 0.968 | 0.312 | ||
| None | att:VOC | NT | ND | ND | |||
| att:ispS | NT | ND | 0.144 | 0.273 | |||
| Heat killed | att:VOC | ND | ND | ND | |||
| att:ispS | ND | ND | ND | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carr, S.; Aldridge, J.; Buan, N.R. Isoprene Production from Municipal Wastewater Biosolids by Engineered Archaeon Methanosarcina acetivorans. Appl. Sci. 2021, 11, 3342. https://doi.org/10.3390/app11083342
Carr S, Aldridge J, Buan NR. Isoprene Production from Municipal Wastewater Biosolids by Engineered Archaeon Methanosarcina acetivorans. Applied Sciences. 2021; 11(8):3342. https://doi.org/10.3390/app11083342
Chicago/Turabian StyleCarr, Sean, Jared Aldridge, and Nicole R. Buan. 2021. "Isoprene Production from Municipal Wastewater Biosolids by Engineered Archaeon Methanosarcina acetivorans" Applied Sciences 11, no. 8: 3342. https://doi.org/10.3390/app11083342
APA StyleCarr, S., Aldridge, J., & Buan, N. R. (2021). Isoprene Production from Municipal Wastewater Biosolids by Engineered Archaeon Methanosarcina acetivorans. Applied Sciences, 11(8), 3342. https://doi.org/10.3390/app11083342