
Influences of Compressive Force and Zoledronic Acid on Osteoblast Proliferation and Differentiation: An In Vitro Study

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cell Proliferation Assay and Morphological Analysis
2.3. Osteoblast Differentiation, ZA Treatment and Compressive Force Application
2.4. cDNA Preparation and Real-Time Polymerase Chain Reaction (PCR)
2.5. Matrix Mineralization
2.6. Statistical Analysis
3. Results
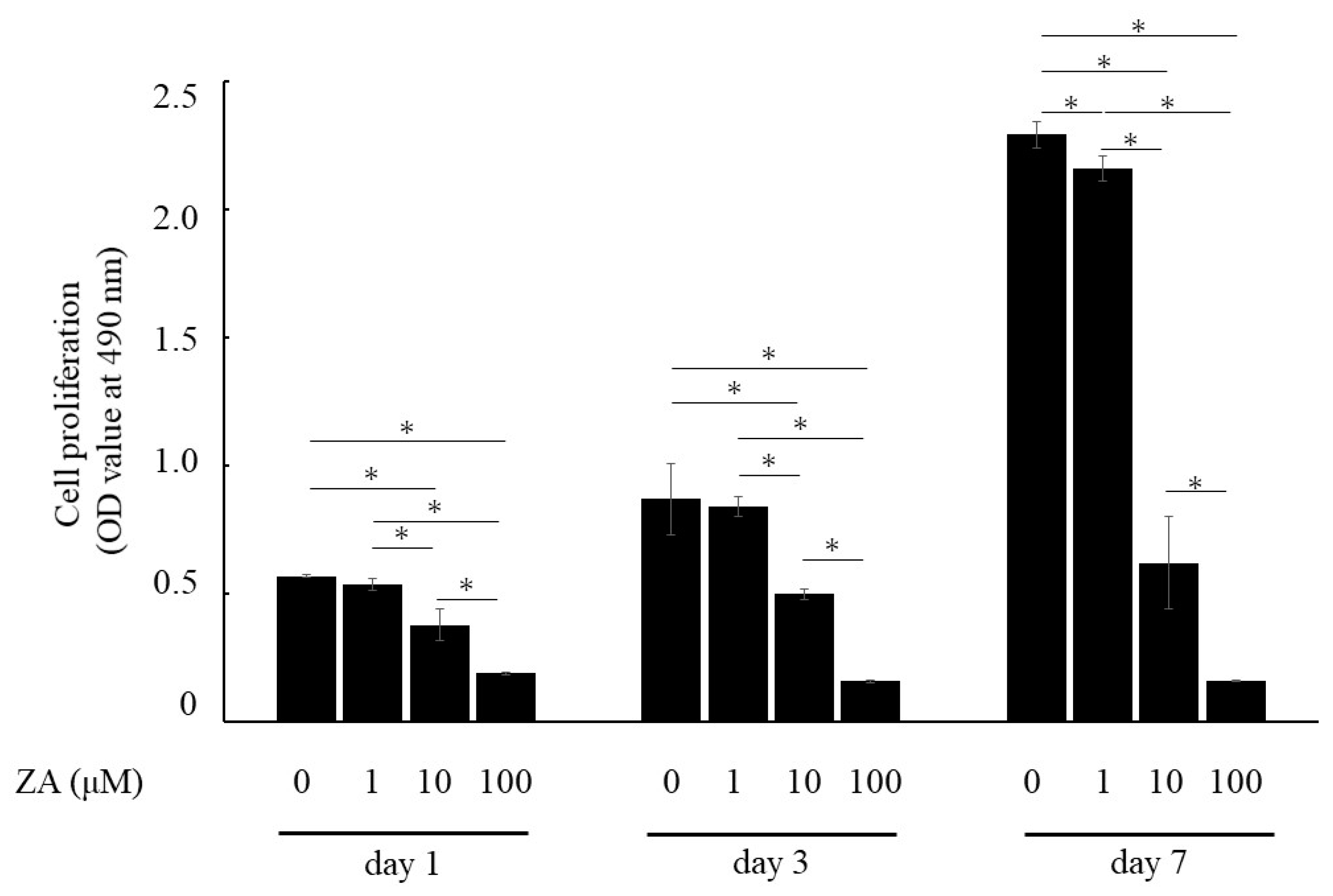
3.1. Cell Proliferation and Cell Morphology
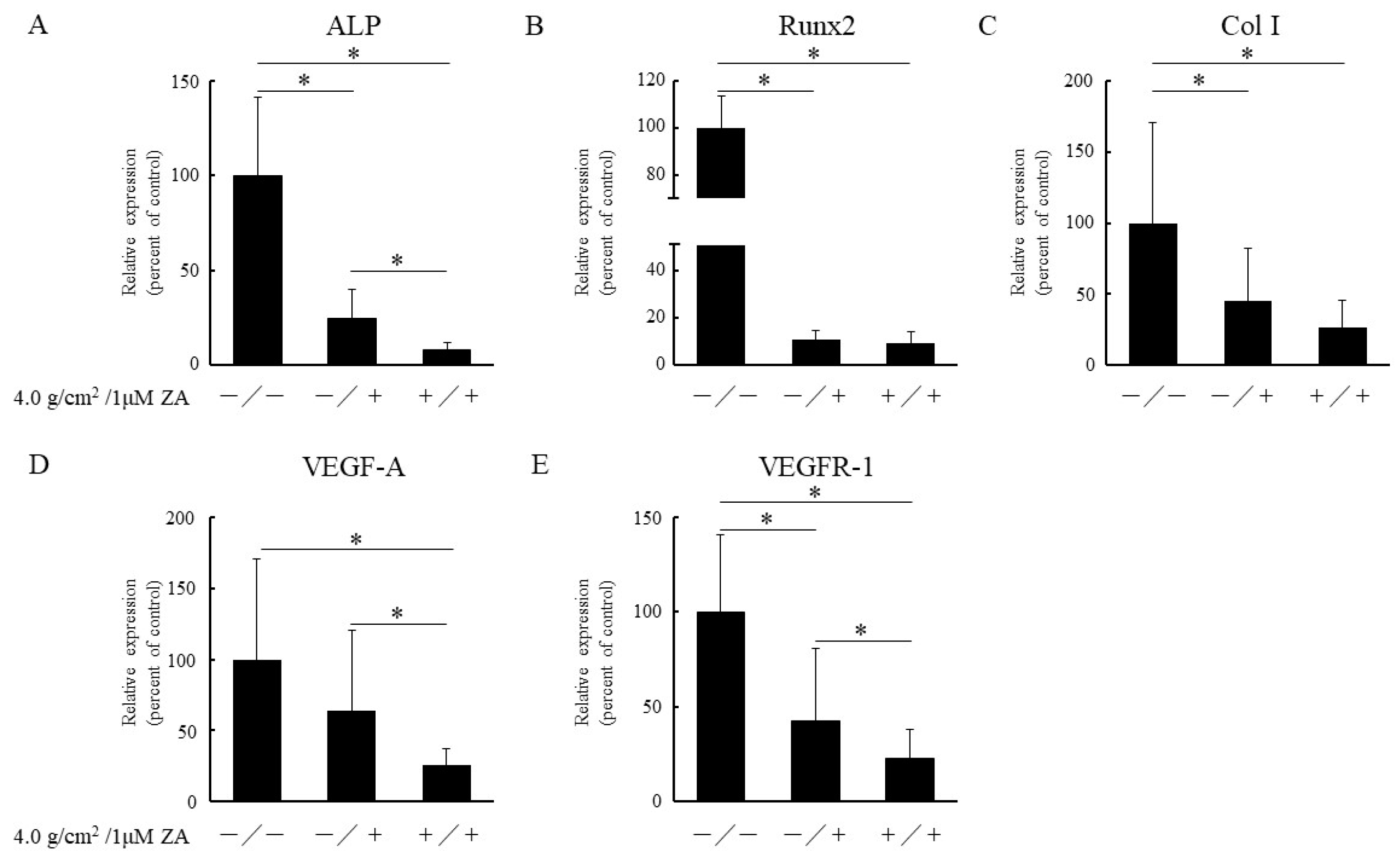
3.2. Real-Time PCR Analysis of Osteoblast-Related Genes
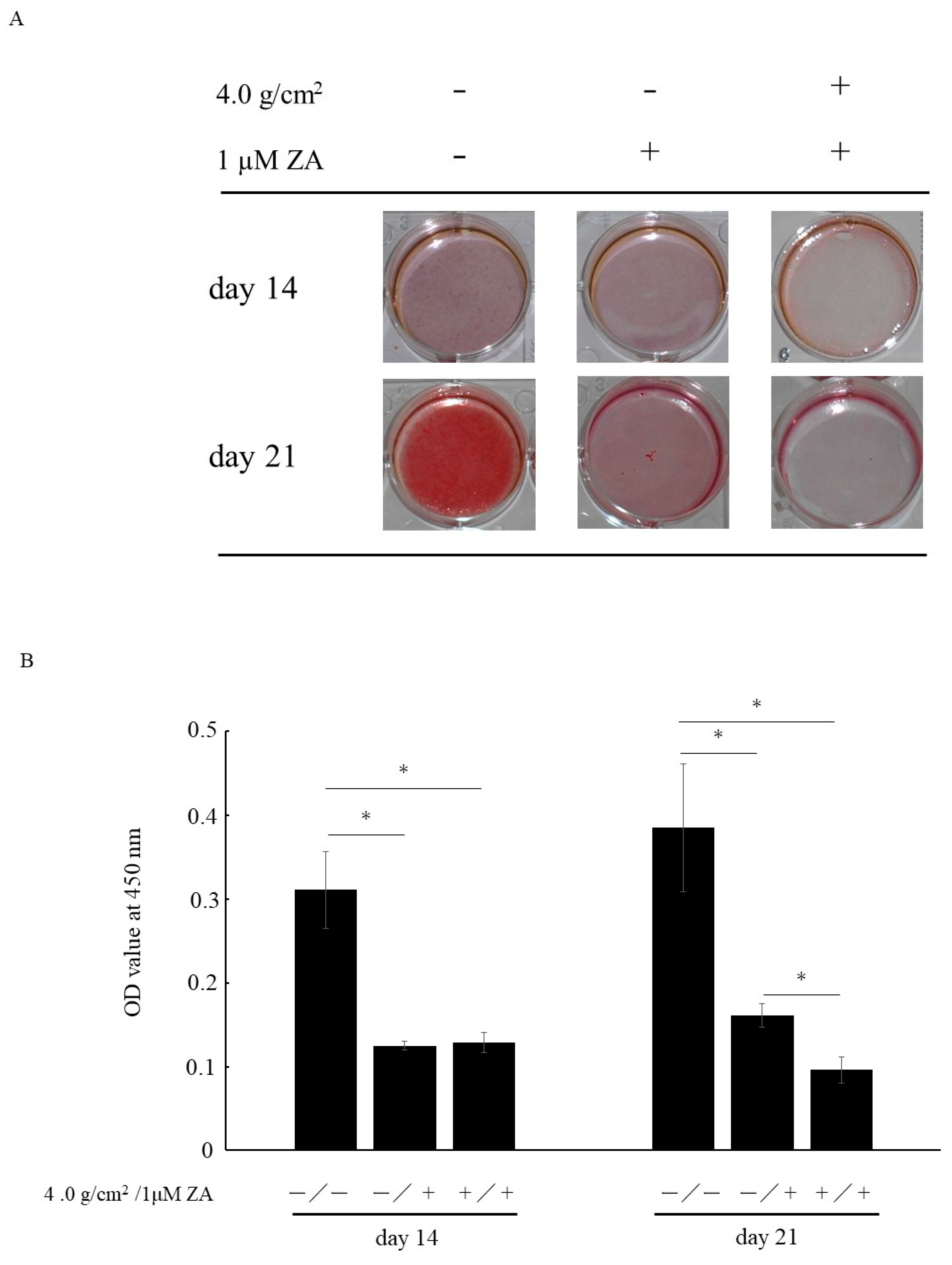
3.3. Matrix Mineralization
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bone, H.G.; Hosking, D.; Devogelaer, J.P.; Tucci, J.R.; Emkey, R.D.; Tonino, R.P.; Rodriguez-Portales, J.A.; Downs, R.W.; Gupta, J.; Santora, A.C.; et al. Alendronate Phase III Osteoporosis Treatment Study Group. Ten years’ experience with alendronate for osteoporosis in postmenopausal women. N. Engl. J. Med. 2004, 350, 1189–1199. [Google Scholar] [CrossRef] [Green Version]
- Aapro, M.; Abrahamsson, P.A.; Body, J.J.; Coleman, R.E.; Colomer, R.; Costa, L.; Crinò, L.; Dirix, L.; Gnant, M.; Gralow, J.; et al. Guidance on the use of bisphosphonates in solid tumours: Recommendations of an international expert panel. Ann. Oncol. 2008, 19, 420–432. [Google Scholar] [CrossRef]
- Ohlrich, E.J.; Coates, D.E.; Cullinan, M.P.; Milne, T.J.; Zafar, S.; Zhao, Y.; Duncan, W.D.; Seymour, G.J. The bisphosphonate zoledronic acid regulates key angiogenesis-related genes in primary human gingival fibroblasts. Arch. Oral. Biol. 2016, 63, 7–14. [Google Scholar] [CrossRef]
- Ruggiero, S.L.; Dodson, T.B.; Fantasia, J.; Goodday, R.; Aghaloo, T.; Mehrotra, B.; O’Ryan, F. American Association of Oral and Maxillofacial Surgeons. American Association of Oral and Maxillofacial Surgeons position paper on medication-related osteonecrosis of the jaw—2014 update. J. Oral Maxillofac. Surg. 2014, 72, 1938–1956. [Google Scholar] [CrossRef]
- Gong, X.; Yu, W.; Zhao, H.; Su, J.; Sheng, Q. Skeletal Site-specific Effects of Zoledronate on in vivo Bone Remodeling and in vitro BMSCs Osteogenic Activity. Sci. Rep. 2017, 7, 36129. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, T.; Yamori, M.; Ishizaki, T.; Asai, K.; Goto, K.; Takahashi, K.; Nakayama, T.; Bessho, K. Increased incidence of osteonecrosis of the jaw after tooth extraction in patients treated with bisphosphonates: A cohort study. Int. J. Oral Maxillofac. Surg. 2012, 41, 1397–1403. [Google Scholar]
- Peer, A.; Khamaisi, M. Diabetes as a risk factor for medication-related osteonecrosis of the jaw. J. Dent. Res. 2015, 94, 252–260. [Google Scholar]
- Soutome, S.; Hayashida, S.; Funahara, M.; Sakamoto, Y.; Kojima, Y.; Yanamoto, S.; Umeda, M. Factors affecting development of medication-related osteonecrosis of the jaw in cancer patients receiving high-dose bisphosphonate or denosumab therapy: Is tooth extraction a risk factor? PLoS ONE 2018, 13, e0201343. [Google Scholar] [CrossRef] [Green Version]
- Ichimiya, H.; Takahashi, T.; Ariyoshi, W.; Takano, H.; Matayoshi, T.; Nishihara, T. Compressive mechanical stress promotes osteoclast formation through RANKL expression on synovial cells. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2007, 103, 334–341. [Google Scholar]
- Pozzi, S.; Vallet, S.; Mukherjee, S.; Cirstea, D.; Vaghela, N.; Santo, L.; Rosen, E.; Ikeda, H.; Okawa, Y.; Kiziltepe, T.; et al. High-dose zoledronic acid impacts bone remodeling with effects on osteoblastic lineage and bone mechanical properties. Clin. Cancer Res. 2009, 15, 5829–5839. [Google Scholar] [CrossRef] [Green Version]
- Patntirapong, S.; Singhatanadgit, W.; Chanruangvanit, C.; Lavanrattanakul, K.; Satravaha, Y. Zoledronic acid suppresses mineralization through direct cytotoxicity and osteoblast differentiation inhibition. J. Oral Pathol. Med. 2012, 41, 713–720. [Google Scholar] [CrossRef]
- Huang, K.C.; Cheng, C.C.; Chuang, P.Y.; Yang, T.Y. The effects of zoledronate on the survival and function of human osteoblast-like cells. BMC Musculoskelet. Disord. 2015, 16, 355. [Google Scholar] [CrossRef] [Green Version]
- Pons-Fuster López, E.; Seoane Leston, J.; López Jornet, P. Epigallocatechin-3-gallate reduces damage to osteoblast-like cells treated with Zoledronic acid. Arch. Oral Biol. 2018, 94, 27–32. [Google Scholar] [CrossRef]
- Komori, T. Regulation of bone development and extracellular matrix protein genes by RUNX2. Cell Tissue Res. 2010, 339, 189–195. [Google Scholar] [CrossRef] [Green Version]
- Robling, A.G.; Castillo, A.B.; Turner, C.H. Biomechanical and molecular regulation of bone remodeling. Annu. Rev. Biomed. Eng. 2006, 8, 455–498. [Google Scholar] [CrossRef]
- Gerber, H.P.; Vu, T.H.; Ryan, A.M.; Kowalski, J.; Werb, Z.; Ferrara, N. VEGF couples hypertrophic cartilage remodeling, ossification and angiogenesis during endochondral bone formation. Nat. Med. 1999, 5, 623–628. [Google Scholar] [CrossRef]
- Jacobsen, K.A.; Al-Aql, Z.S.; Wan, C.; Fitch, J.L.; Stapleton, S.N.; Mason, Z.D.; Cole, R.M.; Gilbert, S.R.; Clemens, T.L.; Morgan, E.F.; et al. Bone formation during distraction osteogenesis is dependent on both VEGFR1 and VEGFR2 signaling. J. Bone Miner. Res. 2008, 23, 596–609. [Google Scholar] [CrossRef] [Green Version]
- Deckers, M.M.; Karperien, M.; van der Bent, C.; Yamashita, T.; Papapoulos, S.E.; Löwik, C.W. Expression of vascular endothelial growth factors and their receptors during osteoblast differentiation. Endocrinology 2000, 141, 1667–1674. [Google Scholar] [CrossRef]
- Schipani, E.; Maes, C.; Carmeliet, G.; Semenza, G.L. Regulation of osteogenesis-angiogenesis coupling by HIFs and VEGF. J. Bone. Miner. Res. 2009, 24, 1347–1353. [Google Scholar] [CrossRef]
- Maruotti, N.; Corrado, A.; Neve, A.; Cantatore, F.P. Bisphosphonates: Effects on osteoblast. Eur. J. Clin. Pharmacol. 2012, 68, 1013–1018. [Google Scholar] [CrossRef]
- Yang, X.; Lu, Y.; Li, Z.; Wang, Y.; Zhao, F.; Han, J. Low concentrations of zoledronic acid are better at regulating bone formation and repair. Intractable Rare Dis. Res. 2013, 2, 18–23. [Google Scholar] [CrossRef] [Green Version]
- Svanberg, G.; Lindhe, J. Vascular reactions in the periodontal ligament incident to trauma from occlusion. J. Clin. Periodontol. 1974, 1, 58–69. [Google Scholar] [CrossRef]
- Branschofsky, M.; Beikler, T.; Schäfer, R.; Flemming, T.F.; Lang, H. Secondary trauma from occlusion and periodontitis. Quintessence Int. 2011, 42, 515–522. [Google Scholar]
- Naert, I.; Duyck, J.; Vandamme, K. Occlusal overload and bone/implant loss. Clin. Oral Implants Res. 2012, 23, 95–107. [Google Scholar] [CrossRef]
- Yanagisawa, M.; Suzuki, N.; Mitsui, N.; Koyama, Y.; Otsuka, K.; Shimizu, N. Compressive force stimulates the expression of osteogenesis-related transcription factors in ROS 17/2.8 cells. Arch. Oral Biol. 2008, 53, 214–219. [Google Scholar] [CrossRef]
- Mitsui, N.; Suzuki, N.; Koyama, Y.; Yanagisawa, M.; Otsuka, K.; Shimizu, N.; Maeno, M. Effect of compressive force on the expression of MMPs, PAs, and their inhibitors in osteoblastic Saos-2 cells. Life Sci. 2006, 79, 575–583. [Google Scholar] [CrossRef]
- Mori, S.; Sato, T.; Hara, T.; Nakashima, K.; Minagi, S. Effect of continuous pressure on histopathological changes in denture-supporting tissues. J. Oral Rehabil. 1997, 24, 37–46. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yusa, K.; Ishikawa, S.; Hemmi, T.; Takano, H.; Fukuda, M.; Iino, M. Influences of Compressive Force and Zoledronic Acid on Osteoblast Proliferation and Differentiation: An In Vitro Study. Appl. Sci. 2021, 11, 11273. https://doi.org/10.3390/app112311273
Yusa K, Ishikawa S, Hemmi T, Takano H, Fukuda M, Iino M. Influences of Compressive Force and Zoledronic Acid on Osteoblast Proliferation and Differentiation: An In Vitro Study. Applied Sciences. 2021; 11(23):11273. https://doi.org/10.3390/app112311273
Chicago/Turabian StyleYusa, Kazuyuki, Shigeo Ishikawa, Tomoharu Hemmi, Hiroshi Takano, Masayuki Fukuda, and Mitsuyoshi Iino. 2021. "Influences of Compressive Force and Zoledronic Acid on Osteoblast Proliferation and Differentiation: An In Vitro Study" Applied Sciences 11, no. 23: 11273. https://doi.org/10.3390/app112311273
APA StyleYusa, K., Ishikawa, S., Hemmi, T., Takano, H., Fukuda, M., & Iino, M. (2021). Influences of Compressive Force and Zoledronic Acid on Osteoblast Proliferation and Differentiation: An In Vitro Study. Applied Sciences, 11(23), 11273. https://doi.org/10.3390/app112311273