
Recombinant Human Erythropoietin Production in Chinese Hamster Ovary Cells Is Enhanced by Supplementation of α-Helix Domain of 30Kc19 Protein


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plasmid Construction and Production of Recombinant Protein in E. coli Expression System
2.2. Cell Culture
2.3. EPO Production from CHO Cells
2.4. Enzyme-Linked Immunosorbent Assay (ELISA)
2.5. Immunofluorescence Analysis
2.6. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS–PAGE) and Immunoblot Analysis
2.7. NADPH/NADP+ Quantitative Assay
2.8. Statistical Analysis
3. Results and Discussion
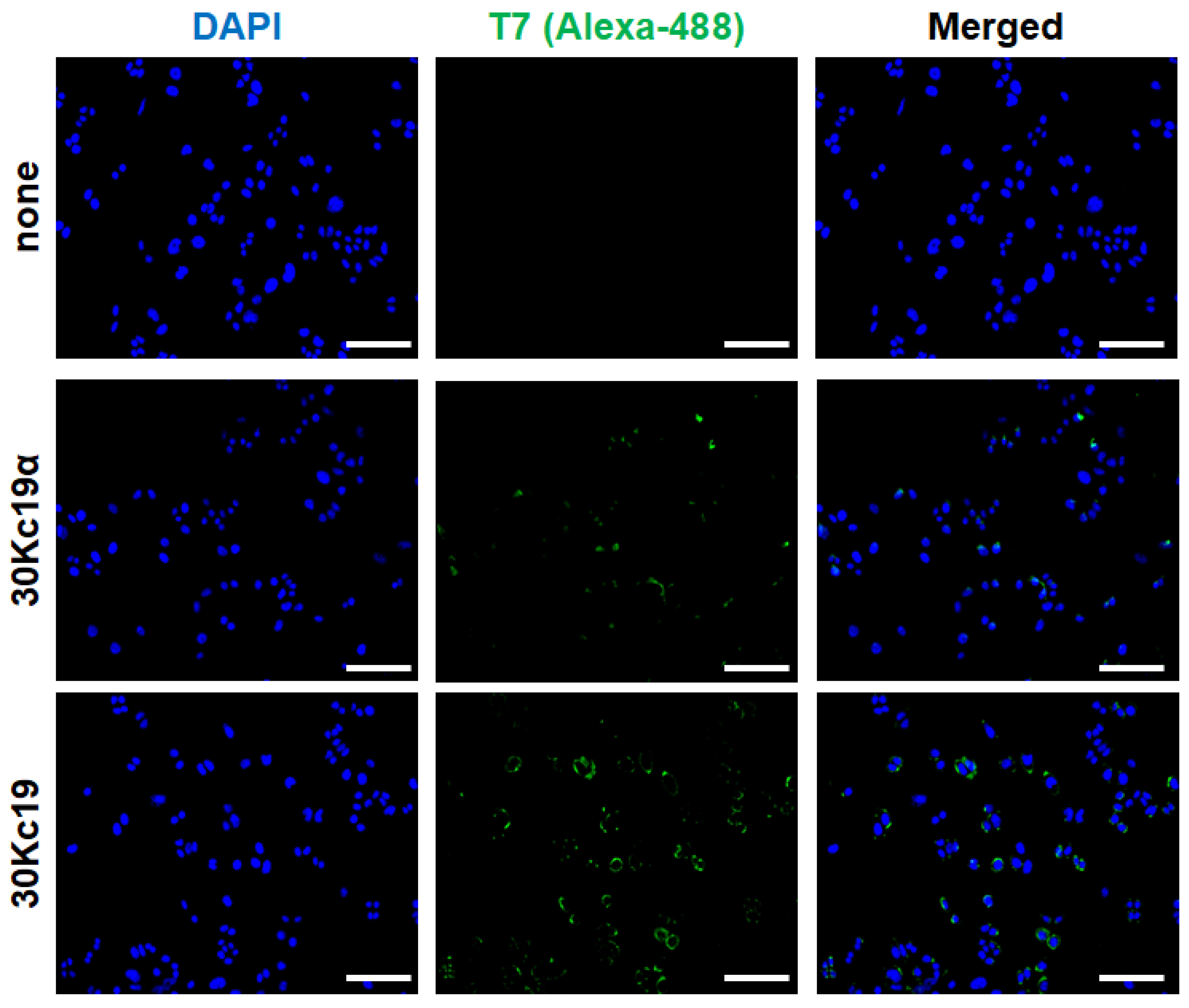
3.1. 30Kc19α Protein Expression and Its Intracellular Delivery
3.2. Effect of 30Kc19α on Cell Growth and Viability
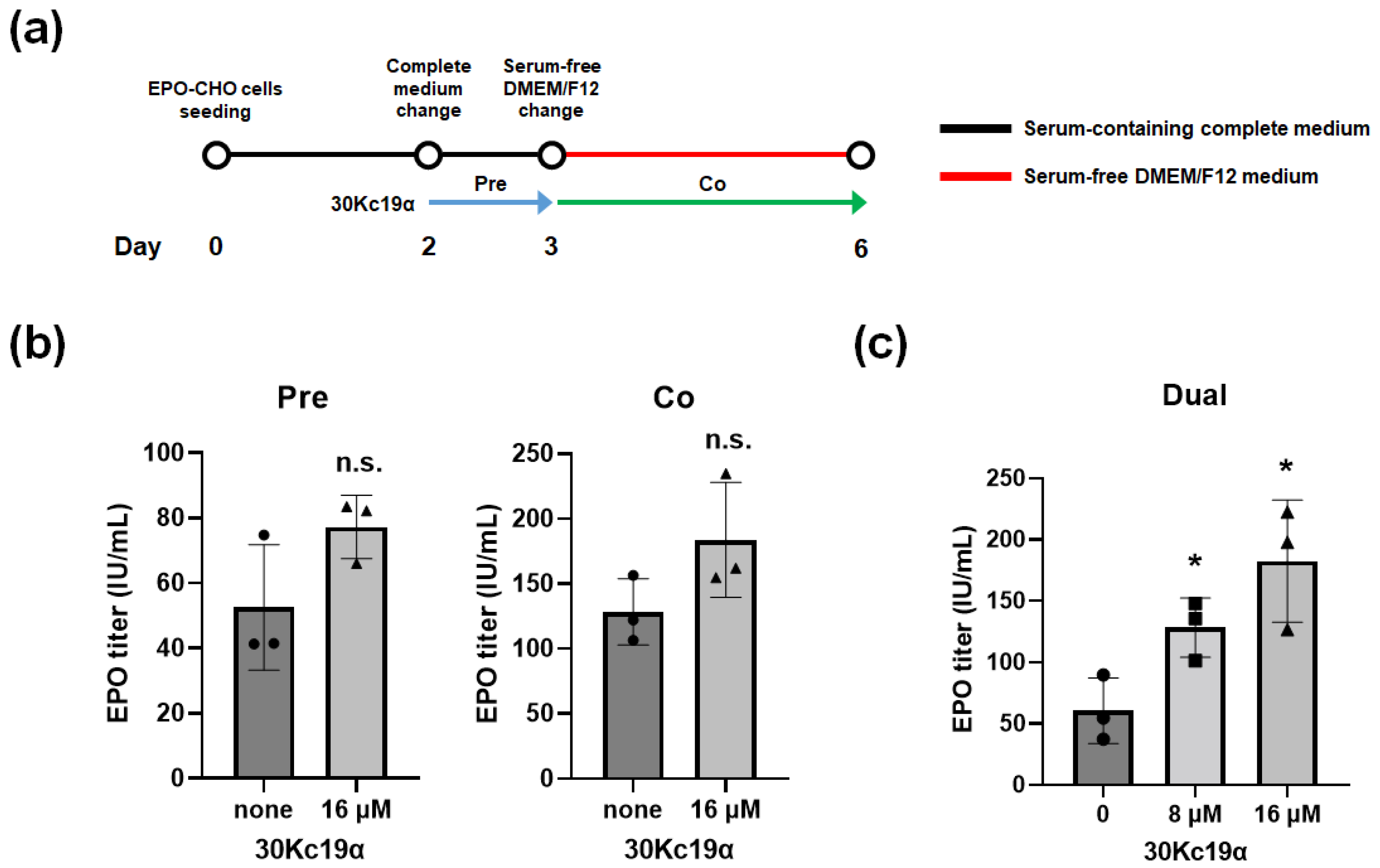
3.3. Improvement of EPO Production
3.4. Effect of 30Kc19α on NADPH/NADP+ Ratio of EPO–CHO Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Johnson, I.S. Human insulin from recombinant DNA technology. Science 1983, 219, 632–637. [Google Scholar] [CrossRef]
- Kesik-Brodacka, M. Progress in biopharmaceutical development. Biotechnol. Appl. Biochem. 2018, 65, 306–322. [Google Scholar] [CrossRef] [PubMed]
- Pavlou, A.K.; Reichert, J.M. Recombinant protein therapeutics—success rates, market trends and values to 2010. Nat. Biotechnol. 2004, 22, 1513–1519. [Google Scholar] [CrossRef]
- Wurm, F.M. Production of recombinant protein therapeutics in cultivated mammalian cells. Nat. Biotechnol. 2004, 22, 1393–1398. [Google Scholar] [CrossRef] [PubMed]
- Omasa, T.; Onitsuka, M.; Kim, W.-D. Cell engineering and cultivation of Chinese hamster ovary (CHO) cells. Curr. Pharm. Biotechnol. 2010, 11, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, Y.-G.; Lee, G.M. CHO cells in biotechnology for production of recombinant proteins: Current state and further potential. Appl. Microbiol. Biotechnol. 2012, 93, 917–930. [Google Scholar] [CrossRef] [PubMed]
- Hossler, P.; Khattak, S.F.; Li, Z.J. Optimal and consistent protein glycosylation in mammalian cell culture. Glycobiology 2009, 19, 936–949. [Google Scholar] [CrossRef] [PubMed]
- Datta, P.; Linhardt, R.J.; Sharfstein, S.T. An’omics approach towards CHO cell engineering. Biotechnol. Bioeng. 2013, 110, 1255–1271. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, K.; Marth, J.D. Glycosylation in cellular mechanisms of health and disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef]
- Yoon, S.K.; Song, J.Y.; Lee, G.M. Effect of low culture temperature on specific productivity, transcription level, and heterogeneity of erythropoietin in Chinese hamster ovary cells. Biotechnol. Bioeng. 2003, 82, 289–298. [Google Scholar] [CrossRef]
- Yoon, S.K.; Choi, S.L.; Song, J.Y.; Lee, G.M. Effect of culture pH on erythropoietin production by Chinese hamster ovary cells grown in suspension at 32.5 and 37.0 °C. Biotechnol. Bioeng. 2005, 89, 345–356. [Google Scholar] [CrossRef]
- Restelli, V.; Wang, M.D.; Huzel, N.; Ethier, M.; Perreault, H.; Butler, M. The effect of dissolved oxygen on the production and the glycosylation profile of recombinant human erythropoietin produced from CHO cells. Biotechnol. Bioeng. 2006, 94, 481–494. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.S.; Kim, T.K.; Chung, J.Y.; Lee, G.M. Osmoprotective effect of glycine betaine on foreign protein production in hyperosmotic recombinant Chinese hamster ovary cell cultures differs among cell lines. Biotechnol. Bioeng. 2000, 70, 167–175. [Google Scholar] [CrossRef]
- Chiang, G.G.; Sisk, W.P. Bcl-xl mediates increased production of humanized monoclonal antibodies in Chinese hamster ovary cells. Biotechnol. Bioeng. 2005, 91, 779–792. [Google Scholar] [CrossRef]
- Jaluria, P.; Betenbaugh, M.; Konstantopoulos, K.; Shiloach, J. Enhancement of cell proliferation in various mammalian cell lines by gene insertion of a cyclin-dependent kinase homolog. BMC Biotechnol. 2007, 7, 71. [Google Scholar] [CrossRef]
- Hwang, S.O.; Chung, J.Y.; Lee, G.M. Effect of doxycycline-regulated ERp57 expression on specific thrombopoietin productivity of recombinant CHO cells. Biotechnol. Prog. 2003, 19, 179–184. [Google Scholar] [CrossRef]
- Jelkmann, W. Physiology and pharmacology of erythropoietin. Transfus. Med. Hemother. 2013, 40, 302–309. [Google Scholar] [CrossRef]
- Moore, E.; Bellomo, R. Erythropoietin (EPO) in acute kidney injury. Ann. Intensive Care 2011, 1, 3. [Google Scholar] [CrossRef]
- Ji, Y.Q.; Zhang, Y.Q.; Li, M.Q.; Du, M.R.; Wei, W.W.; Li, D.J. EPO improves the proliferation and inhibits apoptosis of trophoblast and decidual stromal cells through activating STAT-5 and inactivating p38 signal in human early pregnancy. Int. J. Clin. Exp. Pathol. 2011, 4, 765–774. [Google Scholar] [PubMed]
- Sun, C.; Zhang, S.; Wang, J.; Jiang, W.; Xin, Q.; Chen, X.; Zhang, Z.; Luan, Y. EPO enhances the protective effects of MSCs in experimental hyperoxia-induced neonatal mice by promoting angiogenesis. Aging 2019, 11, 2477–2487. [Google Scholar] [CrossRef]
- Lee, J.S.; Ha, T.K.; Lee, S.J.; Lee, G.M. Current state and perspectives on erythropoietin production. Appl. Microbiol. Biotechnol. 2012, 95, 1405–1416. [Google Scholar] [CrossRef]
- Cazzola, M.; Mercuriali, F.; Brugnara, C. Use of recombinant human erythropoietin outside the setting of uremia. Blood 1997, 89, 4248–4267. [Google Scholar] [CrossRef]
- Wang, Z.; Park, J.H.; Park, H.H.; Tan, W.; Park, T.H. Enhancement of recombinant human EPO production and sialylation in Chinese hamster ovary cells through bombyx mori 30Kc19 gene expression. Biotechnol. Bioeng. 2011, 108, 1634–1642. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Wang, Z.; Jeong, H.-J.; Park, H.H.; Kim, B.-G.; Tan, W.-S.; Choi, S.S.; Park, T.H. Enhancement of recombinant human EPO production and glycosylation in serum-free suspension culture of CHO cells through expression and supplementation of 30Kc19. Appl. Microbiol. Biotechnol. 2012, 96, 671–683. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, J.H.; Park, H.H.; Rhee, W.J.; Choi, S.S.; Park, T.H. A protein delivery system using 30Kc19 cell-penetrating protein originating from silkworm. Biomaterials 2012, 33, 9127–9134. [Google Scholar] [CrossRef]
- Park, J.H.; Park, H.H.; Choi, S.S.; Park, T.H. Stabilization of enzymes by the recombinant 30Kc19 protein. Process. Biochem. 2012, 47, 164–169. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, H.J.; Park, H.H.; Rhee, W.J.; Park, T.H. Stabilization of cellular mitochondrial enzyme complex and sialyltransferase activity through supplementation of 30Kc19 protein. Appl. Microbiol. Biotechnol. 2015, 99, 2155–2163. [Google Scholar] [CrossRef]
- Ryu, J.; Kim, H.; Park, H.H.; Lee, H.J.; Park, J.H.; Rhee, W.J.; Park, T.H. Protein-stabilizing and cell-penetrating properties of α-helix domain of 30Kc19 protein. Biotechnol. J. 2016, 11, 1443–1451. [Google Scholar] [CrossRef]
- Lee, H.; Kim, S.H.L.; Yoon, H.; Ryu, J.; Park, H.H.; Hwang, N.S.; Park, T.H. Intracellular delivery of recombinant RUNX2 facilitated by cell-penetrating protein for the osteogenic differentiation of hMSCs. ACS Biomater. Sci. Eng. 2020, 6, 5202–5214. [Google Scholar] [CrossRef] [PubMed]
- Park, H.H.; Choi, J.; Lee, H.J.; Ryu, J.; Park, J.H.; Rhee, W.J.; Park, T.H. Enhancement of human erythropoietin production in Chinese hamster ovary cells through supplementation of 30Kc19-30Kc6 fusion protein. Process. Biochem. 2015, 50, 973–980. [Google Scholar] [CrossRef]
- Hebditch, M.; Carballo-Amador, M.A.; Charonis, S.; Curtis, R.; Warwicker, J. Protein–sol: A web tool for predicting protein solubility from sequence. Bioinformatics 2017, 33, 3098–3100. [Google Scholar] [CrossRef]
- Yang, C.S.; Zhang, J.; Zhang, L.; Huang, J.; Wang, Y. Mechanisms of body weight reduction and metabolic syndrome alleviation by tea. Mol. Nutr. Food Res. 2016, 60, 160–174. [Google Scholar] [CrossRef]
- Kan, B.; London, I.; Levin, D. Role of reversing factor in the inhibition of protein synthesis initiation by oxidized glutathione. J. Biol. Chem. 1988, 263, 15652–15656. [Google Scholar] [CrossRef]
- Rufino-Palomares, E.E.; Reyes-Zurita, F.J.; García-Salguero, L.; Peragón, J.; de la Higuera, M.; Lupiáñez, J.A. NADPH production, a growth marker, is stimulated by maslinic acid in gilthead sea bream by increased NADP-IDH and ME expression. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 187, 32–42. [Google Scholar] [CrossRef]
- Circu, M.L.; Aw, T.Y. Reactive oxygen species, cellular redox systems, and apoptosis. Free Radic. Biol. Med. 2010, 48, 749–762. [Google Scholar] [CrossRef]
- Itsumi, M.; Inoue, S.; Elia, A.; Murakami, K.; Sasaki, M.; Lind, E.; Brenner, D.; Harris, I.; Chio, I.; Afzal, S. Idh1 protects murine hepatocytes from endotoxin-induced oxidative stress by regulating the intracellular NADP+/NADPH ratio. Cell Death Differ. 2015, 22, 1837–1845. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cha, H.; Park, J.-H. Recombinant Human Erythropoietin Production in Chinese Hamster Ovary Cells Is Enhanced by Supplementation of α-Helix Domain of 30Kc19 Protein. Appl. Sci. 2021, 11, 11009. https://doi.org/10.3390/app112211009
Cha H, Park J-H. Recombinant Human Erythropoietin Production in Chinese Hamster Ovary Cells Is Enhanced by Supplementation of α-Helix Domain of 30Kc19 Protein. Applied Sciences. 2021; 11(22):11009. https://doi.org/10.3390/app112211009
Chicago/Turabian StyleCha, Hyeonjin, and Ju-Hyun Park. 2021. "Recombinant Human Erythropoietin Production in Chinese Hamster Ovary Cells Is Enhanced by Supplementation of α-Helix Domain of 30Kc19 Protein" Applied Sciences 11, no. 22: 11009. https://doi.org/10.3390/app112211009
APA StyleCha, H., & Park, J.-H. (2021). Recombinant Human Erythropoietin Production in Chinese Hamster Ovary Cells Is Enhanced by Supplementation of α-Helix Domain of 30Kc19 Protein. Applied Sciences, 11(22), 11009. https://doi.org/10.3390/app112211009