
Response of Sex Steroid Hormone Synthesis Substrates in Serum and Testes of Male Tilapia (Oreochromis niloticus) Exposed to Methomyl and Its Recovery Pattern

 ,
, 
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish and Chemicals
2.2. Experimental Design
2.3. Quantification of Methomyl
2.4. Sampling and Biochemical Analysis
2.5. Statistical Analysis
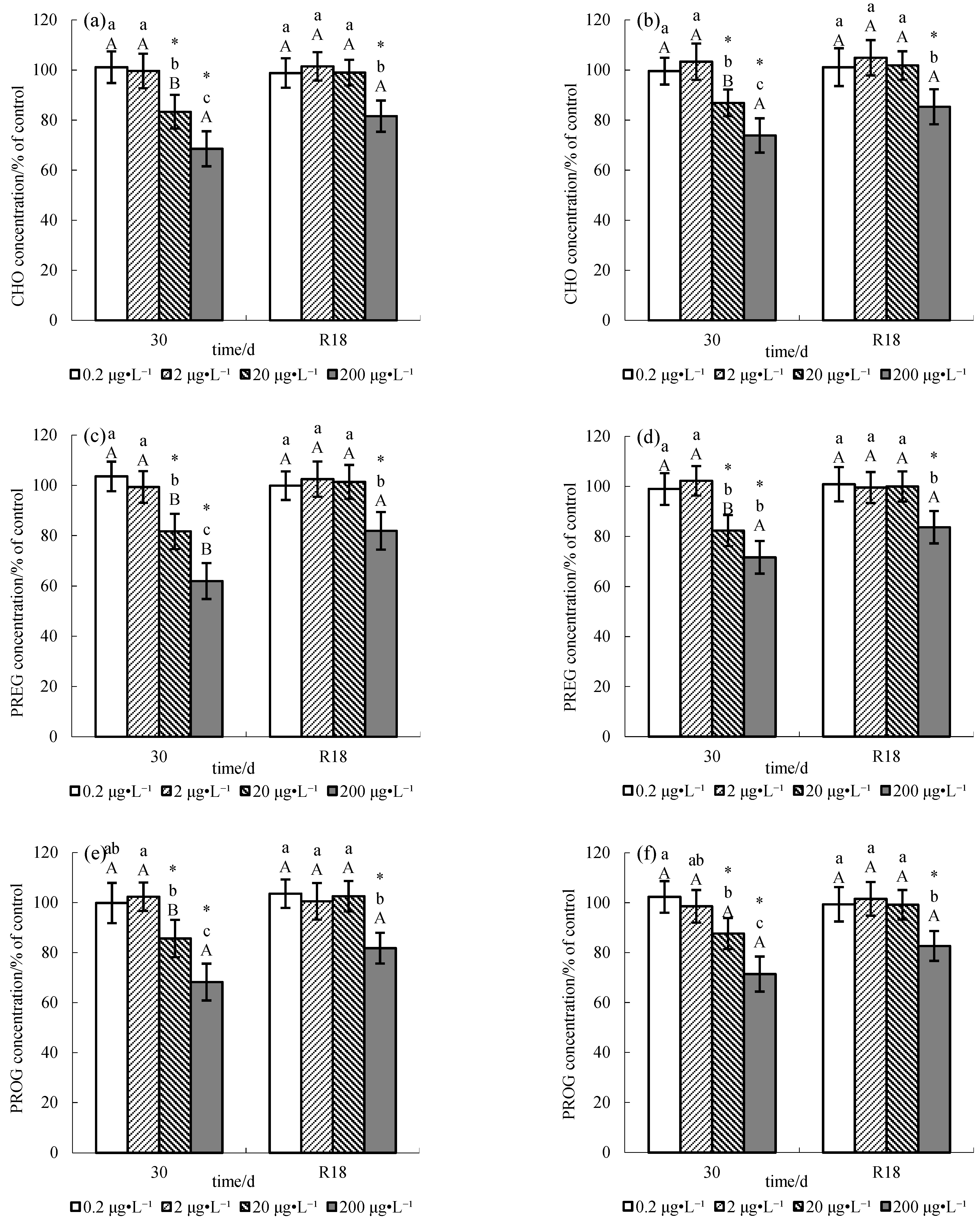
3. Results
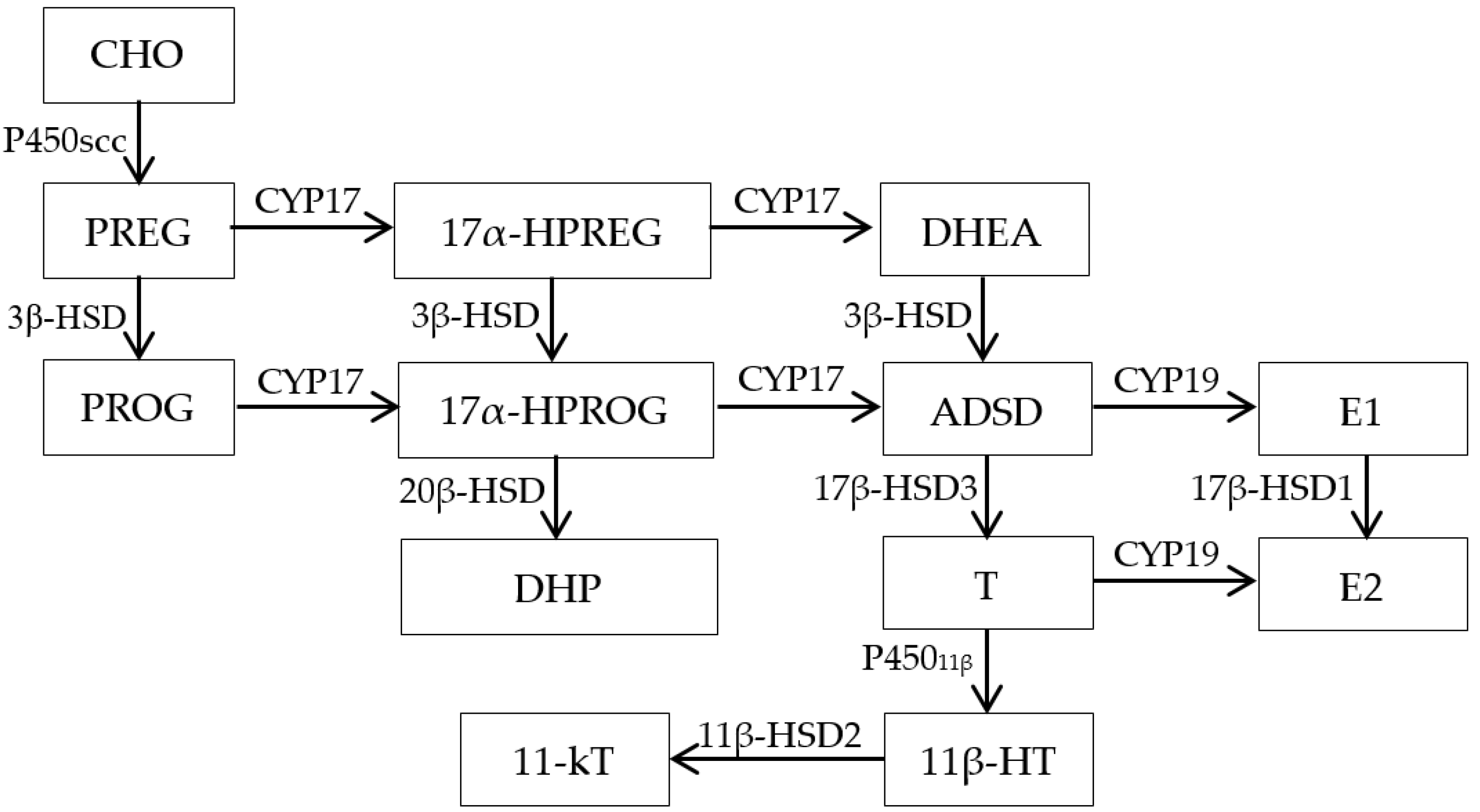
4. Discussion
4.1. Effect of Methomyl on Sex Steroid Hormone Synthesis Substrates
4.2. Recovery Pattern
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Ethic Statement
References
- Kongphonprom, K.; Burakham, R. Determination of carbamate insecticides in water, fruit, and vegetables by ultrasound-assisted dispersive liquid-liquid micro extraction and high-performance liquid chromatography. Anal. Lett. 2016, 49, 753–767. [Google Scholar] [CrossRef]
- Van Scoy, A.R.; Yue, M.; Deng, X.; Tjeerdema, R.S. Environmental fate and toxicology of methomyl. Rev. Environ. Contam. T 2013, 222, 93–109. [Google Scholar]
- Guo, X.B. Environmental Health; Peking University Medical Press: Beijing, China, 2006. [Google Scholar]
- Meng, S.L.; Hu, G.D.; Qiu, L.P.; Song, C.; Fan, L.M.; Chen, J.Z.; Xu, P. Effects of chronic exposure of methomyl on the antioxidant system in kidney of tilapia (Oreochromis niloticus) and recovery pattern. J. Toxicol. Environ. Health A 2013, 76, 937–943. [Google Scholar] [CrossRef]
- Meng, S.L.; Qiu, L.P.; Hu, G.D.; Fan, L.M.; Song, C.; Zheng, Y.; Wu, W.; Qu, J.H.; Li, D.D.; Chen, J.Z.; et al. Effects of methomyl on steroidogenic gene transcription of the hypothalamic-pituitary-gonad-liver axis in male tilapia. Chemosphere 2016, 165, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.L.; Qiu, L.P.; Hu, G.D.; Fan, L.M.; Song, C.; Zheng, Y.; Wu, W.; Qu, J.H.; Li, D.D.; Chen, J.Z.; et al. Effect of methomyl on sex steroid hormone and vitellogenin levels in serum of male tilapia (Oreochromis niloticus) and recovery pattern. Environ. Toxicol. 2017, 32, 1869–1877. [Google Scholar] [CrossRef]
- Meng, S.L.; Chen, X.; Song, C.; Fan, L.M.; Qiu, L.P.; Zheng, Y.; Chen, J.Z.; Xu, P. Effect of chronic exposure to pesticide methomyl on antioxidant defense system in testis of tilapia (Oreochromis niloticus) and its recovery pattern. Appl. Sci. 2021, 11, 3332. [Google Scholar] [CrossRef]
- Maclatchy, D.L.; Vanderkraak, G.J. The phytoestrogen β-sitosterol alters the reproductive endocrine status of goldfish-sciencedirect. Toxicol. Appl. Pharmacol. 1995, 134, 305–312. [Google Scholar] [CrossRef]
- Ibor, O.R.; Adeogun, A.O.; Chukwuka, A.V.; Arukwe, A. Gross pathology, physiological and toxicological responses in relation to metals and persistent organic pollutants (POPs) burden in tilapia species from ogun river, nigeria. Mar. Environ. Res. 2017, 129, 245–257. [Google Scholar] [CrossRef]
- Choi, J.; Lee, G.; Kim, S.; Choi, K. Investigation on sex hormone-disruption effects of two novel brominated flame retardants (DBDPE and BTBPE) in male zebrafish (Danio rerio) and two human cell lines (H295R and MVLN). Appl. Sci. 2021, 11, 3837. [Google Scholar] [CrossRef]
- Ferreira, C.; Oliveira, M.; Santos, M.A.; Pacheco, M. Effects of benzo[a]pyrene, cortisol, and 17-estradiol on liver microsomal erod activity of Anguilla anguilla: An In Vitro approach. Appl. Sci. 2021, 11, 2533. [Google Scholar] [CrossRef]
- Osman, A.G.M.; Reheem, A.E.; AbuelFadl, K.Y.; Gad El-Rab, A.G. Enzymatic and histopathologic biomarkers as indicators of aquatic pollution in fishes. Nat. Sci. 2010, 2, 1302–1311. [Google Scholar] [CrossRef]
- Pereira, S.; Pinto, A.L.; Cortes, R.; Fontainhas-Fernandes, A.; Coimbra, A.M.; Monteiro, S.M. Gill histopathological and oxidative stress evaluation in native fish captured in Portuguese Northwestern Rivers. Ecotoxicol. Environ. Saf. 2013, 90, 157–166. [Google Scholar] [CrossRef]
- Regoli, F.; Giuliani, M.E.; Benedetti, M.; Arukwe, A. Molecular and biochemical biomarkers in environmental monitoring: A comparison of biotransformation and antioxidant defense systems in multiple tissues. Aquat. Toxicol. 2011, 105, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Regoli, F.; Giuliani, M.E. Oxidative pathways of chemical toxicity and oxidative stress biomarkers in marine organisms. Mar. Environ. Res. 2014, 93, 106–117. [Google Scholar] [CrossRef]
- Meng, S.L.; Liu, T.; Song, C.; Zhang, C.; Qiu, L.P.; Chen, J.Z.; Xu, P. Determination of Methomyl Residue in Water by Ultra Performance Liquid Chromatography-Tandem Mass Spectrometry. J. Anhui Agric. Sci. 2018, 46, 166–167. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Wang, H. Effects and Mechanisms of Monocrotophos Pesticide on Sex Hormone Synthesis and Transformation in the Male Goldfish (Carassius auratus); Ocean University of China: Qingdao, China, 2013. [Google Scholar]
- Thongbuakaew, T.; Suwansa-Ard, S.; Chaiyamoon, A.; Cummins, S.F.; Sobhon, P. Sex steroids and steroidogenesis-related genes in the sea cucumber, holothuria scabra and their potential role in gonad maturation. Sci. Rep. 2021, 11, 2194. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Li, X. Research progress in steroidogenic acute regulatory protein. J. Shanxi Norm. Univ. Nat. Sci. Ed. 2009, 37, 85–89. [Google Scholar]
- Barannikova, I.A.; Bayunova, L.V.; Semenkova, T.B. Serum levels of testosterone, 11-ketotestosterone and oestradiol-17β in three species of sturgeon during gonadal development and final maturation induced by hormonal treatment. J. Fish. Biol. 2004, 64, 1330–1338. [Google Scholar] [CrossRef]
- Mu, X.Y. The Toxicity Effect and Mechanism of Difenoconazole on Zebrafish (Danio rerio); Agricultural University: Beijing, China, 2015. [Google Scholar]
- Hogan, N.S.; Currie, S.; Le Blanc, S.; Hewitt, M.L.; MacLatchy, D.L. Modulation of steroidogenesis and estrogen signalling in the estuarine killifish (Fundulus heteroclitus) exposed to ethinylestradiol. Aquat. Toxicol. 2010, 98, 148–156. [Google Scholar] [CrossRef]
- Meng, X.X.; Wei, Y.L.; Liang, M.Q.; Xu, H.G. Progress in Cholesterol Nutritional Requirements of Fish. Chin. J. Anim. Nutr. 2021, 33, 719–728. [Google Scholar]
- Sharpe, R.L.; Woodhouse, A.; Moon, T.W.; Trudeau, V.L.; MacLacthy, D.L. β-Sitosterol and 17β-estradiol alter gonadal steroidogenic acute regulatory protein (StAR) expression in goldfish, Carassius auratus. Gen. Comp. Edocr. 2007, 151, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Jefcoate, C.R.; McNamara, B.C.; Artemenko, I.; Yamazaki, T. Regulation of cholesterol movement to mitochondrial cytochrome P450scc in steroid hormone synthesis. J. Steroid. Biochem. Mol. Biol. 1992, 43, 751–767. [Google Scholar] [CrossRef]
- Babin, P.J.; Vernier, J.M. Plasma lipoproteins in fish. J. Lipid Res. 1989, 30, 467–489. [Google Scholar] [CrossRef]
- Sayed, A.D.; Mahmoud, U.M.; Mekkawy, I.A. eproductive biomarkers to identify endocrine disruption in Clarias gariepinus exposed to 4-nonylphenol. Ecotoxicol. Environ. Saf. 2012, 78, 310–319. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, S.; Chen, X.; Song, C.; Fan, L.; Qiu, L.; Hu, G.; Chen, J.; Xu, P. Response of Sex Steroid Hormone Synthesis Substrates in Serum and Testes of Male Tilapia (Oreochromis niloticus) Exposed to Methomyl and Its Recovery Pattern. Appl. Sci. 2021, 11, 10997. https://doi.org/10.3390/app112210997
Meng S, Chen X, Song C, Fan L, Qiu L, Hu G, Chen J, Xu P. Response of Sex Steroid Hormone Synthesis Substrates in Serum and Testes of Male Tilapia (Oreochromis niloticus) Exposed to Methomyl and Its Recovery Pattern. Applied Sciences. 2021; 11(22):10997. https://doi.org/10.3390/app112210997
Chicago/Turabian StyleMeng, Shunlong, Xi Chen, Chao Song, Limin Fan, Liping Qiu, Gengdong Hu, Jiazhang Chen, and Pao Xu. 2021. "Response of Sex Steroid Hormone Synthesis Substrates in Serum and Testes of Male Tilapia (Oreochromis niloticus) Exposed to Methomyl and Its Recovery Pattern" Applied Sciences 11, no. 22: 10997. https://doi.org/10.3390/app112210997
APA StyleMeng, S., Chen, X., Song, C., Fan, L., Qiu, L., Hu, G., Chen, J., & Xu, P. (2021). Response of Sex Steroid Hormone Synthesis Substrates in Serum and Testes of Male Tilapia (Oreochromis niloticus) Exposed to Methomyl and Its Recovery Pattern. Applied Sciences, 11(22), 10997. https://doi.org/10.3390/app112210997