
Effect of Coptischinensis, Glycyrrhiza uralensis, and Fermented Glycine max Extract as Proactive Therapy for Atopic Dermatitis


Abstract
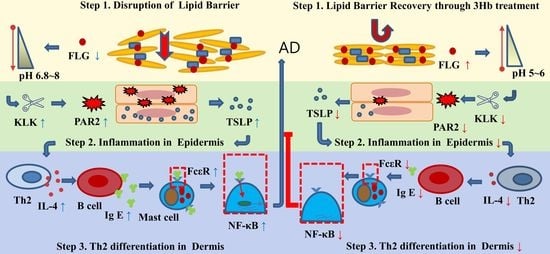
:
1. Introduction
2. Materials and Methods
2.1. Preparation of the 3Hb Extract
2.2. Bioactive Marker Profiling
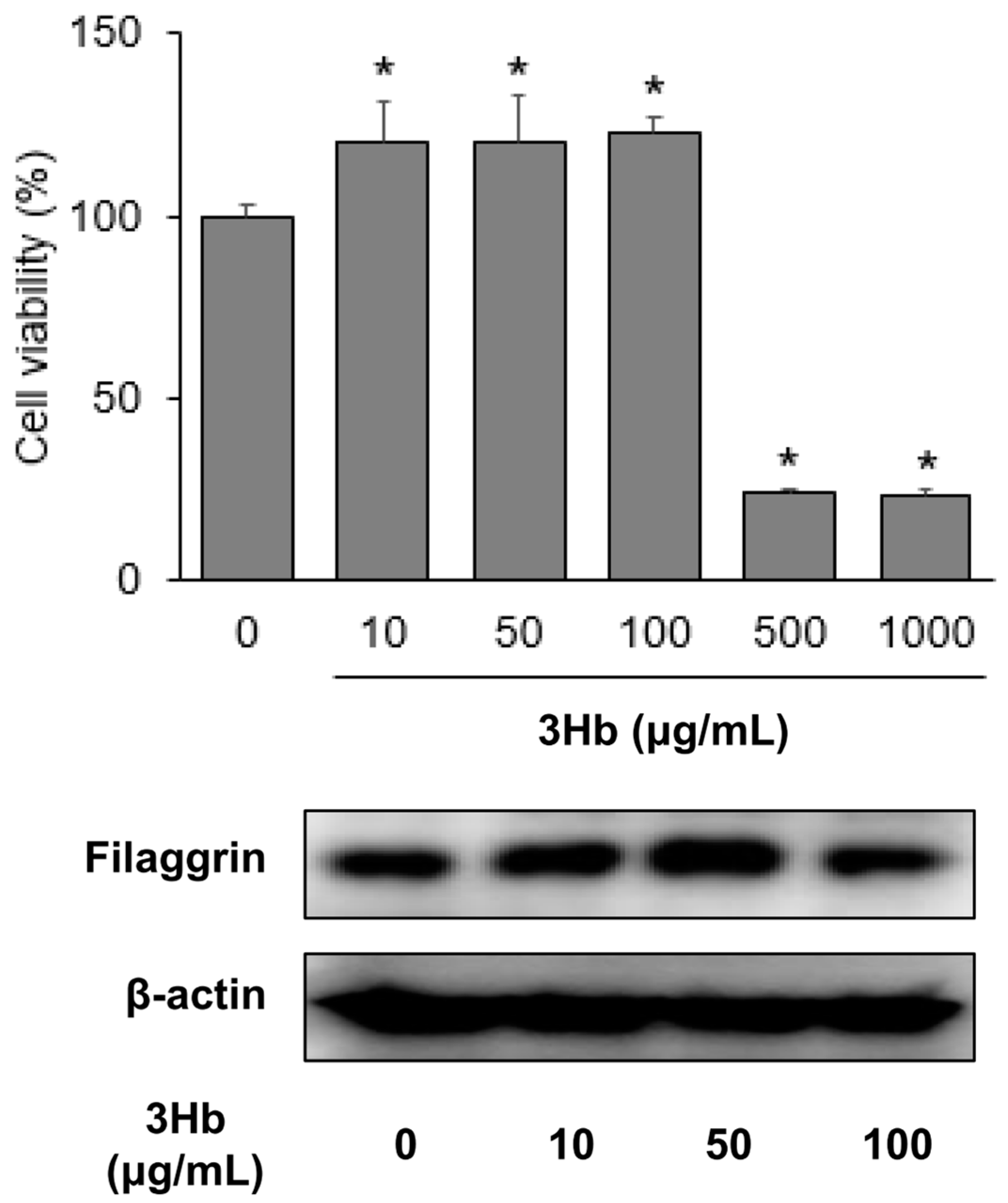
2.3. FLG Generating Capacity
2.3.1. Cell Viability
2.3.2. Western Blot Analysis
2.4. Experimental Animals
2.5. Lipid Barrier Elimination and Administration
2.6. Skin Score, TEWL, and pH Measurement in Skin
2.7. Histochemistry
2.8. Immunohistochemistry
2.9. Image Analysis and Statistical Analysis
3. Results
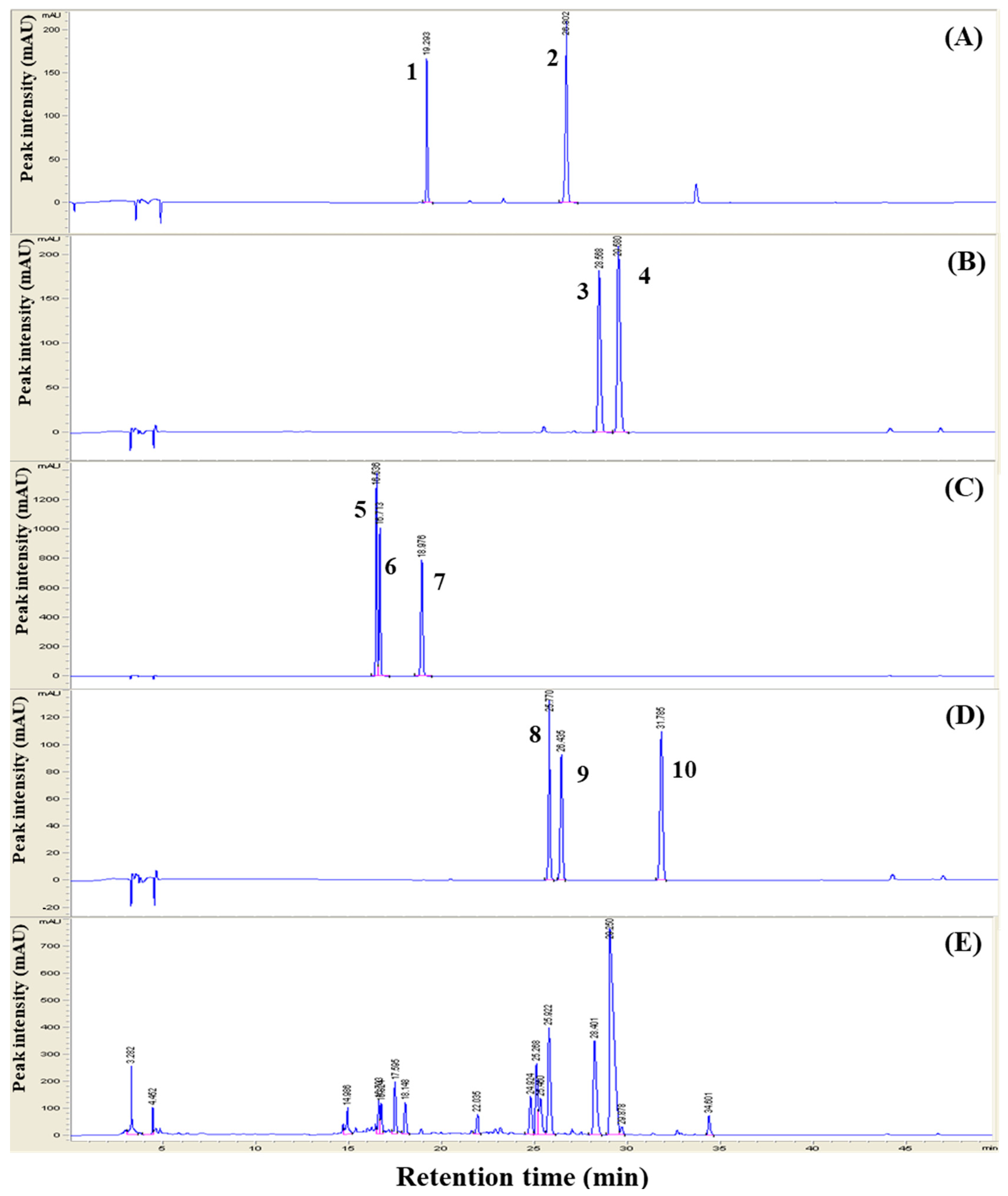
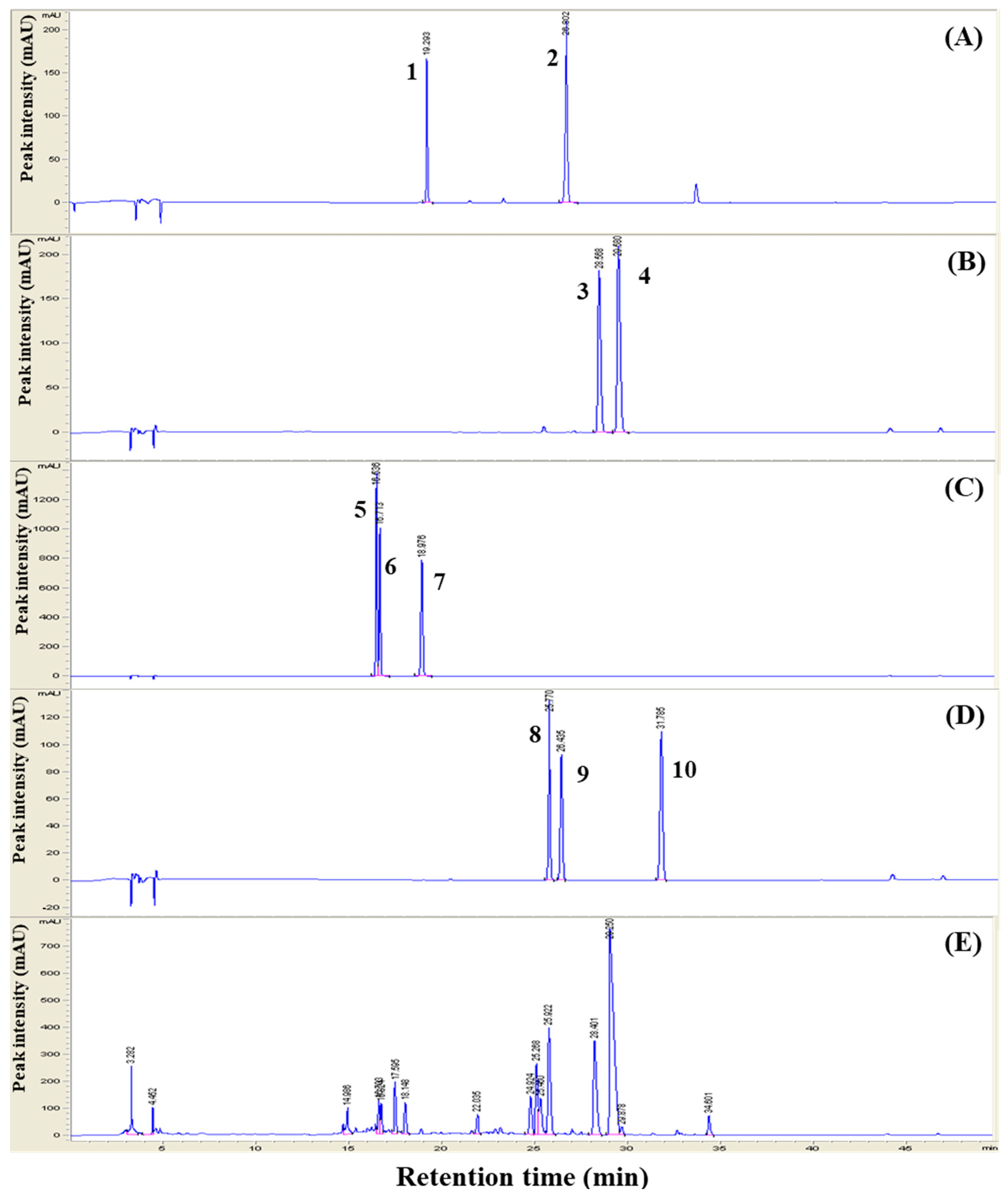
3.1. HPLC Analysis of the Herbal Extracts
3.2. Regulation of Lipid Barrier
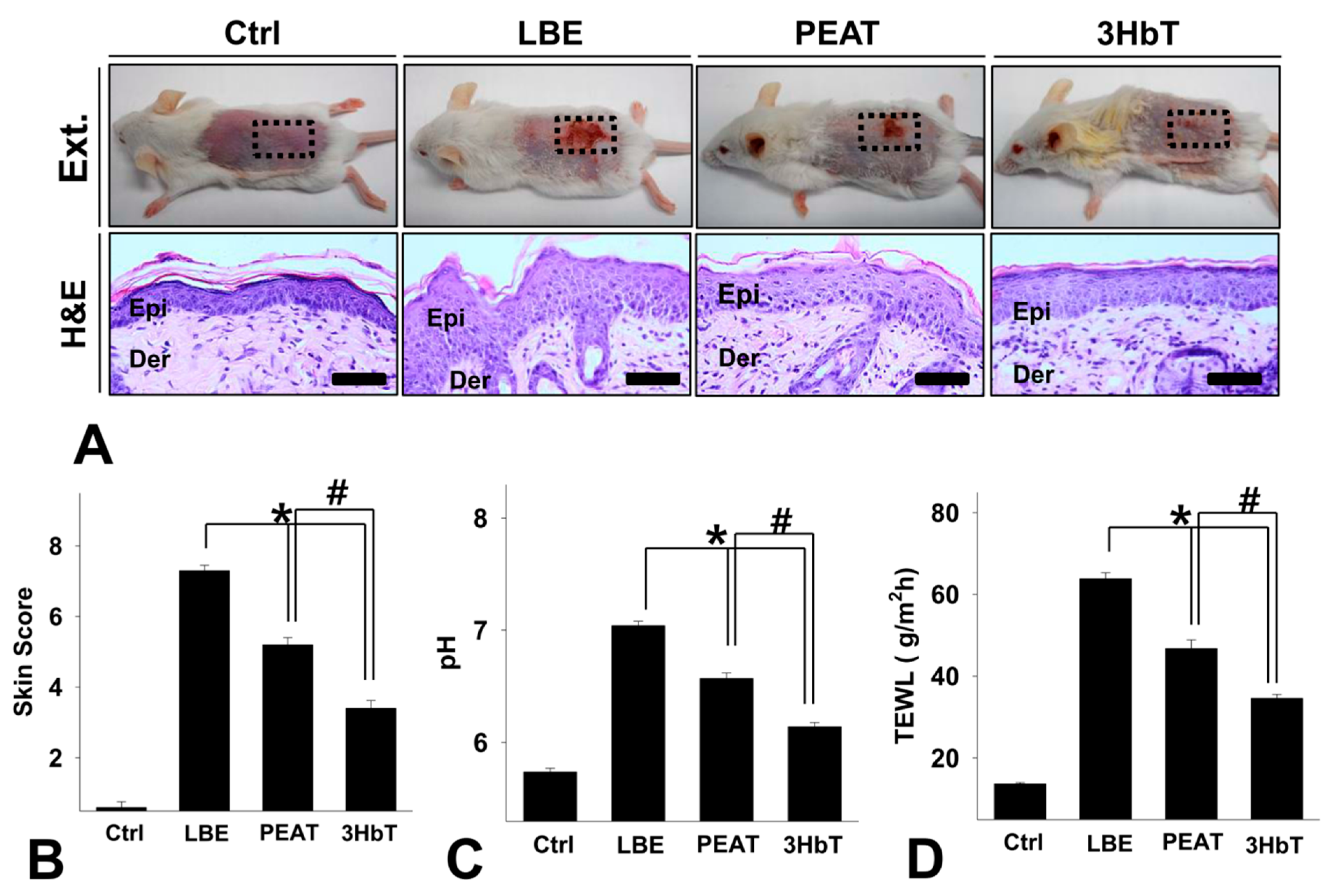
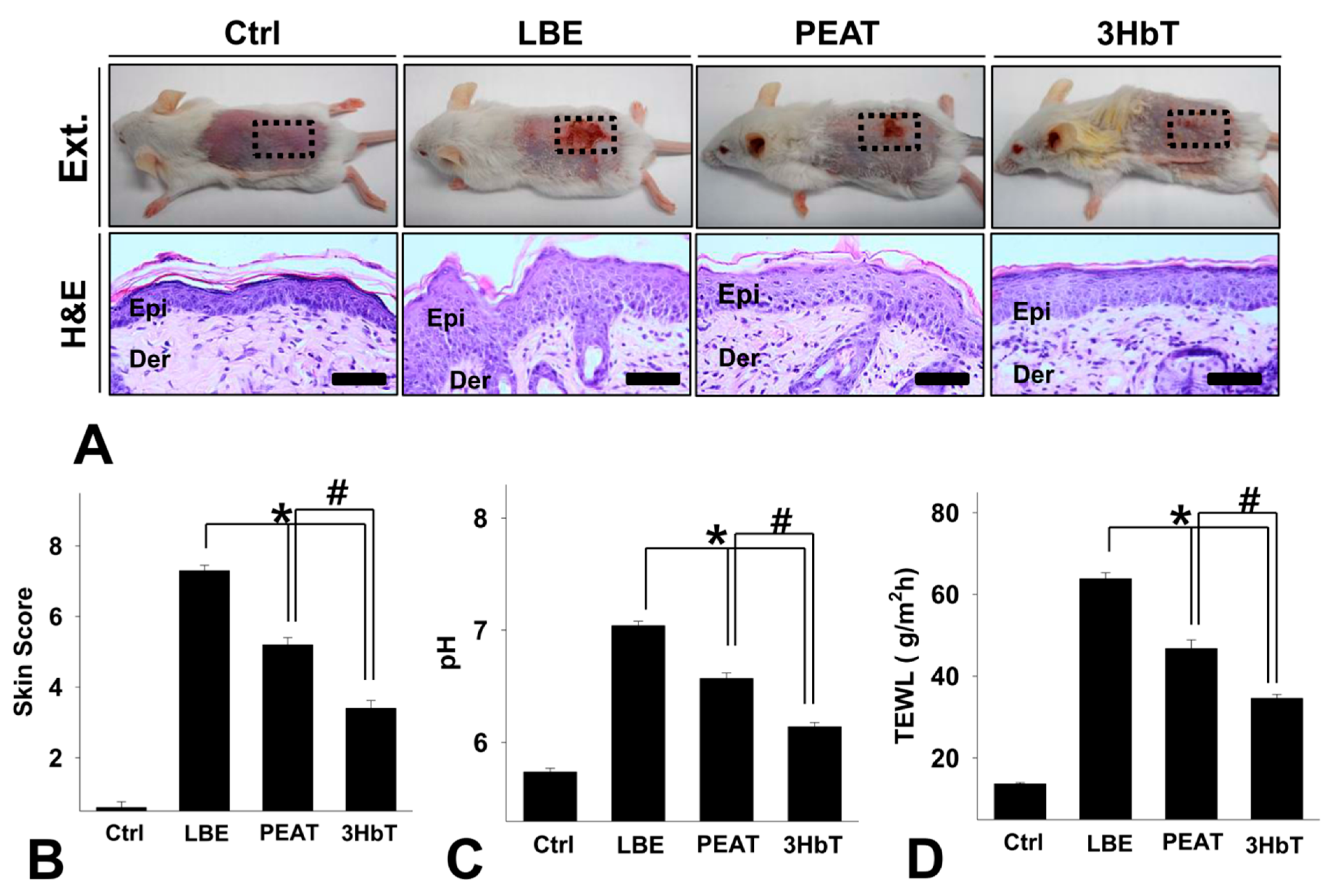
3.3. Changes in General Skin Features
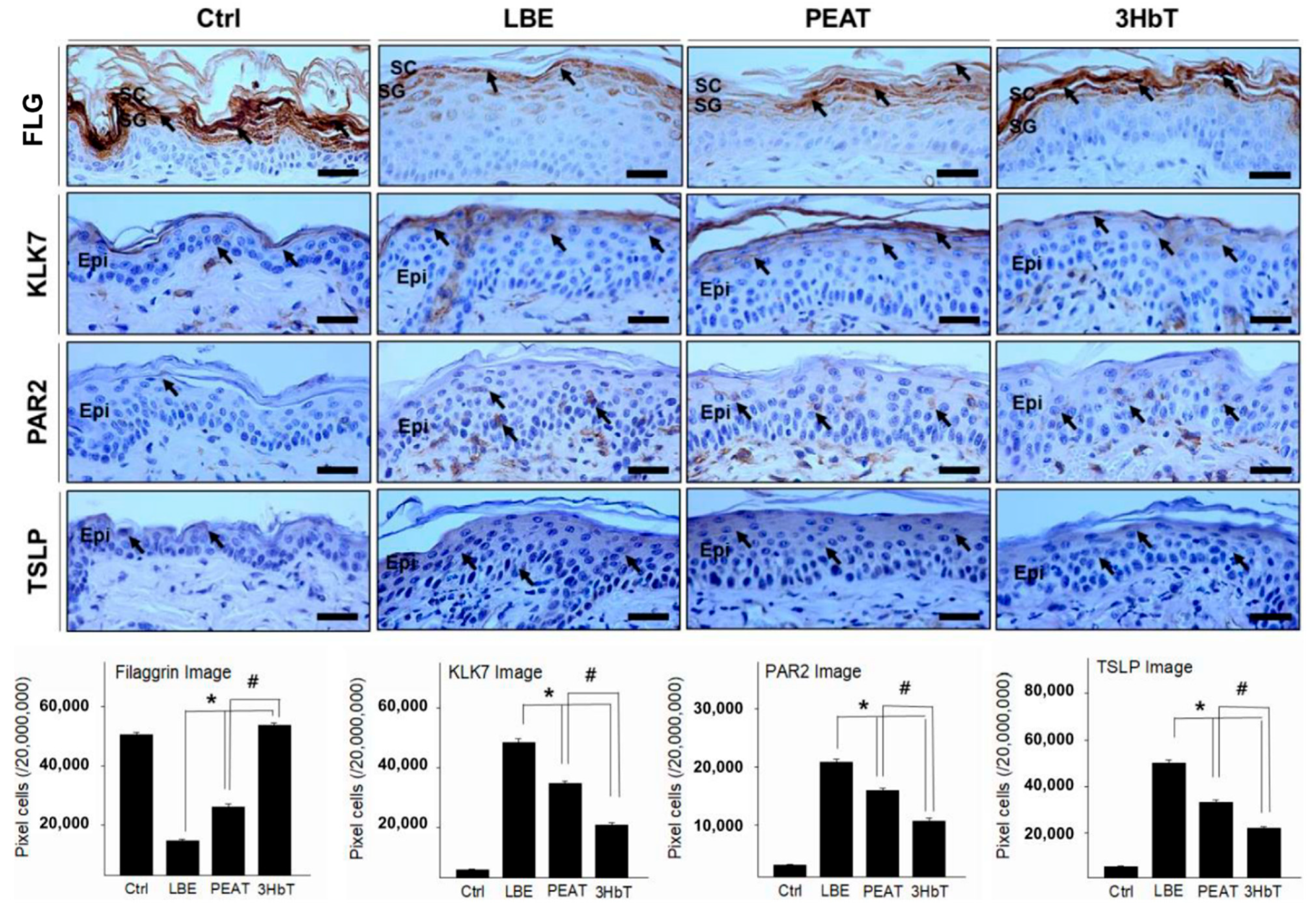
3.4. Effect of Change in FLG-pH-Inflammation
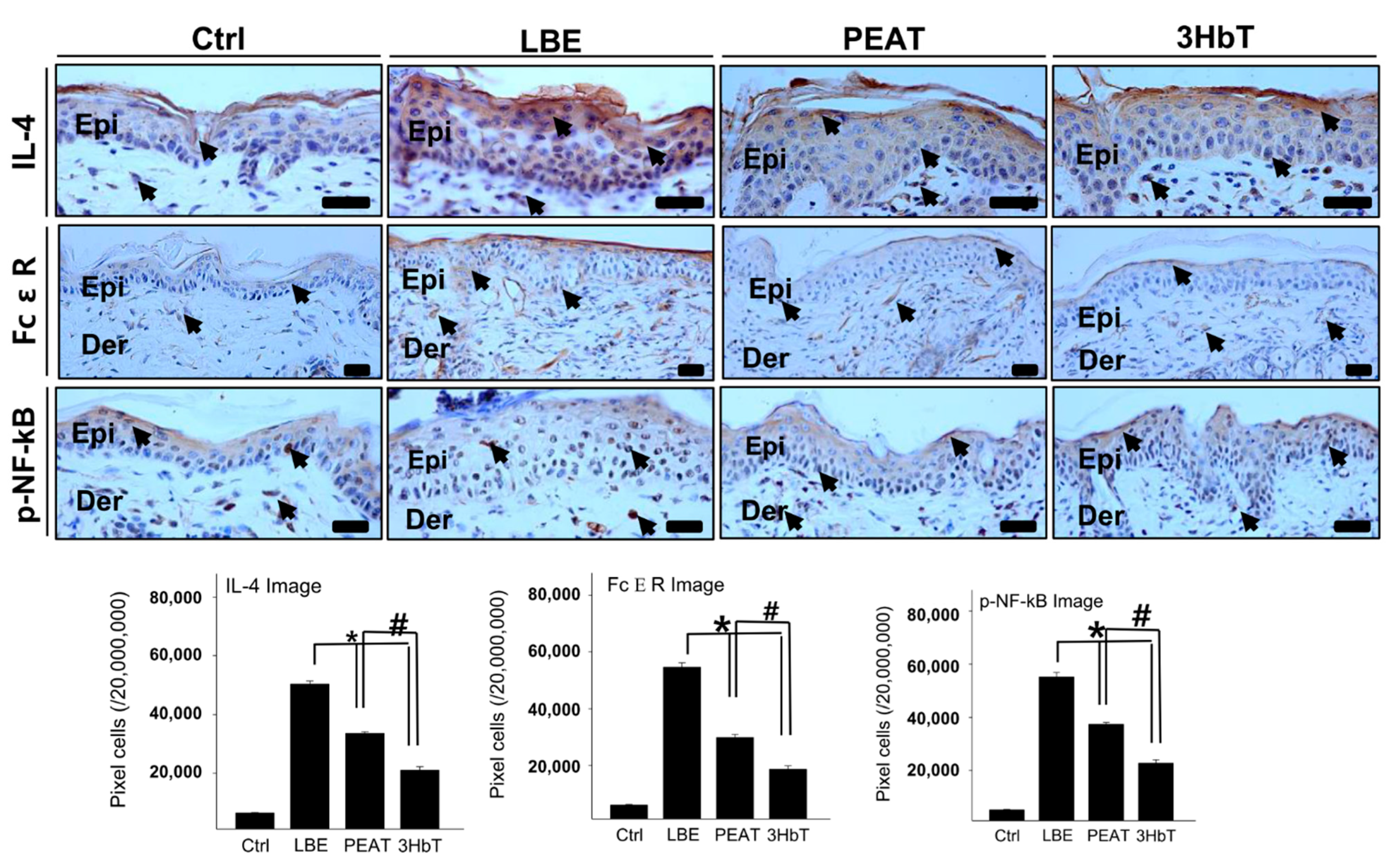
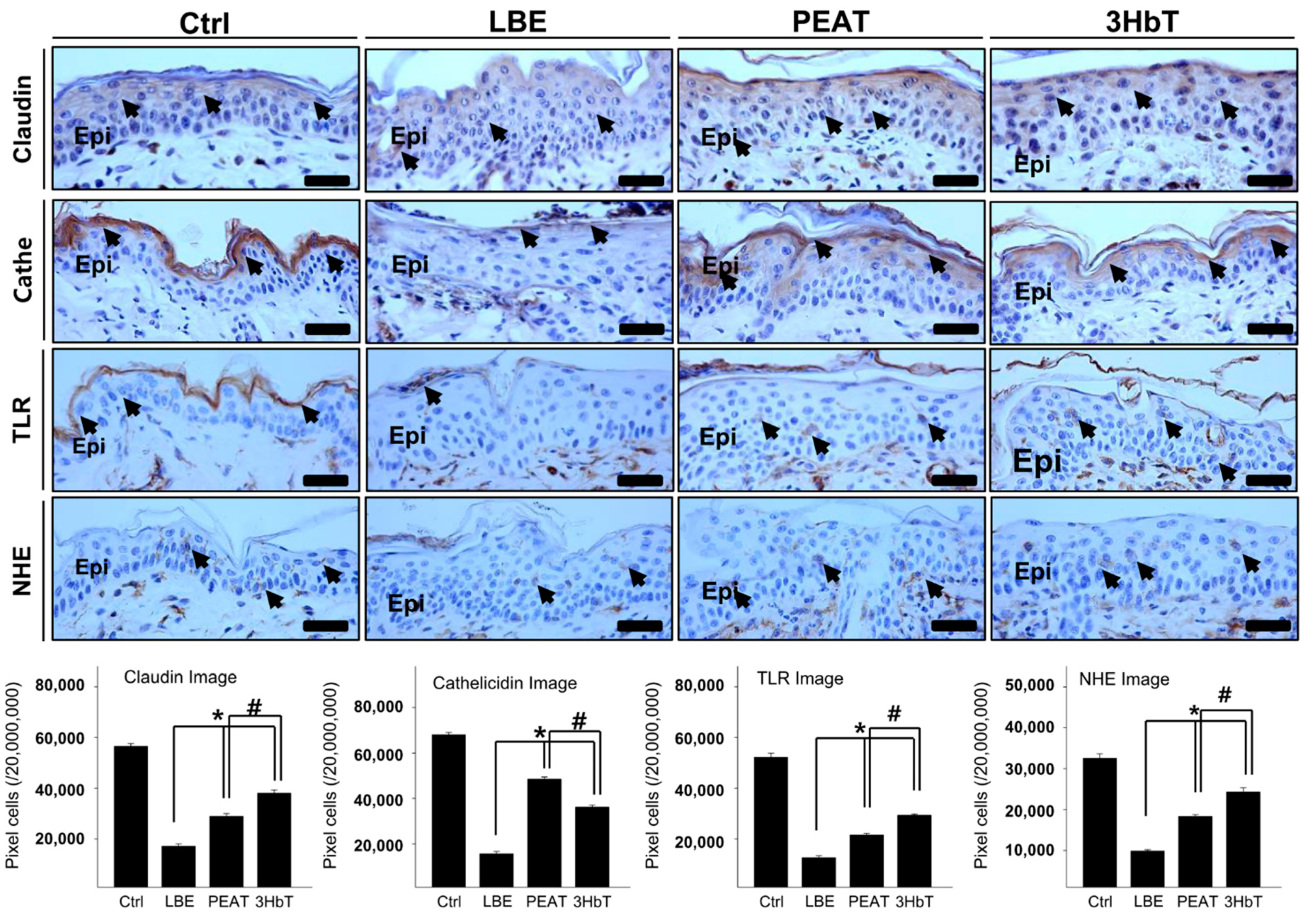
3.5. Regulation of Th2 Differentiation
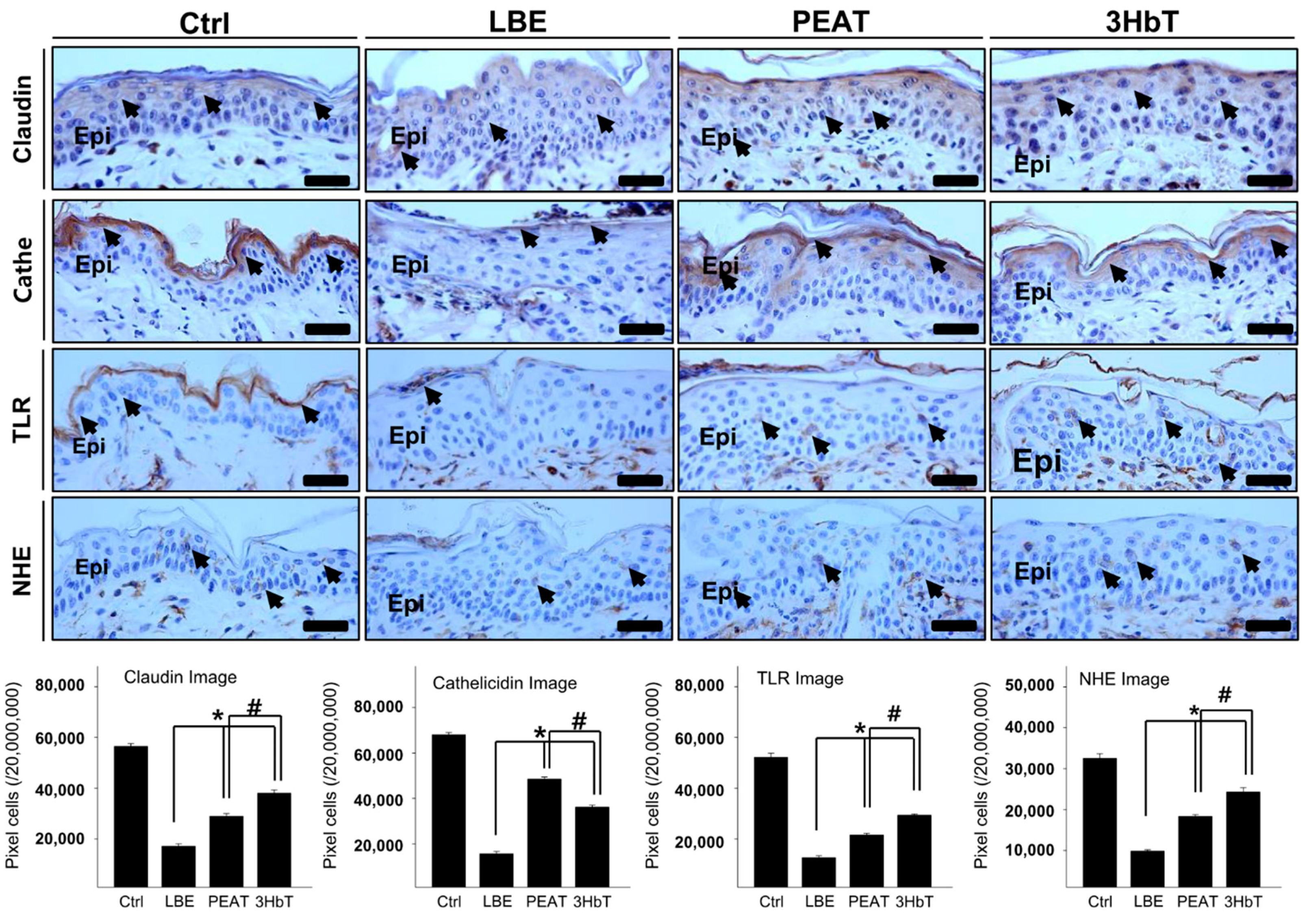
3.6. Regulation of the Skin Microenvironment for Defense
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kim, K.H.; Shim, J.S.; Kim, H.J.; Son, E.D. Penta-O-galloyl-β-D-glucose from Paeonia lactiflora Pall. root extract enhances the expression of skin barrier genes via EGR3. J. Ethnopharmacol. 2020, 10, 112337. [Google Scholar] [CrossRef]
- Matsui, T.; Amagai, M. Dissecting the formation, structure and barrier function of the stratum corneum. Int. Immunol. 2015, 27, 269–280. [Google Scholar] [CrossRef] [Green Version]
- Egawa, G.; Kabashima, K. Barrier dysfunction in the skin allergy. Allergol. Int. 2018, 67, 3–11. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Misery, L.; Proksch, E.; Metz, M.; Ständer, S.; Schmelz, M. Skin Barrier Damage and Itch: Review of Mechanisms, Topical Management and Future Directions. Acta Derm. Venereol. 2019, 99, 1201–1209. [Google Scholar] [CrossRef] [Green Version]
- Takei, K.; Mitoma, C.; Hashimoto-Hachiya, A.; Takahara, M.; Tsuji, G.; Nakahara, T.; Furue, M. Galactomyces fermentation filtrate prevents T helper2-mediated reduction of filaggrin in an aryl hydrocarbon receptor-dependent manner. Clin. Exp. Dermatol. 2015, 40, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Takei, K.; Mitoma, C.; Hashimoto-Hachiya, A.; Uchi, H.; Takahara, M.; Tsuji, G.; Kido-Nakahara, M.; Nakahara, T.; Furue, M. Antioxidant soybean tar Glyteer rescues T-helper-mediated downregulation of filaggrin expression via aryl hydrocarbon receptor. J. Dermatol. 2015, 42, 171–180. [Google Scholar] [CrossRef]
- Tenn, M.W.; Ellis, A.K. The clinical relevance of filaggrin mutations: Effect on allergic disease. Ann. Allergy Asthma Immunol. 2016, 117, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, L.; Lou, G.H.; Zeng, H.R.; Hu, J.; Huang, Q.W.; Peng, W.; Yang, X.B. Coptidis Rhizoma: A comprehensive review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. Pharm. Biol. 2019, 57, 193–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, S.; Li, Z.; Song, W.; Wang, Y.; Liang, W.; Li, K.; Tang, S.; Wang, Q.; Qiao, X.; Zhou, D.; et al. Bioactive Constituents of Glycyrrhiza uralensis (Licorice): Discovery of the Effective Components of a Traditional Herbal Medicine. J. Nat. Prod. 2016, 79, 281–292. [Google Scholar] [CrossRef] [PubMed]
- Jung, A.R.; Ahn, S.H.; Park, I.S.; Park, S.Y.; Jeong, S.I.; Cheon, J.H.; Kim, K.B. Douchi (fermented Glycine max Merr.) alleviates atopic dermatitis-like skin lesions in NC/Nga mice by regulation of PKC and IL-4. BMC Complement. Altern. Med. 2016, 16, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.Y.; Ahn, S.H.; Yang, I.J.; Park, S.Y.; Kim, K.B. Effect of Hataedock Treatment on Epidermal Structure Maintenance through Intervention in the Endocannabinoid System. Evid. Based Complement. Altern. Med. 2020, 2020, 3605153. [Google Scholar] [CrossRef]
- Cha, H.Y.; Ahn, S.H.; Cheon, J.H.; Park, S.Y.; Kim, K.B. Hataedock treatment has preventive therapeutic effects for atopic dermatitis through skin barrier protection in Dermatophagoides farinae-induced NC/Nga mice. J. Ethnopharmacol. 2017, 206, 327–336. [Google Scholar] [CrossRef]
- Man, M.Q.; Hu, L.Z.; Elias, P.M. Herbal Medicines Prevent the Development of Atopic Dermatitis by Multiple Mechanisms. Chin. J. Integr. Med. 2019, 25, 151–160. [Google Scholar] [CrossRef]
- Hussain, Z.; Thu, H.E.; Shuid, A.N.; Kesharwani, P.; Khan, S.; Hussain, F. Phytotherapeutic potential of natural herbal medicines for the treatment of mild-to-severe atopic dermatitis: A review of human clinical studies. Biomed. Pharmacother. 2017, 93, 596–608. [Google Scholar] [CrossRef]
- Wu, S.; Yu, D.; Liu, W.; Zhang, J.; Liu, X.; Wang, J.; Yu, M.; Li, Z.; Chen, Q.; Li, X.; et al. Magnoflorine from Coptis chinese has the potential to treat DNCB-induced Atopic dermatits by inhibiting apoptosis of keratinocyte. Bioorganic Med. Chem. 2020, 28, 115093. [Google Scholar] [CrossRef]
- Kim, K.J.; Xuan, S.H.; Park, S.N. Licoricidin, an isoflavonoid isolated from Glycyrrhiza uralensis Fisher, prevents UVA-induced photoaging of human dermal fibroblasts. Int. J. Cosmet. Sci. 2017, 39, 133–140. [Google Scholar] [CrossRef]
- Zou, K.; Li, Z.; Zhang, Y.; Zhang, H.Y.; Zhu, W.L.; Shi, J.Y.; Jia, Q.; Li, Y.M. Advances in the study of berberine and its derivatives: A focus on anti-inflammatory and anti-tumor effects in the digestive system. Acta Pharmacol. Sin. 2017, 38, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Long, J.; Song, J.; Zhong, L.; Liao, Y.; Liu, L.; Li, X. Palmatine: A review of its pharmacology, toxicity and pharmacokinetics. Biochimie 2019, 162, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.W.; Zhao, R.J.; Park, S.J.; Lee, J.R.; Cho, I.J.; Yang, C.H.; Kim, S.J.; Kim, S.C. Anti-inflammatory effects of liquiritigenin as a consequence of the inhibition of NF-κB-dependent iNOS and proinflammatory cytokines production. Br. J. Pharmacol. 2008, 154, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.Y.; Ha, J.Y.; Kim, K.M.; Jung, Y.S.; Jung, J.C.; Oh, S. Anti-Inflammatory activities of licorice extract and its active compounds, glycyrrhizic acid, liquiritin and liquiritigenin, in BV2 cells and mice liver. Molecules 2015, 20, 13041–13054. [Google Scholar] [CrossRef]
- Zhang, J.H.; Tatsumi, E.; Ding, C.H.; Li, L.T. Angiotensin I-converting enzyme inhibitory peptides in Douchi, a Chinese traditional fermented soybean product. Food Chem. 2006, 98, 551–557. [Google Scholar] [CrossRef]
- Lin, J.; Xu, Y.; Zhao, T.; Sun, L.; Yang, M.; Liu, T.; Sun, H.; Zhang, L. Genistein suppresses smooth muscle cell-derived foam cell formation through tyrosine kinase pathway. Biochem. Biophys. Res. Commun. 2015, 463, 1297–1304. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.S.; Hong, L.; Guan, Y.; Dong, X.W.; Zheng, H.S.; Tan, G.L.; Xie, Q.X. Inhibition of airway inflammation, hyperresponsiveness and remodeling by soy isoflavone in a murine model of allergic asthma. Int. Immunopharmacol. 2011, 11, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Cha, H.Y.; Ahn, S.H.; Cheon, J.H.; Park, I.S.; Kim, J.T.; Kim, K.B. Hataedock Treatment Has Preventive Therapeutic Effects in Atopic Dermatitis-Induced NC/Nga Mice under High-Fat Diet Conditions. Evid. Based Complement. Altern. Med. 2016, 2016, 1739760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Kim, T.H.; Kang, M.S.; Ahn, J.O.; Choi, J.H.; Chung, J.Y. Comparison of the presentation of atopic dermatitis induced by trinitrochlorobenzene and house dust mite in NC/Nga mice. J. Vet. Sci. 2020, 21, 59. [Google Scholar] [CrossRef] [PubMed]
- Hart, P.H.; Norval, M. The Multiple Roles of Urocanic Acid in Health and Disease. J. Investig. Dermatol. 2021, 141, 496–502. [Google Scholar] [CrossRef]
- Brown, M.A.; Hural, J. Functions of IL-4 and Control of Its Expression. Crit. Rev. Immunol. 2017, 37, 181–212. [Google Scholar] [CrossRef]
- Schwartz, C.; Eberle, J.U.; Voehringer, D. Basophils in inflammation. Eur. J. Pharmacol. 2016, 778, 90–95. [Google Scholar] [CrossRef]
- Blank, U.; Charles, N.; Benhamou, M. The high-affinity immunoglobulin E receptor as pharmacological target. Eur. J. Pharmacol. 2016, 778, 24–32. [Google Scholar] [CrossRef]
- Elieh Ali Komi, D.; Wöhrl, S.; Bielory, L. Mast Cell Biology at Molecular Level: A Comprehensive Review. Clin. Rev. Allergy Immunol. 2020, 58, 342–365. [Google Scholar] [CrossRef]
- Fan, H.J.; Zhao, X.S.; Tan, Z.B.; Liu, B.; Xu, H.L.; Wu, Y.T.; Xie, L.P.; Bi, Y.M.; Lai, Y.G.; Liang, H.F.; et al. Effects and mechanism of action of Huang-Lian-Jie-Du-Tang in atopic dermatitis-like skin dysfunction in vivo and in vitro. J. Ethnopharmacol. 2019, 240, 111937. [Google Scholar] [CrossRef]
- Gruber, R.; Börnchen, C.; Rose, K.; Daubmann, A.; Volksdorf, T.; Wladykowski, E.; Vidal-Y-Sy, S.; Peters, E.M.; Danso, M.; Bouwstra, J.A.; et al. Diverse regulation of claudin-1 and claudin-4 in atopic dermatitis. Am. J. Pathol. 2015, 185, 2777–2789. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Gallo, R.L. The Critical and Multifunctional Roles of Antimicrobial Peptides in Dermatology. Dermatol. Clin. 2017, 35, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Liu, W.; Zhang, L.J. The Role of Toll-Like Receptors in Skin Host Defense, Psoriasis, and Atopic Dermatitis. J. Immunol. Res. 2019, 2019, 1824624. [Google Scholar] [CrossRef] [PubMed]
- Jang, H.; Matsuda, A.; Jung, K.; Karasawa, K.; Matsuda, K.; Oida, K.; Ishizaka, S.; Ahn, G.; Amagai, Y.; Moon, C.; et al. Skin pH Is the Master Switch of Kallikrein 5-Mediated Skin Barrier Destruction in a Murine Atopic Dermatitis Model. J. Investig. Dermatol. 2016, 136, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.J.; Rawlings, A.V. The chemistry, function and (patho)physiology of stratum corneum barrier ceramides. Int. J. Cosmet. Sci. 2017, 39, 366–372. [Google Scholar] [CrossRef] [Green Version]
- Lowe, A.J.; Leung, D.Y.M.; Tang, M.L.K.; Su, J.C.; Allen, K.J. The skin as a target for prevention of the atopic march. Ann. Allergy Asthma Immunol. 2018, 120, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Sandilands, A.; Sutherland, C.; Irvine, A.D.; McLean, W.H.I. Filaggrin in the frontline: Role in skin barrier function and disease. J. Cell Sci. 2009, 122, 1285–1294. [Google Scholar] [CrossRef] [Green Version]
- Drislane, C.; Irvine, A.D. The role of filaggrin in atopic dermatitis and allergic disease. Ann. Allergy Asthma Immunol. 2020, 124, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Čepelak, I.; Dodig, S.; Pavić, I. Filaggrin and atopic march. Biochem. Med. (Zagreb) 2019, 29, 020501. [Google Scholar] [CrossRef]
- Seguchi, T.; Cui, C.Y.; Kusuda, S.; Takahashi, M.; Aisu, K.; Tezuka, T. Decreased expression of filaggrin in atopic skin. Arch. Dermatol. Res. 1996, 288, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Kishibe, M. Physiological and pathological roles of kallikrein-related peptidases in the epidermis. J. Dermatol. Sci. 2019, 95, 50–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redhu, D.; Franke, K.; Kumari, V.; Francuzik, W.; Babina, M.; Worm, M. Thymic stromal lymphopoietin production induced by skin irritation results from concomitant activation of protease-activated receptor 2 and interleukin 1 pathways. Br. J. Dermatol. 2020, 182, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Furue, M.; Chiba, T.; Tsuji, G.; Ulzii, D.; Kido-Nakahara, M.; Nakahara, T.; Kadono, T. Atopic dermatitis: Immune deviation, barrier dysfunction, IgE autoreactivity and new therapies. Allergol. Int. 2017, 66, 398–403. [Google Scholar] [CrossRef]
- Proksch, E. pH in nature, humans and skin. J. Dermatol. 2018, 45, 1044–1052. [Google Scholar] [CrossRef]
- Peters, N.; Peters, A.T. Atopic dermatitis. Allergy Asthma Proc. 2019, 40, 433–436. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Information | Chromatographic Results (mg/kg) (4) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Liquiritin | Liquiritigenin | Berberine | Palmatine | Daidzein | Glycitein | Genistein | Daidzin | Glycitin | Geinistin | |
| Glycyrrhiziae (1) | 3.51 | 0.10 | - | - | - | - | - | - | - | - |
| Coptis (2) | - | - | 4.84 | 14.88 | - | - | - | - | - | - |
| Glycine (3) | - | - | - | - | 0.03 | 0.01 | 0.02 | 0.17 | 0.09 | 0.33 |
| 3Hb (1:1:1) | 0.25 | 0.49 | 7.19 | 21.97 | 2.96 | 0.55 | 0.06 | 2.19 | 1.13 | 0.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, B.-C.; Ahn, S.-H.; Yang, I.-J.; Kim, K.-B. Effect of Coptischinensis, Glycyrrhiza uralensis, and Fermented Glycine max Extract as Proactive Therapy for Atopic Dermatitis. Appl. Sci. 2021, 11, 8380. https://doi.org/10.3390/app11188380
Park B-C, Ahn S-H, Yang I-J, Kim K-B. Effect of Coptischinensis, Glycyrrhiza uralensis, and Fermented Glycine max Extract as Proactive Therapy for Atopic Dermatitis. Applied Sciences. 2021; 11(18):8380. https://doi.org/10.3390/app11188380
Chicago/Turabian StylePark, Beom-Chan, Sang-Hyun Ahn, In-Jun Yang, and Ki-Bong Kim. 2021. "Effect of Coptischinensis, Glycyrrhiza uralensis, and Fermented Glycine max Extract as Proactive Therapy for Atopic Dermatitis" Applied Sciences 11, no. 18: 8380. https://doi.org/10.3390/app11188380
APA StylePark, B.-C., Ahn, S.-H., Yang, I.-J., & Kim, K.-B. (2021). Effect of Coptischinensis, Glycyrrhiza uralensis, and Fermented Glycine max Extract as Proactive Therapy for Atopic Dermatitis. Applied Sciences, 11(18), 8380. https://doi.org/10.3390/app11188380