
Immunological and Pathophysiological Outcomes of Helminth Infections and Type 2 Diabetes Comorbidity Studies in Humans and Experimental Animals—A Scoping Review


Abstract
:1. Introduction
2. Materials and Methods
2.1. Identification of the Research Question
2.2. Literature Search Strategy
2.3. Study Selection
2.4. Charting the Data and Summarizing the Results
3. Results
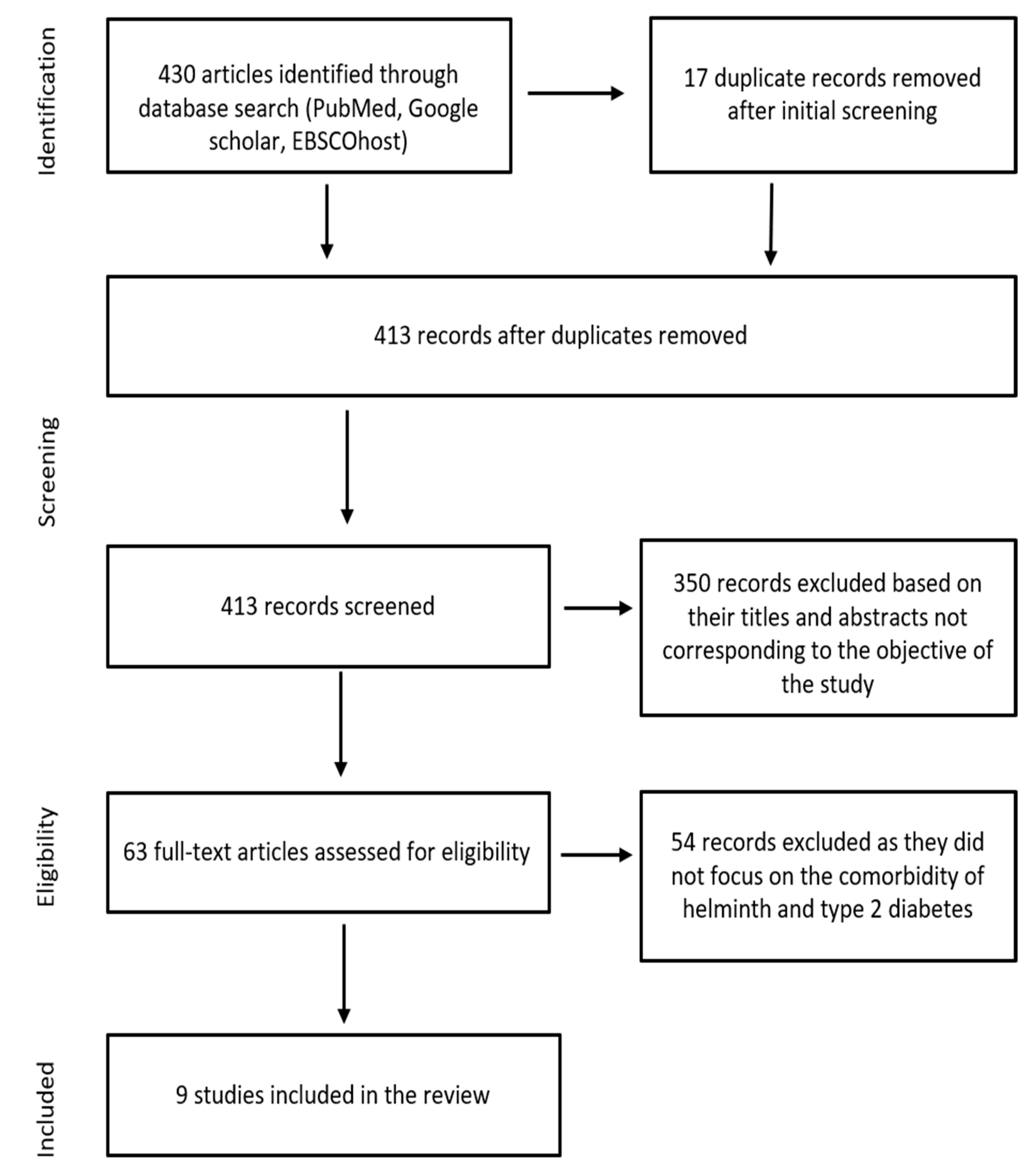
3.1. Eligibility Search Results
3.2. Animal Studies on Helminths Infection and T2DM Comorbidity
3.3. Human Studies on Helminths Infection and T2DM Comorbidity
4. Discussion
4.1. Outcomes of Helminths Infection and T2DM Comorbidity in Experimental Animal Studies
4.2. Outcomes of Helminths Infection and T2DM Comorbidity in Human Studies
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Wu, Y.; Ding, Y.; Tanaka, Y.; Zhang, W. Risk factors contributing to type 2 diabetes and recent advances in the treatment and prevention. Int. J. Med. Sci. 2014, 11, 1185–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajamanickam, A.; Munisankar, S.; Bhootra, Y.; Dolla, C.; Thiruvengadam, K.; Nutman, T.B.; Babu, S. Metabolic consequences of concomitant strongyloides stercoralis infection in patients with type 2 diabetes mellitus. Clin. Infect. Dis. 2018, 69, 697–704. [Google Scholar] [CrossRef] [PubMed]
- Hilal, A.M.; Hamad, H.K. The role of some cytokines in diabetic patients infected with toxoplasmosis. Plant Arch. 2019, 19, 602–605. [Google Scholar]
- Oyebode, O.; Erukainure, O.L.; Sanni, O.; Islam, S. Crassocephalum rubens (Juss. Ex Jacq.) S. Moore improves pancreatic histology, insulin secretion, liver and kidney functions and ameliorates oxidative stress in fructose-streptozotocin induced type 2 diabetic rats. Drug Chem. Toxicol. 2020, 1–10. [Google Scholar] [CrossRef]
- Machado, E.R.; Matos, N.O.; Rezende, S.M.; Carlos, D.; Silva, T.C.; Rodrigues, L.; Almeida, M.J.R.; De Oliveira, M.R.F.; Muniz-Junqueira, M.I.; Gurgel-Gonçalves, R. Host-parasite interactions in individuals with type 1 and 2 diabetes result in higher frequency of ascaris lumbricoides and giardia lamblia in type 2 diabetic individuals. J. Diabetes Res. 2018, 2018, 4238435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Htun, N.S.N.; Odermatt, P.; Paboriboune, P.; Sayasone, S.; Vongsakid, M.; Phimolsarn-Nusith, V.; Tran, X.D.; Ounnavong, P.-S.; Andriama-Hefasoa, N.; Senvanpan, N.-D.; et al. Association between helminth infections and diabetes mellitus in adults from the Lao People’s Democratic Republic: A cross-sectional study. Infect. Dis. Poverty 2018, 7, 105. [Google Scholar] [CrossRef]
- Aravindhan, V.; Mohan, V.; Surendar, J.; Rao, M.M.; Pavankumar, N.; Deepa, M.; Rajagopalan, R.; Kumaraswami, V.; Nutman, T.B.; Babu, S. Decreased prevalence of lymphatic filariasis among diabetic subjects associated with a diminished pro-inflammatory cytokine response (CURES 83). PLoS Negl. Trop. Dis. 2010, 4, e707. [Google Scholar] [CrossRef] [Green Version]
- Hotez, P.J.; Alvarado, M.; Basáñez, M.-G.; Bolliger, I.; Bourne, R.; Boussinesq, M.; Brooker, S.J.; Brown, A.S.; Buckle, G.; Budke, C.M.; et al. The Global Burden of Disease Study 2010: Interpretation and implications for the neglected tropical diseases. PLoS Negl. Trop. Dis. 2014, 8, e2865. [Google Scholar] [CrossRef] [Green Version]
- Shen, S.-W.; Lu, Y.; Li, F.; Shen, Z.-H.; Xu, M.; Yao, W.-F.; Feng, Y.-B.; Yun, J.-T.; Wang, Y.-P.; Ling, W.; et al. The potential long-term effect of previous schistosome infection reduces the risk of metabolic syndrome among Chinese men. Parasite Immunol. 2015, 37, 333–339. [Google Scholar] [CrossRef]
- Su, C.W.; Chen, C.-Y.; Li, Y.; Long, S.R.; Massey, W.; Kumar, D.V.; Walker, W.A.; Shi, H.N. Helminth infection protects against high fat diet-induced obesity via induction of alternatively activated macrophages. Sci. Rep. 2018, 8, 4607. [Google Scholar] [CrossRef]
- Berbudi, A.; Surendar, J.; Ajendra, J.; Gondorf, F.; Schmidt, D.; Neumann, A.-L.; Wardani, A.P.; Layland, L.E.; Hoffmann, L.S.; Pfeifer, A.; et al. Filarial infection or antigen administration improves glucose tolerance in diet-induced obese mice. J. Innate Immun. 2016, 8, 601–616. [Google Scholar] [CrossRef] [PubMed]
- Maizels, R.M.; McSorley, H.J. Regulation of the host immune system by helminth parasites. J. Allergy Clin. Immunol. 2016, 138, 666–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olefsky, J.M.; Glass, C.K. Macrophages, inflammation, and insulin resistance. Annu. Rev. Physiol. 2010, 72, 219–246. [Google Scholar] [CrossRef]
- Cruz, N.G.; Sousa, L.; Sousa, M.O.; Pietrani, N.T.; Fernandes, A.P.; Gomes, K.B. The linkage between inflammation and type 2 diabetes mellitus. Diabetes Res. Clin. Pr. 2013, 99, 85–92. [Google Scholar] [CrossRef]
- Hübner, M.P.; Shi, Y.; Torrero, M.N.; Mueller, E.; Larson, D.; Soloviova, K.; Gondorf, F.; Hoerauf, A.; Killoran, K.E.; Stocker, J.T.; et al. Helminth protection against autoimmune diabetes in nonobese diabetic mice is independent of a type 2 immune shift and requires TGF-β. J. Immunol. 2011, 188, 559–568. [Google Scholar] [CrossRef]
- Taira, K.; Yazawa, R.; Watanabe, A.; Ishikawa, Y.; Okamoto, M.; Takahashi, A.; Asai, F. Syphacia muris infection delays the onset of hyperglycemia in WBN/Kob-Leprfa rats, a new type 2 diabetes mellitus model. Helminthologia 2015, 52, 58–62. [Google Scholar] [CrossRef] [Green Version]
- Daniele, G.; Mendoza, R.G.; Winnier, D.; Fiorentino, T.V.; Pengou, Z.; Cornell, J.; Andreozzi, F.; Jenkinson, C.; Cersosimo, E.; Federici, M.; et al. The inflammatory status score including IL-6, TNF-α, osteopontin, fractalkine, MCP-1 and adiponectin underlies whole-body insulin resistance and hyperglycemia in type 2 diabetes mellitus. Acta Diabetol. 2013, 51, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Arksey, H.; O’Malley, L. Scoping studies: Towards a methodological framework. Int. J. Soc. Res. Methodol. 2005, 8, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; The PRISMA Group. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. Ann. Intern. Med. 2009, 89, 873–880. [Google Scholar]
- Thabet, H.S.; Saleh, N.K.M.; Thabet, S.S.; Abdel-Aziz, M.; Kalleny, N.K. Decreased basal non-insulin-stimulated glucose uptake by diaphragm in streptozotocin-induced diabetic mice infected with Schistosoma mansoni. Parasitol. Res. 2008, 103, 595–601. [Google Scholar] [CrossRef]
- Morimoto, M.; Azuma, N.; Kadowaki, H.; Abe, T.; Suto, Y. Regulation of type 2 diabetes by helminth-induced Th2 immune response. J. Veter. Med. Sci. 2016, 78, 1855–1864. [Google Scholar] [CrossRef] [Green Version]
- Hays, R.; Esterman, A.; Giacomin, P.; Loukas, A.; McDermott, R. Does strongyloides stercoralis infection protect against type 2 diabetes in humans? Evidence from Australian Aboriginal adults. Diabetes Res. Clin. Pr. 2015, 107, 355–361. [Google Scholar] [CrossRef]
- Hays, R.; Giacomin, P.; Olma, L.; Esterman, A.; McDermott, R. The relationship between treatment for Strongyloides stercoralis infection and type 2 diabetes mellitus in an Australian Aboriginal population: A three-year cohort study. Diabetes Res. Clin. Pr. 2017, 134, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Hays, R.; Esterman, A.; McDermott, R. Type 2 diabetes mellitus is associated with strongyloides stercoralis treatment failure in Australian Aboriginals. PLoS Negl. Trop. Dis. 2015, 9, e0003976. [Google Scholar] [CrossRef]
- Rajamanickam, A.; Munisankar, S.; Dolla, C.; Menon, P.A.; Thiruvengadam, K.; Nutman, T.B.; Babu, S. Helminth infection modulates systemic pro-inflammatory cytokines and chemokines implicated in type 2 diabetes mellitus pathogenesis. PLoS Negl. Trop. Dis. 2020, 14, e0008101. [Google Scholar] [CrossRef] [Green Version]
- Donath, M.Y. Inflammation as a sensor of metabolic stress in obesity and type 2 diabetes. Endocrinology 2011, 152, 4005–4006. [Google Scholar] [CrossRef]
- Gol, S.; Pena, R.N.; Rothschild, M.F.; Tor, M.; Estany, J. A polymorphism in the fatty acid desaturase-2 gene is associated with the arachidonic acid metabolism in pigs. Sci. Rep. 2018, 8, 14336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lu, J.; Huang, Y.; Wang, T.; Xu, Y.; Xu, M.; Li, M.; Wang, W.; Li, D.; Bi, Y.; et al. Association of previous schistosome infection with diabetes and metabolic syndrome: A cross-sectional study in rural China. J. Clin. Endocrinol. Metab. 2013, 98, E283–E287. [Google Scholar] [CrossRef] [PubMed]
- Berbudi, A.; Ajendra, J.; Wardani, A.P.; Hoerauf, A.; Hübner, M. Parasitic helminths and their beneficial impact on type 1 and type 2 diabetes. Metab. Res. Rev. 2015, 32, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Tracey, E.F.; McDermott, R.A.; McDonald, M.I. Do worms protect against the metabolic syndrome? A systematic review and meta-analysis. Diabetes Res. Clin. Pr. 2016, 120, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Cani, P.D. Gut microbiota and pregnancy, a matter of inner life. Br. J. Nutr. 2008, 101, 1579–1580. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S. Alternative activation of macrophages. Nat. Rev. Immunol. 2003, 3, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Prins, J.; Venkatesh, B. Clinical review: Adiponectin biology and its role in inflammation and critical illness. Crit. Care 2011, 15, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Van Exel, E.; Gussekloo, J.; De Craen, A.J.; Frölich, M.; Der Wiel, A.B.-V.; Westendorp, R.G. Low production capacity of Interleukin-10 associates with the metabolic syndrome and type 2 diabetes: The Leiden 85-Plus study. Diabetes 2002, 51, 1088–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szebeni, A.; Schloot, N.; Kecskemeti, V.; Hosszúfalusi, N.; Pánczél, P.; Prohaszka, Z.; Fust, G.; Uray, K.; Hudecz, F.; Meierhoff, G. Th1 and Th2 cell responses of type 1 diabetes patients and healthy controls to human heat-shock protein 60 peptides AA437-460 and AA394-408. Inflamm. Res. 2005, 54, 415–419. [Google Scholar] [CrossRef]
- Odegaard, J.I.; Chawla, A. Leukocyte set points in metabolic disease. F1000 Biol. Rep. 2012, 4, 13. [Google Scholar] [CrossRef]
- Daniłowicz-Luebert, E.; O’Regan, N.L.; Steinfelder, S.; Hartmann, S. Modulation of specific and allergy-related immune responses by Helminths. J. Biomed. Biotechnol. 2011, 2011, 821578. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Quintela, A.; Alende, R.; Gude, F.; Campos, J.; Rey, J.; Meijide, L.M.; Vidal, C. Serum levels of immunoglobulins (IgG, IgA, IgM) in a general adult population and their relationship with alcohol consumption, smoking and common metabolic abnormalities. Clin. Exp. Immunol. 2008, 151, 42–50. [Google Scholar] [CrossRef]


{kind=link}
| Reference | Study Objectives | Country | Animals (Number) | Animals (Model) | Helminth Species | Major Findings |
|---|---|---|---|---|---|---|
| [20] | To investigate the effect of schistosomiasis infection on glucose uptake by the diaphragm. | Egypt | 55 | Female Swiss albino mice | Schistosoma mansoni | i. Diabetic mice infected with S. mansoni showed a depressed glucose uptake by the diaphragm when compared to the control diabetic mice. ii. Decreased glycogen content in the skeletal muscle isolated from the diabetic mice was observed as compared to non-infected group. iii. Conclusion made was that chronic S. mansoni infection provides protection against streptozotocin induced T-cell mediated pancreatitis. |
| [21] | To investigate whether nematode infection can modulate T2DM pathology through cytokine regulation in a T2DM mouse model. | Japan | Not described (ND) | Male KK-Ay/TaJcl Mice | Heligmosomoides polygyrus | i. Parasite induced Th2 immune responses prevented type 2 diabetes in KK-Ay/TaJcl mice. ii. Eosinophils were mobilized in the submucosa of H. polygyrus-infected diabetic mice. iii. fat accumulation in the liver was observed in the H. polygyrus infected diabetic mice. iv. No difference in sodium glucose transporter (SGLT1) gene expression was observed in H. polygyrus infected diabetic mice. |
| [16] | To determine whether experimental infection with Syphacia muris delays the onset of hyperglycemia in fa/fa rats. | Japan | 12 | Male fa/fa rats | Syphacia muris | i. Significant decrease in blood glucose level was observed in the infected group when compared to that of control group. ii. S. muris infection also showed a delay in the onset of hyperglycemia in fa/fa rats. iii. S. muris infection did not have effect on the body weight, water and food intake or the organs of the rats that were examined. iv. Immuno-histochemical and histopathological examination of the pancreas showed traces of inflammation in both the S. muris-infected and the non-infected fa/fa rats. |
| Reference | Study Objectives | Country | Population (N) | Gender | Helminths | Major Findings |
|---|---|---|---|---|---|---|
| [7] | To assess the baseline prevalence and the correlation of sero-positivity of W. bancrofti among diabetic subjects. | India | 1463 | Male/ Female | Wuchereria bancrofti | i. Decrease in prevalence of W. bancrofti was seen in subjects with type 2 diabetes, the decrease was associated with lower antigen load and anti-W. bancrofti IgG antibodies. ii. Low prevalence of W. bancrofti was also due to W. bancrofti -mediated mortality since W. bancrofti is known to be a chronic non-lethal disease. iii. W. bancrofti comorbidity with T2DM showed a decreased level of TNF-α and IL-6 cytokines which was already associated with insulin resistance. iv. In the comorbidity group of both W. bancrofti and T2DM, there was a reduction in the pro-inflammatory cytokines TNF-α and IL-6 when compared to the group without W. bancrofti suggesting the influence W. bancrofti has on the development of insulin resistance (IR). |
| [22] | To explore the relationship between infection with Strongyloides stercoralis and the likelihood of having type 2 diabetic mellitus. | Australia | 259 | Male/ Female | Strongyloides stercoralis | i. According to the result of this study, helminths infections resulted in an improved metabolic profile through immune-modulation process of helminth infections. ii. Significant increase in eosinophil count was observed among patients with T2DM when compared to those without. iii. Chronic S. stercoralis infection may, over time, protect against the development of T2DM. iv. Comorbidity of T2DM and S. stercoralis protects against the development of T2DM in humans. |
| [2] | To access the relationship between a soil transmitted helminth Strongyloides stercoralis and T2DM. | India | 118 | Male/ Female | Strongyloides stercoralis | i. S. stercoralis was associated with decrease in insulin and glucagon level which was reversed after treatment with anthelmintic. ii. There was no significant difference in resistin leptin and visfatin between the groups. iii. S. stercoralis provides a degree of protection against T2DM by modulating adipocytokines. |
| [23] | To determine the effect of treatment for S stercoralis on type 2 diabetes mellitus in an Australian Aboriginal population. | Australia | 259 | Male/ Female | Strongyloides stercoralis | i. Treatment of S. stercoralis infection led to the reduction of intensity of inflammatory reaction which resulted in an improved glycemic control. ii. More pronounced weighted immune reaction with increased levels of Th2 inflammatory cytokines were observed in the diabetic group. iii. Persisting positivity to S. stercoralis may have effect in the reduction of cytokine production. |
| [24] | To explore the efficacy of ivermectin in the treatment of serological diagnosed cases of S. stercoralis infection in an Aboriginal community and to describe factors that may influence the outcome of treatment. | Australia | 259 | Male/ Female | Strongyloides stercoralis | i. The study showed that ivermectin is an effective treatment for S. stercoralis and that pre-existing T2DM might be a risk factor for treatment failure. ii. T2DM patients with relative infections of S. stercoralis have higher numbers of auto-infective larvae with subsequent re-establishment of a patent infection. |
| [25] | To examine the association of cytokines and chemokines in helminth-diabetes comorbidity. | India | 118 | Male/ Female | Strongyloides stercoralis | i. Co-existent chronic S. stercoralis infection is associated with a dampened inflammatory cytokine and chemokine response in T2DM. ii. The study demonstrated the depression of circulating levels of cytokines and chemokines in the S. stercoralis and T2DM comorbidity group. iii. A degree of protection was provided by S. stercoralis from the pathology associated with T2DM by modulating the levels of cytokines and chemokines milieu. |
| References | Country | Animal/ Human Host | Total Sample Size (N) | Type of Diabetes/Helminths | Samples Collected for Determination of Infection | Diagnostic Techniques Used | Outcome of T2DM and Helminth(s) Comorbidity Group | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diabetes | Helminths | Diabetes | Helminths | Helminths | Diabetes | Protection | No Change | No Protection | ||||
| [20] | Egypt | Female Swiss albino mice | 55 | T2DM | Schistosoma mansoni | Blood | Liver/ Intestine | Portal perfusion | Glucose oxidase | + | - | - |
| [21] | Japan | Male KK-Ay/TaJcl mice | Not described (ND) | T2DM | Heligmosomoides polygyrus | Blood | Small intestine/Liver | PCR | Fuji Drichem System | + | - | - |
| [16] | Japan | Male fa/fa rats | 12 | T2DM | Syphacia muris | Blood | Eggs collected from the perianal region | Cellophane tape method | Glucose oxidase | + | - | - |
| [7] | India | Humans (male/ female) | 1463 | T2DM | Wuchereria bancrofti | Blood | Serum | W. bancrofti Og4C3 antigen capture | Glucose oxidase | + | - | - |
| [22] | Australia | Humans (male/ female) | 259 | T2DM | Strongyloides stercoralis | Blood | Not described (ND) | S. stercoralis ELISA test | Glucose oxidase | + | - | - |
| [2] | India | Humans (male/ female) | 118 | T2DM | Strongyloides stercoralis | Blood | Stool sample | Stool microscopy | Glucose oxidase | + | - | - |
| [23] | Australia | Humans (male/ female) | 259 | T2DM | Strongyloides stercoralis | Blood | Faeces | Multivalent PCR test | Glucose oxidase | + | - | - |
| [24] | Australia | Humans (male/ female) | 259 | T2DM | Strongyloides stercoralis | Blood | Not described (ND) | S. stercoralis ELISA test | Glucose oxidase | + | - | - |
| [25] | India | Humans (male/ female) | 118 | T2DM | Strongyloides stercoralis | Blood | Stool | Kato-Katz | Glucose oxidase | + | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silas, E.; Ndlovu, S.; Tshilwane, S.I.; Mukaratirwa, S. Immunological and Pathophysiological Outcomes of Helminth Infections and Type 2 Diabetes Comorbidity Studies in Humans and Experimental Animals—A Scoping Review. Appl. Sci. 2021, 11, 8079. https://doi.org/10.3390/app11178079
Silas E, Ndlovu S, Tshilwane SI, Mukaratirwa S. Immunological and Pathophysiological Outcomes of Helminth Infections and Type 2 Diabetes Comorbidity Studies in Humans and Experimental Animals—A Scoping Review. Applied Sciences. 2021; 11(17):8079. https://doi.org/10.3390/app11178079
Chicago/Turabian StyleSilas, Ekuyikeno, Siyanda Ndlovu, Selaelo Ivy Tshilwane, and Samson Mukaratirwa. 2021. "Immunological and Pathophysiological Outcomes of Helminth Infections and Type 2 Diabetes Comorbidity Studies in Humans and Experimental Animals—A Scoping Review" Applied Sciences 11, no. 17: 8079. https://doi.org/10.3390/app11178079
APA StyleSilas, E., Ndlovu, S., Tshilwane, S. I., & Mukaratirwa, S. (2021). Immunological and Pathophysiological Outcomes of Helminth Infections and Type 2 Diabetes Comorbidity Studies in Humans and Experimental Animals—A Scoping Review. Applied Sciences, 11(17), 8079. https://doi.org/10.3390/app11178079