
Characterization of Two Hydrogen Peroxide Resistant Peroxidases from Rhodococcus opacus 1CP

 ,
, 
(This article belongs to the Section Applied Biosciences and Bioengineering)
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Culture Conditions
2.2. Cloning and Expression
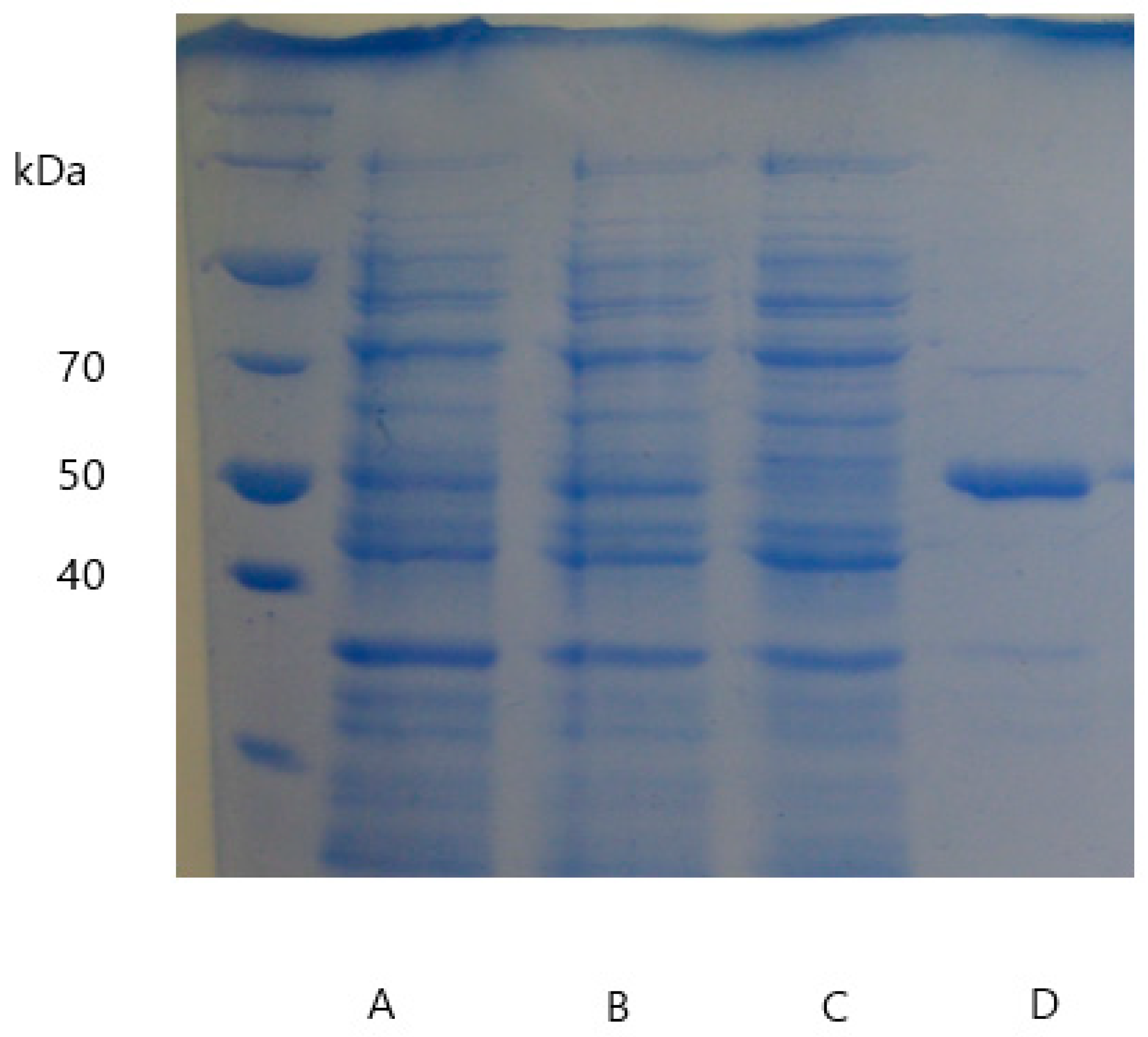
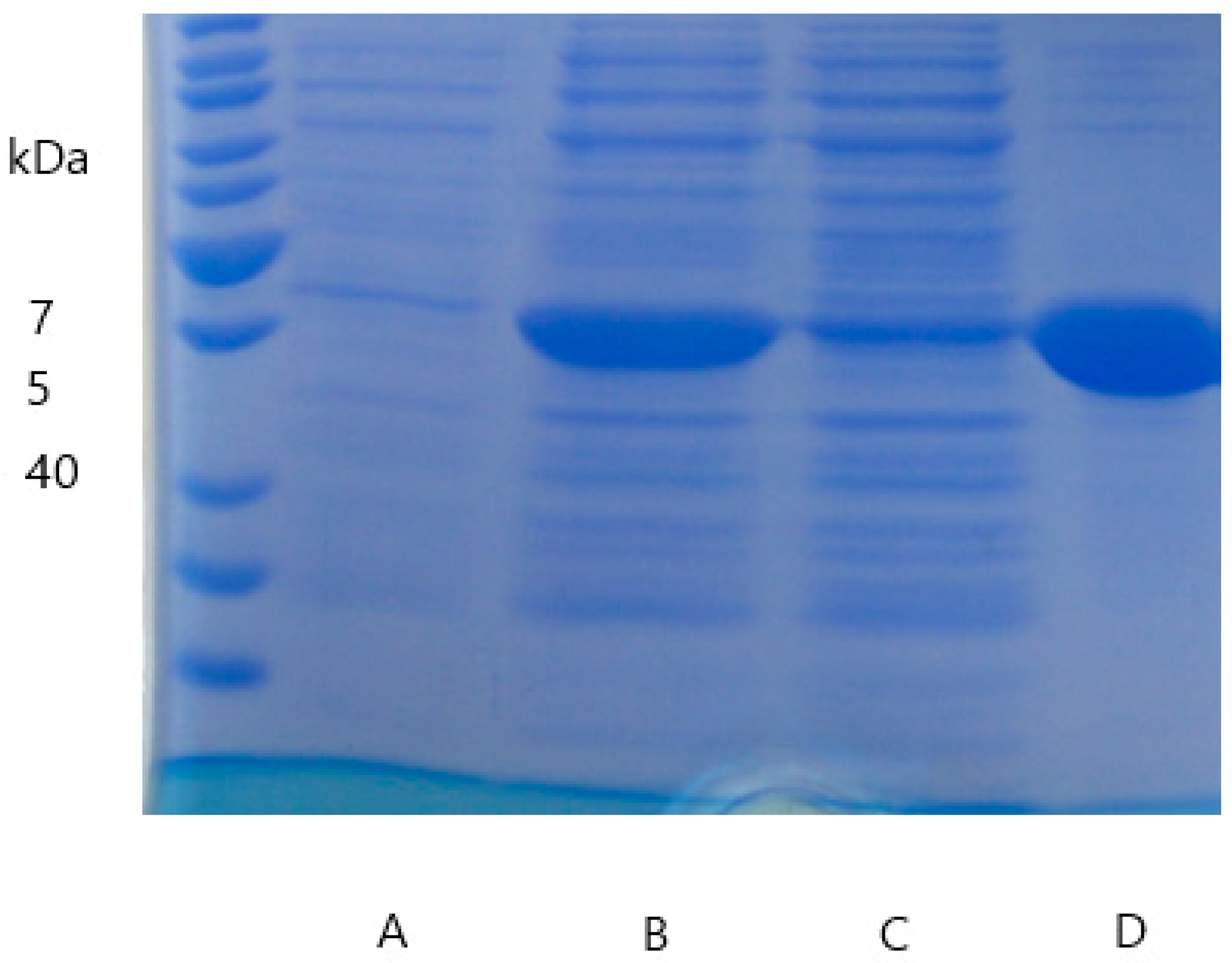
2.3. Enzyme Purification, Quantification, and Verification
2.4. Enzyme Characterization
2.5. Dye-Decolorization Capability
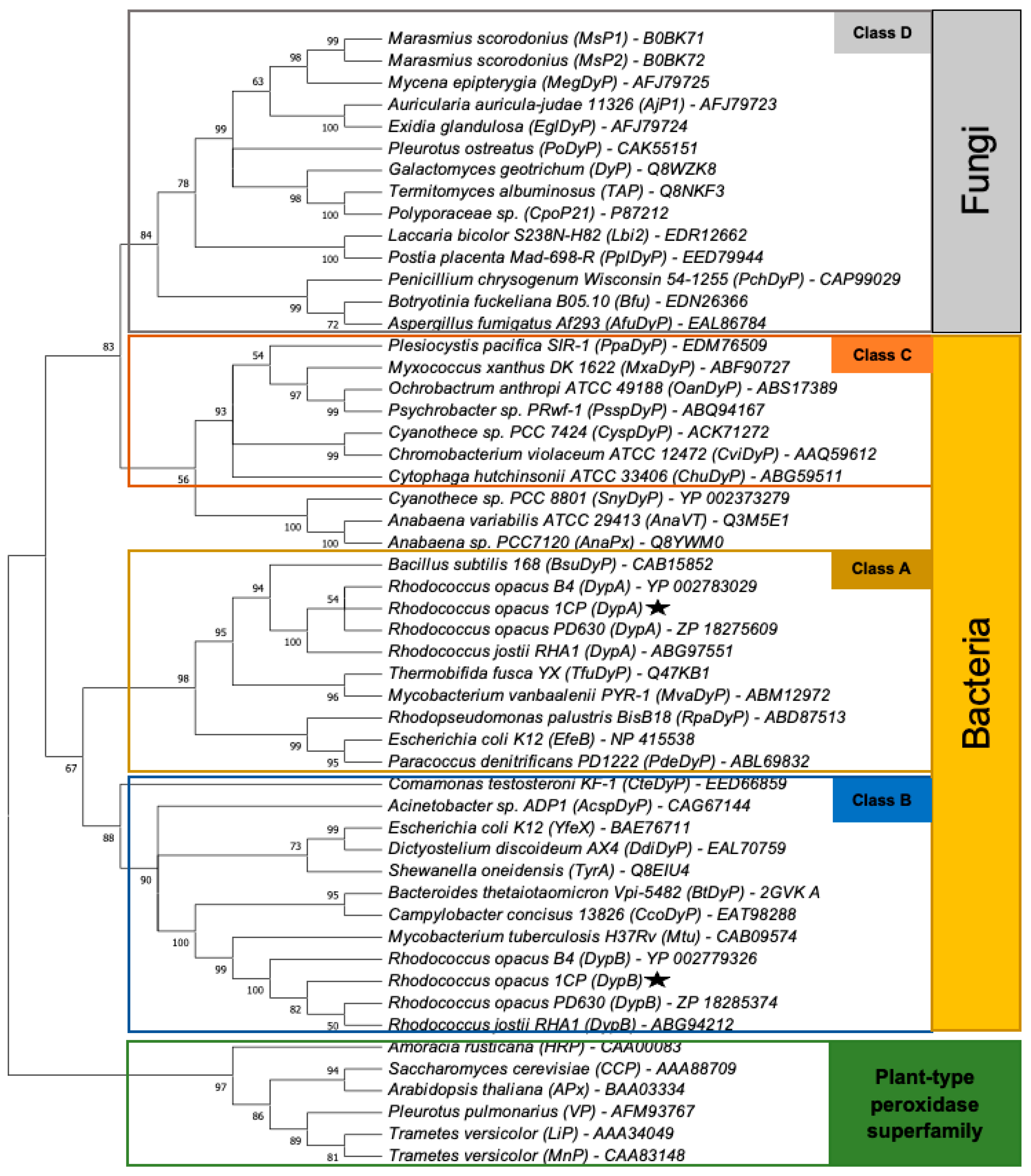
2.6. Sequence Analysis
3. Results
3.1. Identification of Dyp Genes
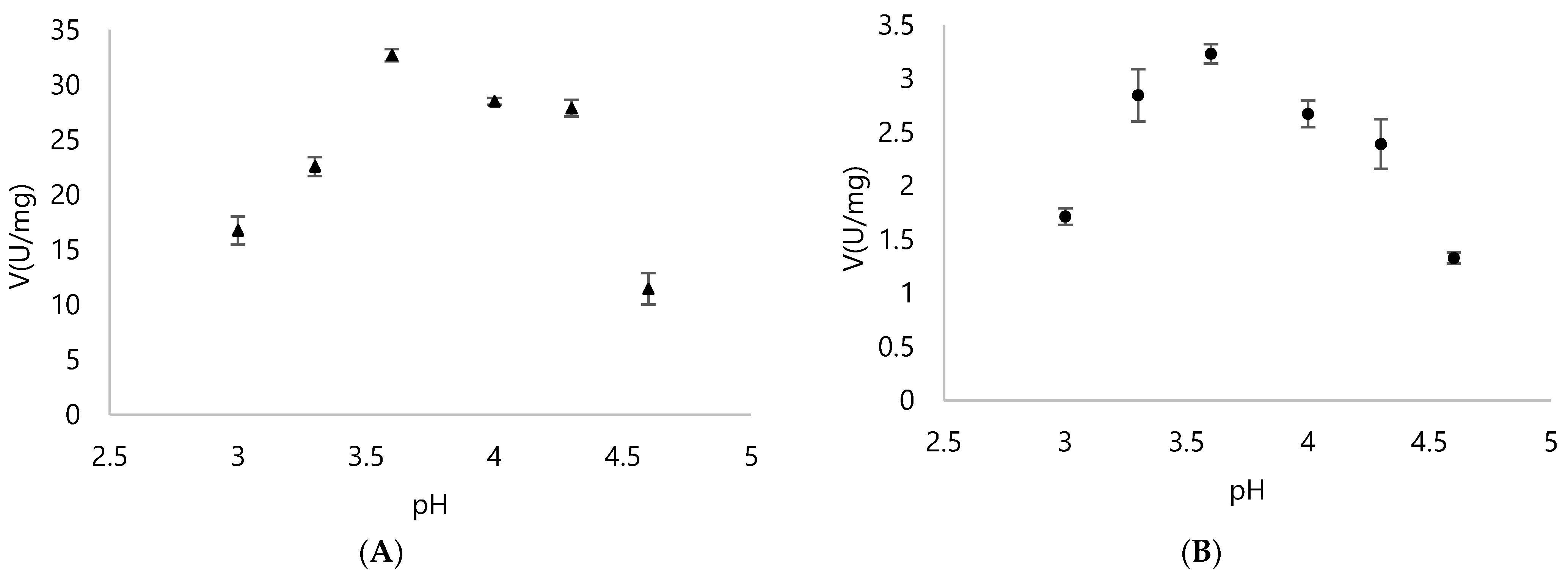
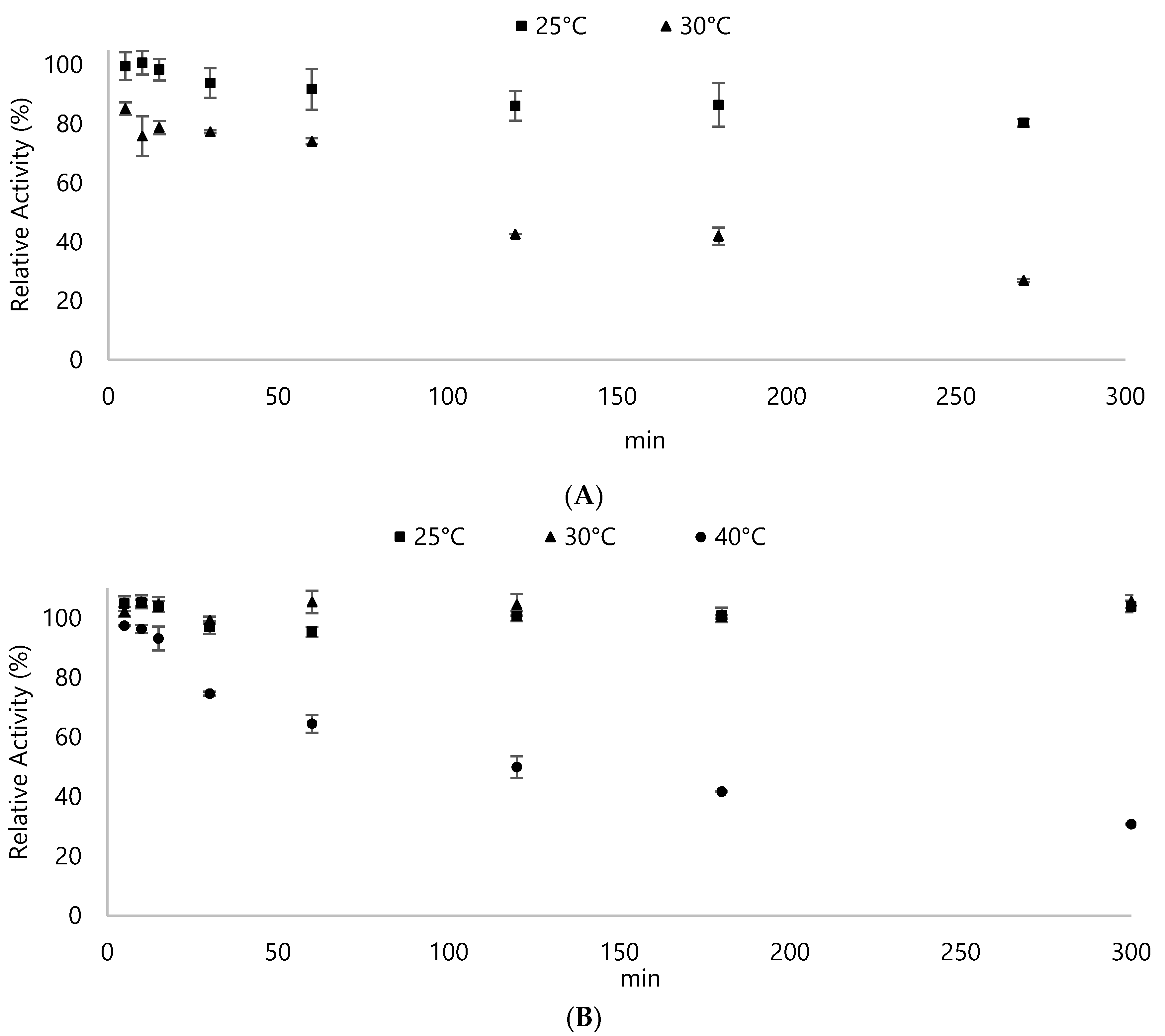
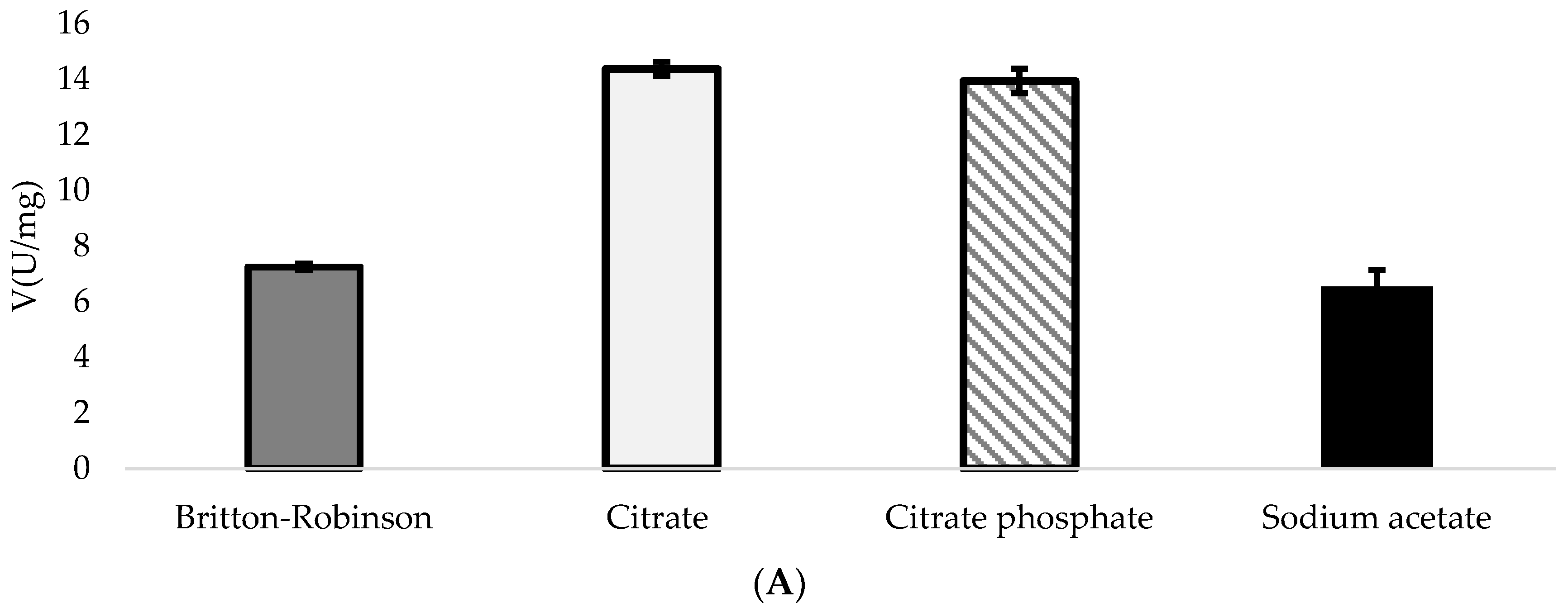
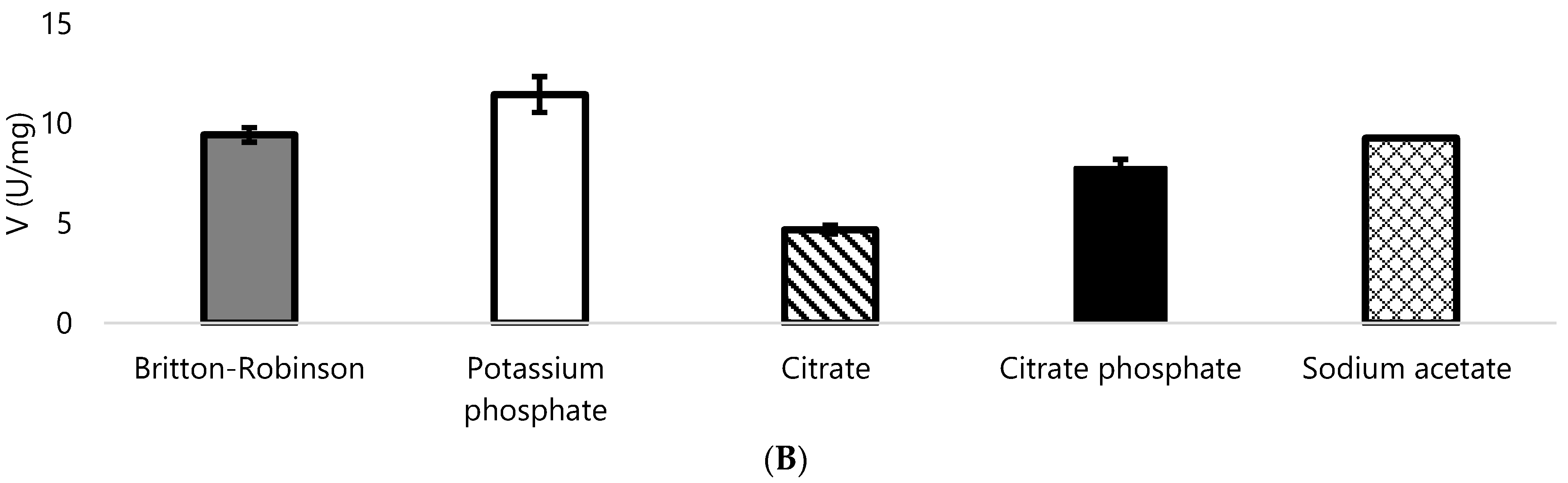
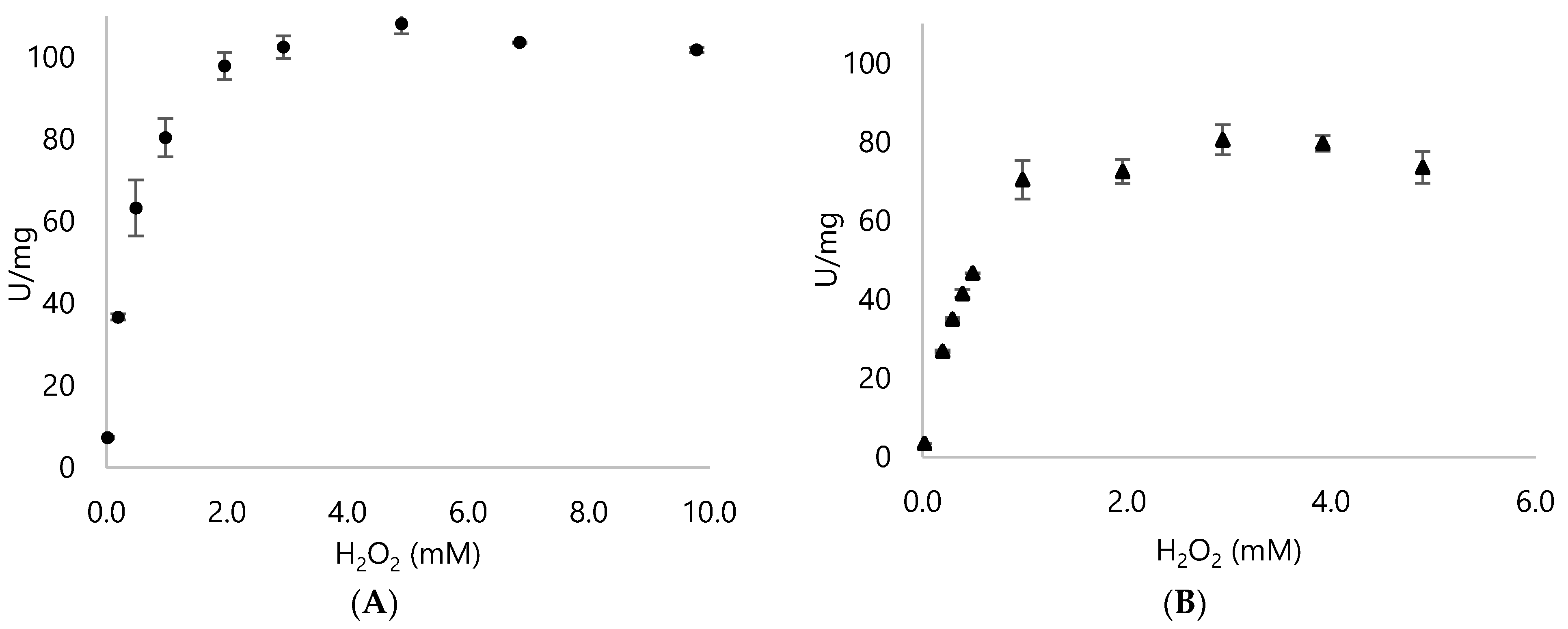
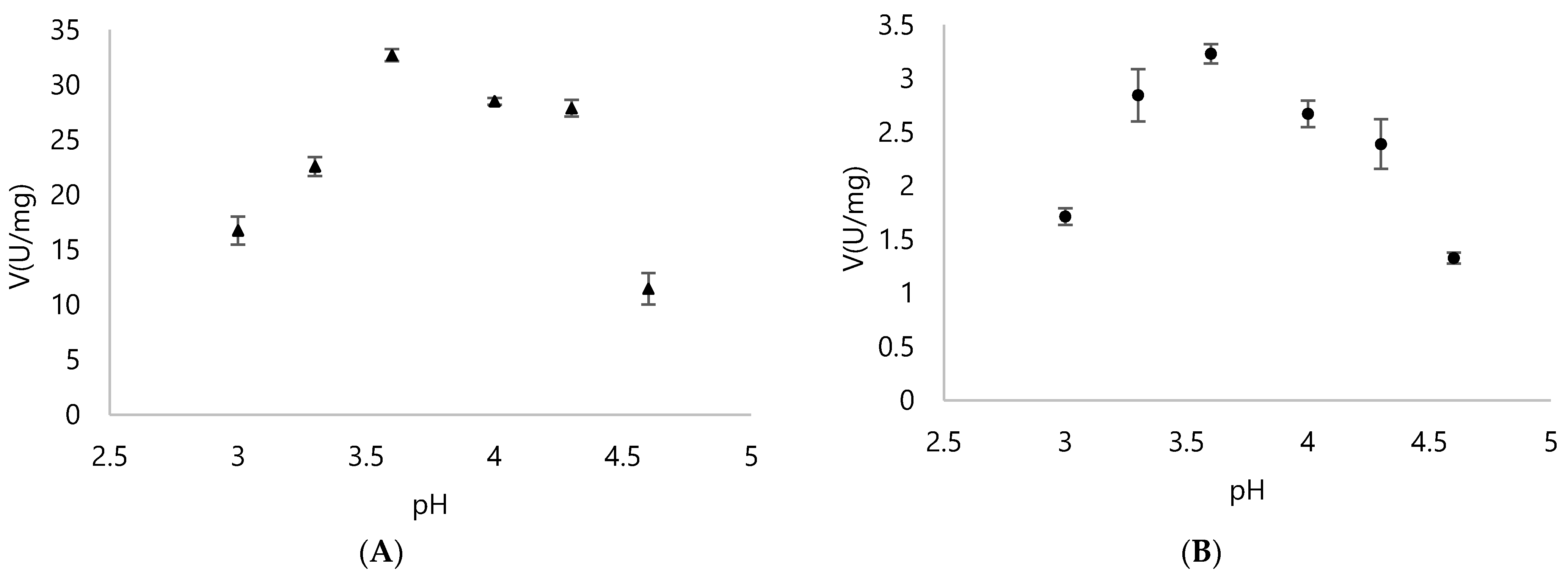
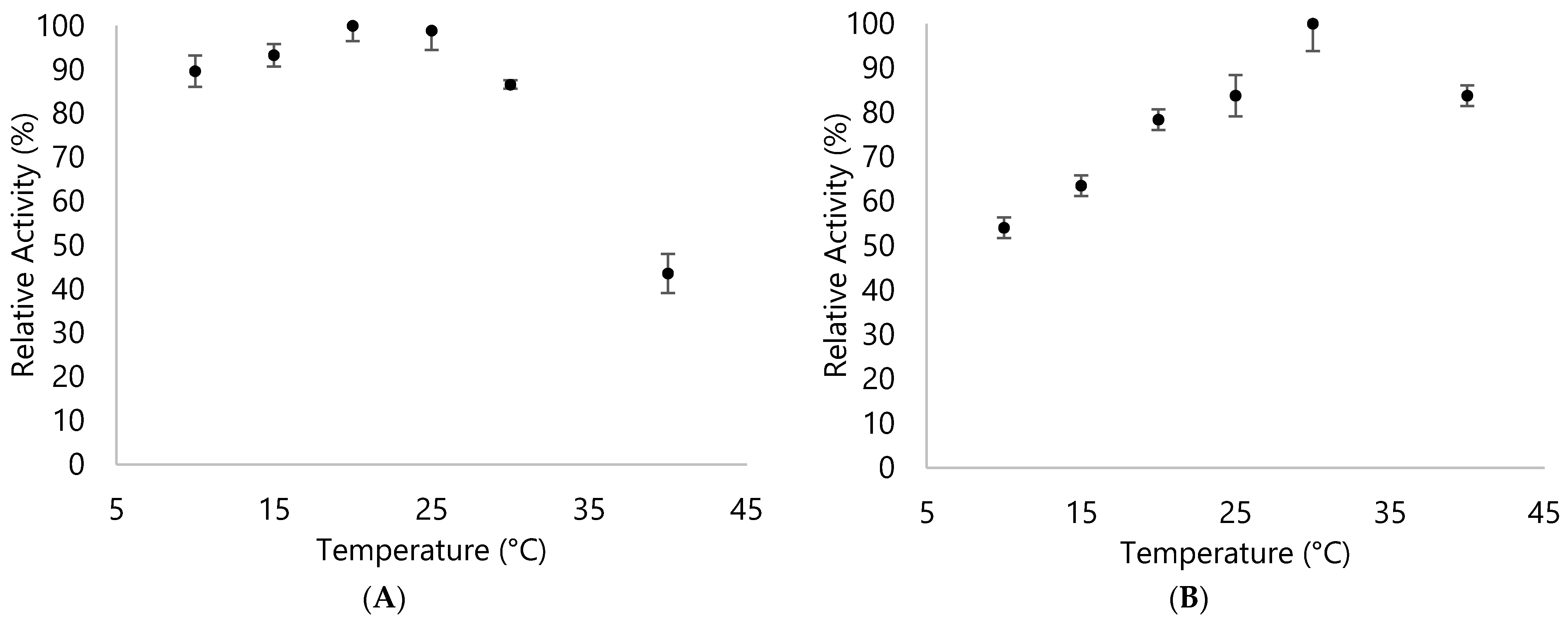
3.2. Biochemical Properties of DyPA and DyPB
3.3. Activities of DyPA and DypB with RB5 as a Substrate
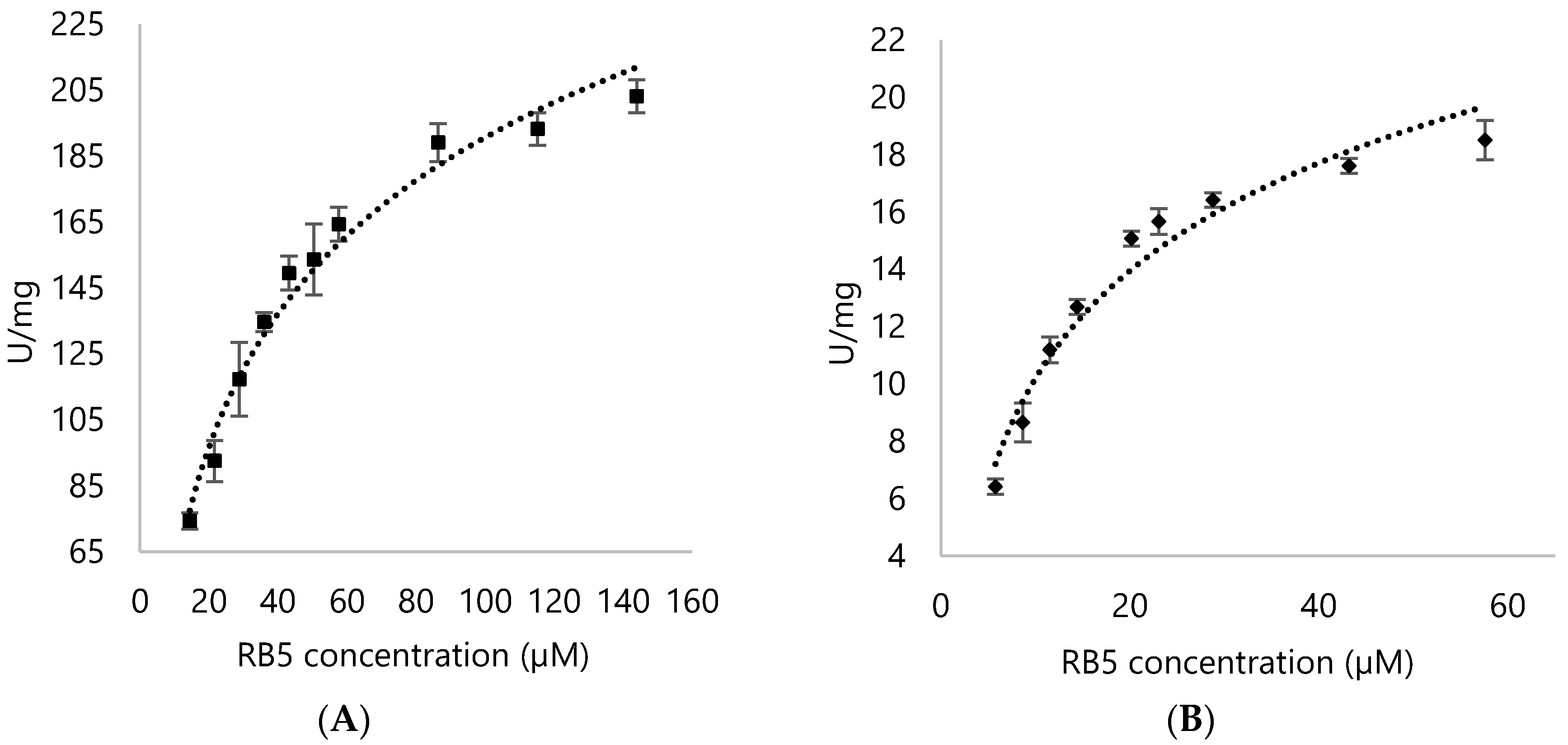
3.4. Kinetic Parameters for DypA and DypB
3.5. DypA and DypB Activities against Different Dyes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Term | Attributes | Supplier/Reference |
|---|---|---|---|
| R. opacus 1CP | - | donator; Gram-positive actinobacterium | Environmental Microbiology Group, TU Freiberg [70] |
| E. coli DH10B | - | intermediate host; blue-white-selection | Invitrogen, Karlsruhe, Germany Biological Institute, TU Dresden |
| E. coli Rosetta2 (DE3) pLysS | R2 | expression host; camR; derivative of E. coli BL21 (DE3) pLysS, additional supply of tRNAs for 7 rare codons (AGA, AGG, AUA, CUA, GGA, CCC, CGG) | MERCK Millipore, Darmstadt, Germany Strain Collection, Institute for Chemistry, TU Dresden |
| E. coli Rosetta2 (DE3) pLysS_pET16bp | R2pet | expression host; derivative of E. coli Rosetta2 (DE3) pLysS, additionally harbors pET16bp | this study |
| E. coli Rosetta2 (DE3) pLysS_pET16bp_dypA | R2A | expression host; derivative of E. coli Rosetta2 (DE3) pLysS, capable of producing recombinant DypA | this study |
| E. coli Rosetta2 (DE3) pLysS_pET16bp_dypB | R2B | expression host; derivative of E. coli Rosetta2 (DE3) pLysS, capable of producing recombinant DypA | this study |
| Plasmid | Attributes | Supplier/Reference |
|---|---|---|
| pUC19 | cloning vector, 2686 bp, bla (ampR), lacZα for blue-white selection | supplied by Dr. S. Flecks (Biochemistry, TU Dresden) |
| pUC19_dypA | plasmid derived from pUC19 containing dypA (gene of interest) | this study |
| pUC19_dypB | plasmid derived from pUC19 containing dypB (gene of interest) | this study |
| pET16bp | expression vector derived from pET16b with expanded MCS, 5740 bp, bla (ampR), T7 promoter, lacI repressor, allows expression of recombinant proteins with N-terminal His10-tag | supplied by Dr. D. Tischler (Environmental Microbiology Group, TU Freiberg) |
| pET16bp_dypA | plasmid derived from pET16bp containing dypA (gene of interest) | this study |
| pET16bp_dypB | plasmid derived from pET16bp containing dypB (gene of interest) | this study |
| Primer | Sequence 5′→3′ | Specification and Use |
|---|---|---|
| DypA_1CP_fw | CATATGATCATGACTGATCCGACCG | with NdeI restriction site, used for PCR amplification of dypA in order to clone it into pUC19 and pET16bp |
| DypA_1CP_rev | GCGGCCGCAGGCTCTACGTGAACAG | with NotI restriction site, used for PCR amplification of dypA in order to clone it into pUC19 and pET16bp |
| DypB_1CP_fw | CATATGCCAGGCCCAGTCGCGAG | with NdeI restriction site, used for PCR amplification of dypB in order to clone it into pUC19 and pET16bp |
| DypB_1CP_rev | GCGGCCGCTCATTGCGATACTCCT | with NotI restriction site, used for PCR amplification of dypB in order to clone it into pUC19 and pET16bp |
| M13 uni (-21) | TGTAAAACGACGGCCAGT | forward sequencing primer used to verify dypA in pUC19 |
| pUC19_seq_fw | CTTAACTATGCGGCATCAGAGCAGA | forward sequencing primer used to verify dypB in pUC19 |
| M13 rev (-49) | GAGCGGATAACAATTTCACACAGG | reverse sequencing primer used to verify dypA and dypB in pUC19 |
| T7 | TAATACGACTCACTATAGGG | forward sequencing primer used to verify dypA and dypB in pET16bp |
| T7 term | CTAGTTATTGCTCAGCGGT | reverse sequencing primer used to verify dypA and dypB in pET16bp |
| Domain/Phyla | Name | Species | NCBI Accession nr. |
|---|---|---|---|
| Fungi | DyP | Galactomyces geotrichum Dec1 | Q8WZK8 |
| MsP1 | Marasmius scorodonius | B0BK71 | |
| MsP1 | Marasmius scorodonius | B0BK71 | |
| MsP2 | Marasmius scorodonius | B0BK72 | |
| AjP1 | Auricularia auricula-judae 11326 | AFJ79723 | |
| PoDyp | Pleurotus ostreatus | CAK55151 | |
| EglDyp | Exidia glandulosa | AFJ79724 | |
| MegDyp | Mycena epipterygia | AFJ79725 | |
| TAP | Termitomyces albuminosus | Q8NKF3 | |
| CpoP21 | Polyporaceae sp. | P87212 | |
| Lbi2 | Laccaria bicolor S238N-H82 | EDR12662 | |
| Bfu | Botryotinia fuckeliana B05.10 | EDN26366 | |
| PplDyp | Postia placenta Mad-698-R | EED79944 | |
| AfuDyp | Aspergillus fumigatus Af293 | EAL86784 | |
| PchDyp | Penicillium chrysogenum Wisconsin 54-1255 | CAP99029 | |
| Bacteria/α-Proteo- | PdeDyp | Paracoccus denitrificans PD1222 | ABL69832 |
| RpaDyp | Rhodopseudomonas palustris BisB18 | ABD87513 | |
| OanDyp | Ochrobactrum anthropi ATCC 49188 | ABS17389 | |
| Bacteria/β-Proteo- | CteDyp | Comamonas testosteroni KF-1 | EED66859 |
| CviDyp | Chromobacterium violaceum ATCC 12472 | AAQ59612 | |
| Bacteria/γ-Proteo- | YfeX | Escherichia coli K12 | BAE76711 |
| YfeB | Escherichia coli K12 | NP_415538 | |
| TyrA | Shewanella oneidensis | Q8EIU4 | |
| AcspDyp | Acinetobacter sp. ADP1 | CAG67144 | |
| PsspDyp | Psychrobacter sp. PRwf-1 | ABQ94167 | |
| Bacteria/δ-Proteo- | PpaDyp | Plesiocystis pacifica SIR-1 | EDM76509 |
| MxaDyp | Myxococcus xanthus DK 1622 | ABF90727 | |
| Bacteria/ε-Proteo- | CcoDyp | Campylobacter concisus 13826 | EAT98288 |
| Bacteria/Actino- | DypA | Rhodococcus opacus PD630 | ZP_18275609 |
| DypA | Rhodococcus opacus B4 | YP_002783029 | |
| DypA | Rhodococcus jostii RHA1 | ABG97551 | |
| DypA | Rhodococcus opacus 1CP | ANS26740 | |
| DypB | Rhodococcus opacus PD630 | ZP_18285374 | |
| DypB | Rhodococcus opacus B4 | YP_002779326 | |
| DypB | Rhodococcus jostii RHA1 | ABG94212 | |
| DypB | Rhodococcus opacus 1CP | ANS30609 | |
| TfuDyp | Thermobifida fusca YX | Q47KB1 | |
| MvaDyp | Mycobacterium vanbaalenii PYR-1 | ABM12972 | |
| Mtu | Mycobacterium tuberculosis H37Rv | CAB09574 | |
| Bacteria/Bacteroidetes | BtDyp | Bacteroides thetaiotaomicron Vpi-5482 | 2GVK_A |
| ChuDyp | Cytophaga hutchinsonii ATCC 33406 | ABG59511 | |
| Bacteria/Firmicutes | BsuDyp | Bacillus subtilis 168 | CAB15852 |
| Bacteria/Cyano- | AnaPx | Anabaena sp. PCC 7120 | Q8YWM0 |
| AnaVT | Anabaena variabilis ATCC 29413 | Q3M5E1 | |
| SnyDyp | Cyanothece sp. PCC 8801 | YP_002373279 | |
| CyspDyp | Cyanothece sp. PCC 7424 | ACK71272 | |
| DdiDyp | Dictyostelium discoideum AX4 | EAL70759 |
| H2O2 conc. (mM) | DypA (U/mg) | DypB (U/mg) |
|---|---|---|
| 0.2 | 36.65 ± 0.75 | 26.82 ± 0.34 |
| 0.5 | 63.24 ± 6.82 | 46.67 ± 0.03 |
| 1.0 | 80.4 ± 4.72 | 70.37 ± 4.91 |
| 2.0 | 97.84 ± 3.31 | 72.43 ± 3.02 |
| 2.9 | 102.44 ± 2.75 | 80.50 ± 3.82 |
| 4.9 | 108.15 ± 2.51 | 79.63 ± 1.96 |
| 9.8 | 101.79 ± 0.65 | 58.69 ± 3.96 |
| 195.8 | 54.03 ± 4.05 | 49.69 ± 1.6 |
References
- Soetaert, W.; Vandamme, E.J. The scope and impact of industrial biotechnology. In Industrial Biotechnology: Sustainable Growth and Economic Success; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2010. [Google Scholar]
- Bornscheuer, U.T.; Buchholz, K. Highlights in biocatalysis–historical landmarks and current trends. Eng. Life Sci. 2005, 5, 309–323. [Google Scholar] [CrossRef]
- Chatha, S.A.S.; Asgher, M.; Iqbal, H.M. Enzyme-based solutions for textile processing and dye contaminant biodegradation—A review. Environ. Sci. Pollut. Res. 2017, 24, 14005–14018. [Google Scholar] [CrossRef]
- Colpa, D.I.; Fraaije, M.W.; van Bloois, E. DyP-type peroxidases: A promising and versatile class of enzymes. J. Ind. Microbiol. Biotechnol. 2014, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Eltis, L.D. The multihued palette of dye-decolorizing peroxidases. Arch. Biochem. Biophys. 2015, 574, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Hofrichter, M.; Ullrich, R. Oxidations catalyzed by fungal peroxygenases. Curr. Opin. Chem. Biol. 2014, 19, 116–125. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R.; Pecyna, M.J.; Liers, C.; Lundell, T. New and classic families of secreted fungal heme peroxidases. Appl. Microbiol. Biotechnol. 2010, 87, 871–897. [Google Scholar] [CrossRef]
- Sugano, Y. DyP-type peroxidases comprise a novel heme peroxidase family. Cell. Mol. Life Sci. 2009, 66, 1387–1403. [Google Scholar] [CrossRef] [PubMed]
- Falade, A.O.; Mabinya, L.V.; Okoh, A.I.; Nwodo, U.U. Biochemical and molecular characterization of a novel dye-decolourizing peroxidase from Raoultella ornithinolytica OKOH-1. Int. J. Biol. Macromol. 2019, 121, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Falade, A.O.; Nwodo, U.U.; Iweriebor, B.C.; Green, E.; Mabinya, L.V.; Okoh, A.I. Lignin peroxidase functionalities and prospective applications. MicrobiologyOpen 2017, 6, e00394. [Google Scholar] [CrossRef] [Green Version]
- de Gonzalo, G.; Colpa, D.I.; Habib, M.H.; Fraaije, M.W. Bacterial enzymes involved in lignin degradation. J. Biotechnol. 2016, 236, 110–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreno, A.D.; Ibarra, D.; Alvira, P.; Tomás-Pejó, E.; Ballesteros, M. A review of biological delignification and detoxification methods for lignocellulosic bioethanol production. Crit. Rev. Biotechnol. 2015, 35, 342–354. [Google Scholar] [CrossRef]
- Zakzeski, J.; Bruijnincx, P.C.; Jongerius, A.L.; Weckhuysen, B.M. The catalytic valorization of lignin for the production of renewable chemicals. Chem. Rev. 2010, 110, 3552–3599. [Google Scholar] [CrossRef] [PubMed]
- FitzPatrick, M.; Champagne, P.; Cunningham, M.F.; Whitney, R.A. A biorefinery processing perspective: Treatment of lignocellulosic materials for the production of value-added products. Bioresour. Technol. 2010, 101, 8915–8922. [Google Scholar] [CrossRef] [PubMed]
- Lambertz, C.; Ece, S.; Fischer, R.; Commandeur, U. Progress and obstacles in the production and application of recombinant lignin-degrading peroxidases. Bioengineered 2016, 7, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Conesa, A.; Punt, P.J.; van den Hondel, C.A. Fungal peroxidases: Molecular aspects and applications. J. Biotechnol. 2002, 93, 143–158. [Google Scholar] [CrossRef]
- Arnao, M.B.; Acosta, M.; del Rio, J.A.; Varon, R.; Garcia-Canovas, F. A kinetic study on the suicide inactivation of peroxidase by hydrogen peroxide. Biochim. Biophys. Acta BBA-Protein Struct. Mol. Enzymol. 1990, 1041, 43–47. [Google Scholar] [CrossRef]
- Lauber, C.; Schwarz, T.; Nguyen, Q.K.; Lorenz, P.; Lochnit, G.; Zorn, H. Identification, heterologous expression and characterization of a dye-decolorizing peroxidase of Pleurotus sapidus. AMB Express 2017, 7, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Colpa, D.I.; Lončar, N.; Schmidt, M.; Fraaije, M.W. Creating oxidase–peroxidase fusion enzymes as a toolbox for cascade reactions. ChemBioChem 2017, 18, 2226. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Grigg, J.C.; Qin, W.; Kadla, J.F.; Murphy, M.E.; Eltis, L.D. Improved manganese-oxidizing activity of DypB, a peroxidase from a lignolytic bacterium. ACS Chem. Biol. 2013, 8, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Brissos, V.; Tavares, D.; Sousa, A.C.; Robalo, M.P.; Martins, L.O. Engineering a bacterial DyP-type peroxidase for enhanced oxidation of lignin-related phenolics at alkaline pH. ACS Catal. 2017, 7, 3454–3465. [Google Scholar] [CrossRef]
- Ogola, H.J.O.; Kamiike, T.; Hashimoto, N.; Ashida, H.; Ishikawa, T.; Shibata, H.; Sawa, Y. Molecular characterization of a novel peroxidase from the cyanobacterium Anabaena sp. strain PCC 7120. Appl. Environ. Microbiol. 2009, 75, 7509–7518. [Google Scholar] [CrossRef] [Green Version]
- Min, K.; Gong, G.; Woo, H.M.; Kim, Y.; Um, Y. A dye-decolorizing peroxidase from Bacillus subtilis exhibiting substrate-dependent optimum temperature for dyes and β-ether lignin dimer. Sci. Rep. 2015, 5, 1–8. [Google Scholar]
- van Bloois, E.; Pazmiño DE, T.; Winter, R.T.; Fraaije, M.W. A robust and extracellular heme-containing peroxidase from Thermobifida fusca as prototype of a bacterial peroxidase superfamily. Appl. Microbiol. Biotechnol. 2010, 86, 1419–1430. [Google Scholar] [CrossRef] [Green Version]
- Rahmanpour, R.; Rea, D.; Jamshidi, S.; Fülöp, V.; Bugg, T.D. Structure of Thermobifida fusca DyP-type peroxidase and activity towards Kraft lignin and lignin model compounds. Arch. Biochem. Biophys. 2016, 594, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lončar, N.; Colpa, D.I.; Fraaije, M.W. Exploring the biocatalytic potential of a DyP-type peroxidase by profiling the substrate acceptance of Thermobifida fusca DyP peroxidase. Tetrahedron 2016, 72, 7276–7281. [Google Scholar] [CrossRef]
- Brown, M.E.; Barros, T.; Chang, M.C. Identification and characterization of a multifunctional dye peroxidase from a lignin-reactive bacterium. ACS Chem. Biol. 2012, 7, 2074–2081. [Google Scholar] [CrossRef] [PubMed]
- Sainsbury, P.D.; Hardiman, E.M.; Ahmad, M.; Otani, H.; Seghezzi, N.; Eltis, L.D.; Bugg, T.D. Breaking down lignin to high-value chemicals: The conversion of lignocellulose to vanillin in a gene deletion mutant of Rhodococcus jostii RHA1. ACS Chem. Biol. 2013, 8, 2151–2156. [Google Scholar] [CrossRef]
- Roberts, J.N.; Singh, R.; Grigg, J.C.; Murphy, M.E.; Bugg, T.D.; Eltis, L.D. Characterization of dye-decolorizing peroxidases from Rhodococcus jostii RHA1. Biochemistry 2011, 50, 5108–5119. [Google Scholar] [CrossRef]
- Ahmad, M.; Roberts, J.N.; Hardiman, E.M.; Singh, R.; Eltis, L.D.; Bugg, T.D. Identification of DypB from Rhodococcus jostii RHA1 as a lignin peroxidase. Biochemistry 2011, 50, 5096–5107. [Google Scholar] [CrossRef]
- Yu, W.; Liu, W.; Huang, H.; Zheng, F.; Wang, X.; Wu, Y.; Li, K.; Xie, X.; Jin, Y. Application of a novel alkali-tolerant thermostable DyP-type peroxidase from Saccharomonospora viridis DSM 43017 in biobleaching of eucalyptus kraft pulp. PLoS ONE 2014, 9, e110319. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Liu, C.; Li, B.; Yuan, H.; Yang, J.; Zheng, B. Identification and molecular characterization of a novel DyP-type peroxidase from Pseudomonas aeruginosa PKE117. Appl. Biochem. Biotechnol. 2012, 166, 774–785. [Google Scholar] [CrossRef]
- Rahmanpour, R.; Bugg, T.D. Characterisation of Dyp-type peroxidases from Pseudomonas fluorescens Pf-5: Oxidation of Mn (II) and polymeric lignin by Dyp1B. Arch. Biochem. Biophys. 2015, 574, 93–98. [Google Scholar] [CrossRef]
- Santos, A.; Mendes, S.; Brissos, V.; Martins, L.O. New dye-decolorizing peroxidases from Bacillus subtilis and Pseudomonas putida MET94: Towards biotechnological applications. Appl. Microbiol. Biotechnol. 2014, 98, 2053–2065. [Google Scholar] [CrossRef] [PubMed]
- Létoffé, S.; Heuck, G.; Delepelaire, P.; Lange, N.; Wandersman, C. Bacteria capture iron from heme by keeping tetrapyrrol skeleton intact. Proc. Natl. Acad. Sci. USA 2009, 106, 11719–11724. [Google Scholar] [CrossRef] [Green Version]
- Tatsumi, N.; Inui, M. (Eds.) Corynebacterium glutamicum: Biology and Biotechnology; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Volume 23. [Google Scholar]
- Hwang, K.S.; Kim, H.U.; Charusanti, P.; Palsson, B.Ø.; Lee, S.Y. Systems biology and biotechnology of Streptomyces species for the production of secondary metabolites. Biotechnol. Adv. 2014, 32, 255–268. [Google Scholar] [CrossRef] [PubMed]
- Cappelletti, M.; Presentato, A.; Piacenza, E.; Firrincieli, A.; Turner, R.J.; Zannoni, D. Biotechnology of Rhodococcus for the production of valuable compounds. Appl. Microbiol. Biotechnol. 2020, 104, 8567–8594. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, H.M.; Mayer, F.; Fabritius, D.; Steinbüchel, A. Formation of intracytoplasmic lipid inclusions by Rhodococcus opacus strain PD630. Arch. Microbiol. 1996, 165, 377–386. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhao, L.; Zan, X.; Song, Y.; Ratledge, C. Boosting fatty acid synthesis in Rhodococcus opacus PD630 by overexpression of autologous thioesterases. Biotechnol. Lett. 2016, 38, 999–1008. [Google Scholar] [CrossRef]
- Na, K.S.; Kuroda, A.; Takiguchi, N.; Ikeda, T.; Ohtake, H.; Kato, J. Isolation and characterization of benzene-tolerant Rhodococcus opacus strains. J. Biosci. Bioeng. 2005, 99, 378–382. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, S.; Satoi, M.; Iwasa, Y.; Honda, K.; Sameshima, Y.; Omasa, T.; Kato, J.; Ohtake, H. Utilization of hydrophobic bacterium Rhodococcus opacus B-4 as whole-cell catalyst in anhydrous organic solvents. Appl. Microbiol. Biotechnol. 2007, 74, 761–767. [Google Scholar] [CrossRef]
- Tischler, D.; Eulberg, D.; Lakner, S.; Kaschabek, S.R.; van Berkel, W.J.; Schlomann, M. Identification of a novel self-sufficient styrene monooxygenase from Rhodococcus opacus 1CP. J. Bacteriol. 2009, 191, 4996–5009. [Google Scholar] [CrossRef] [Green Version]
- Oelschlägel, M.; Gröning, J.A.; Tischler, D.; Kaschabek, S.R.; Schlömann, M. Styrene oxide isomerase of Rhodococcus opacus 1CP, a highly stable and considerably active enzyme. Appl. Environ. Microbiol. 2012, 78, 4330–4337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gröning, J.A.; Eulberg, D.; Tischler, D.; Kaschabek, S.R.; Schlömann, M. Gene redundancy of two-component (chloro) phenol hydroxylases in Rhodococcus opacus 1CP. FEMS Microbiol. Lett. 2014, 361, 68–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, J.; Schlömann, M.; Tischler, D. Biochemical characterization of an azoreductase from Rhodococcus opacus 1CP possessing methyl red degradation ability. J. Mol. Catal. B Enzym. 2016, 130, 9–17. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Shrestha, R.; Huang, G.; Meekins, D.A.; Geisbrecht, B.V.; Li, P. Mechanistic insights into dye-decolorizing peroxidase revealed by solvent isotope and viscosity effects. ACS Catal. 2017, 7, 6352–6364. [Google Scholar] [CrossRef] [Green Version]
- Graves, P.E.; Henderson, D.P.; Horstman, M.J.; Solomon, B.J.; Olson, J.S. Enhancing stability and expression of recombinant human hemoglobin in E. coli: Progress in the development of a recombinant HBOC source. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2008, 1784, 1471–1479. [Google Scholar] [CrossRef] [Green Version]
- Sudhamsu, J.; Kabir, M.; Airola, M.V.; Patel, B.A.; Yeh, S.R.; Rousseau, D.L.; Crane, B.R. Co-expression of ferrochelatase allows for complete heme incorporation into recombinant proteins produced in E. coli. Protein Expr. Purif. 2010, 73, 78–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.T.; Santama, N.; Dacey, S.; Edwards, M.; Bray, R.C.; Thorneley, R.N.; Burke, J.F. Expression of a synthetic gene for horseradish peroxidase C in Escherichia coli and folding and activation of the recombinant enzyme with Ca2+ and heme. J. Biol. Chem. 1990, 265, 13335–13343. [Google Scholar] [CrossRef]
- Kery, V.; Elleder, D.; Kraus, J.P. δ-Aminolevulinate increases heme saturation and yield of human cystathionine β-synthase expressed in Escherichia coli. Arch. Biochem. Biophys. 1995, 316, 24–29. [Google Scholar] [CrossRef] [PubMed]
- Weickert, M.J.; Pagratis, M.; Curry, S.R.; Blackmore, R. Stabilization of apoglobin by low temperature increases yield of soluble recombinant hemoglobin in Escherichia coli. Appl. Environ. Microbiol. 1997, 63, 4313–4320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varnado, C.L.; Goodwin, D.C. System for the expression of recombinant hemoproteins in Escherichia coli. Protein Expr. Purif. 2004, 35, 76–83. [Google Scholar] [CrossRef]
- Sugano, Y.; Ishii, Y.; Shoda, M. Role of H164 in a unique dye-decolorizing heme peroxidase DyP. Biochem. Biophys. Res. Commun. 2004, 322, 126–132. [Google Scholar] [CrossRef]
- Chen, C.; Li, T. Bacterial dye-decolorizing peroxidases: Biochemical properties and biotechnological opportunities. Phys. Sci. Rev. 2016, 1, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Chaplin, A.K.; Chicano, T.M.; Hampshire, B.V.; Wilson, M.T.; Hough, M.A.; Svistunenko, D.A.; Worrall, J.A. An Aromatic Dyad Motif in Dye Decolourising Peroxidases Has Implications for Free Radical Formation and Catalysis. Chem.–A Eur. J. 2019, 25, 6141–6153. [Google Scholar] [CrossRef] [Green Version]
- Catucci, G.; Valetti, F.; Sadeghi, S.J.; Gilardi, G. Biochemical features of dye-decolorizing peroxidases: Current impact on lignin degradation. Biotechnol. Appl. Biochem. 2020, 67, 751–759. [Google Scholar] [CrossRef]
- Sugano, Y.; Muramatsu, R.; Ichiyanagi, A.; Sato, T.; Shoda, M. DyP, a unique dye-decolorizing peroxidase, represents a novel heme peroxidase family: ASP171 replaces the distal histidine of classical peroxidases. J. Biol. Chem. 2007, 282, 36652–36658. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, K.; Nishihashi, Y.; Narioka, T.; Yoshida, T.; Morita, M.; Sugano, Y. Characterization of a novel DyP-type peroxidase from Streptomyces avermitilis. J. Biosci. Bioeng. 2017, 123, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Wariishi, H.; Gold, M.H. Lignin peroxidases Compound III. J. Biol. Chem. 1989, 265, 2070–2077. [Google Scholar] [CrossRef]
- de Eugenio, L.I.; Peces-Pérez, R.; Linde, D.; Prieto, A.; Barriuso, J.; Ruiz-Dueñas, F.J.; Martínez, M.J. Characterization of a Dye-Decolorizing Peroxidase from Irpex lacteus Expressed in Escherichia coli: An Enzyme with Wide Substrate Specificity Able to Transform Lignosulfonates. J. Fungi 2021, 7, 325. [Google Scholar] [CrossRef] [PubMed]
- Valderrama, B.; Ayala, M.; Vazquez-Duhalt, R. Suicide inactivation of peroxidases and the challenge of engineering more robust enzymes. Chem. Biol. 2002, 9, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Liers, C.; Bobeth, C.; Pecyna, M.; Ullrich, R.; Hofrichter, M. DyP-like peroxidases of the jelly fungus Auricularia auricula-judae oxidize nonphenolic lignin model compounds and high-redox potential dyes. Appl. Microbiol. Biotechnol. 2010, 85, 1869–1879. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, F.; Kajino, T.; Sugiyama, H.; Asami, O.; Takahashi, H. Thermally stable and hydrogen peroxide tolerant manganese peroxidase (MnP) from Lenzites betulinus. FEBS Lett. 2002, 530, 249–252. [Google Scholar] [CrossRef] [Green Version]
- Sugano, Y.; Matsushima, Y.; Tsuchiya, K.; Aoki, H.; Hirai, M.; Shoda, M. Degradation pathway of an anthraquinone dye catalyzed by a unique peroxidase DyP from Thanatephorus cucumeris Dec 1. Biodegradation 2009, 20, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Lončar, N.; Drašković, N.; Božić, N.; Romero, E.; Simić, S.; Opsenica, I.; Vujčić, Z.; Fraaije, M.W. Expression and Characterization of a Dye-decolorizing Peroxidase from Pseudomonas fluorescens Pf0-1. Catalysts 2019, 9, 463. [Google Scholar] [CrossRef] [Green Version]
- Gorlatov, S.N.; Maltseva, O.V.; Shevchenko, V.I.; Golovleva, L.A. Degradation of chlorophenols by a culture of Rhodococcus erythropolis. Microbiology 1989, 58, 647–651. [Google Scholar]


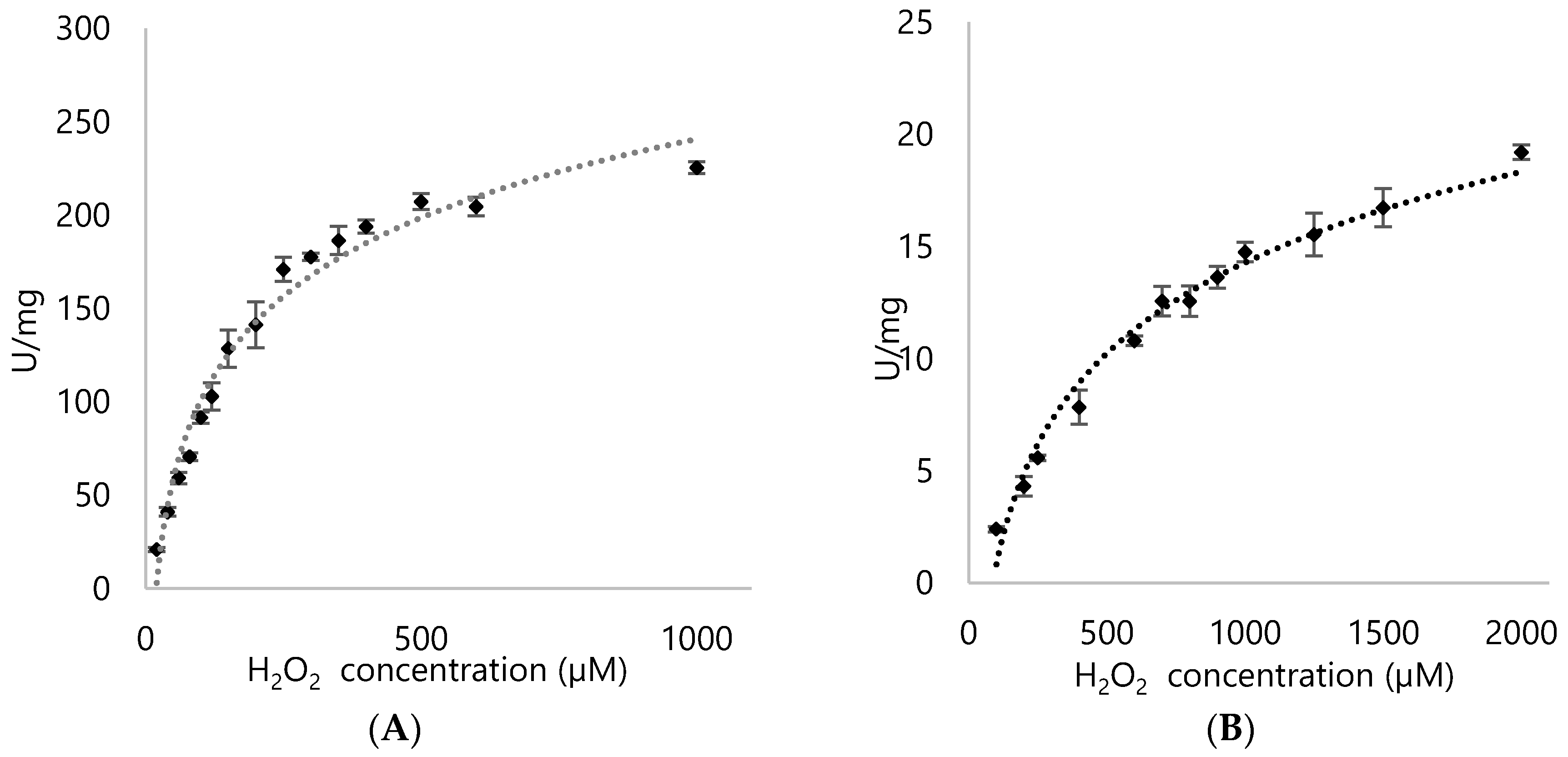
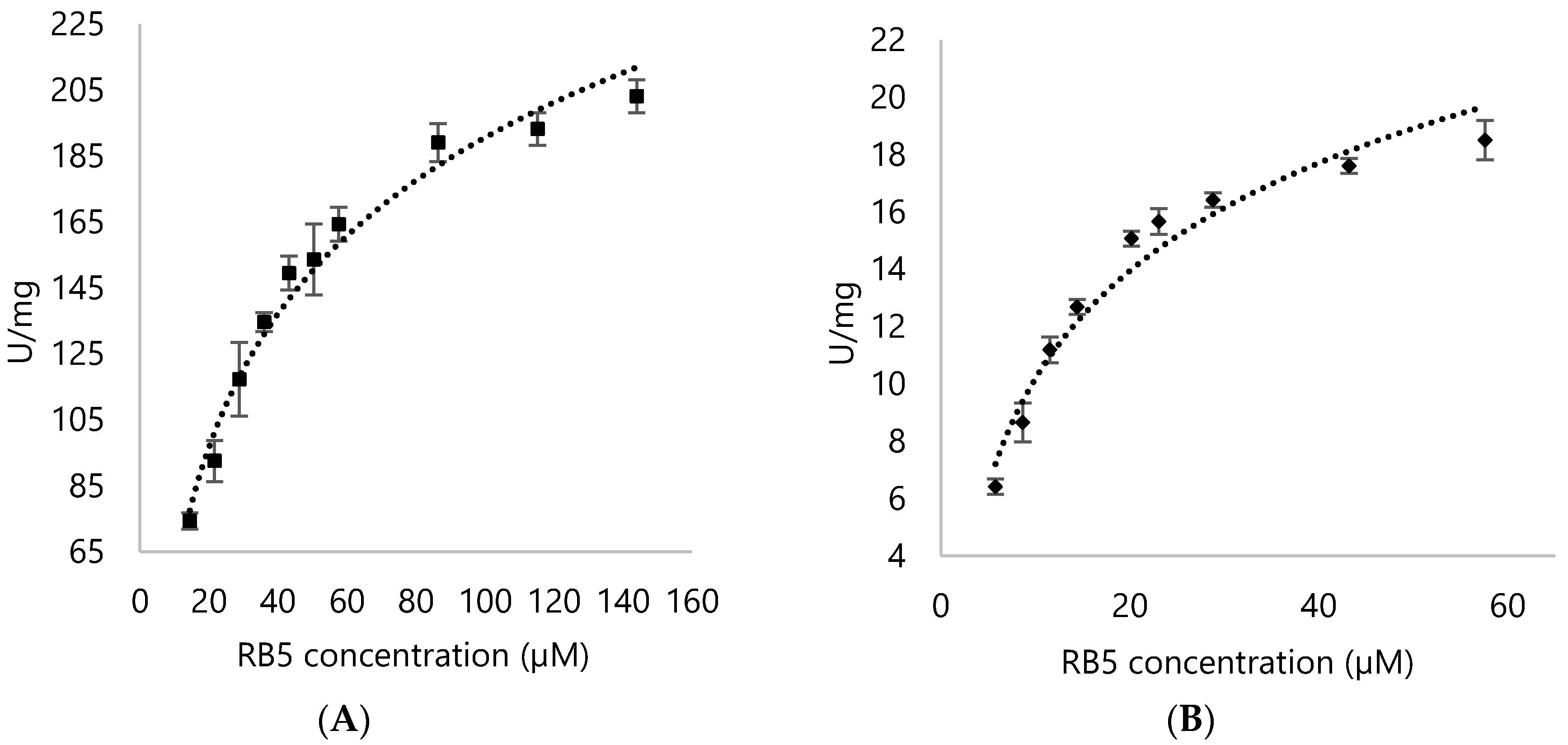
| Enzyme | Substrate | Km (µM) | Vmax (U/mg) | Kcat (s−1) | Kcat/Km (µM/s−1) |
|---|---|---|---|---|---|
| DypA | H2O2 | 196.89 ± 16.95 | 282.39 ± 9.52 | 228.17 ± 7.69 | 1.16 ± 0.45 |
| RB5 | 33.34 ± 2.37 | 254.6 ± 6.69 | 205.72 ± 5.41 | 6.17 ± 2.28 | |
| DypB | H2O2 | 1011.8 ± 78.15 | 28.74 ± 1.11 | 19.07 ± 0.89 | 0.018 ± 0.011 |
| RB5 | 12.78 ± 1.15 | 23.21 ± 0.96 | 15.47 ± 0.78 | 1.21 ± 0.68 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ngo, A.C.R.; Conrad, C.; Gómez Baraibar, Á.; Matura, A.; van Pée, K.-H.; Tischler, D. Characterization of Two Hydrogen Peroxide Resistant Peroxidases from Rhodococcus opacus 1CP. Appl. Sci. 2021, 11, 7941. https://doi.org/10.3390/app11177941
Ngo ACR, Conrad C, Gómez Baraibar Á, Matura A, van Pée K-H, Tischler D. Characterization of Two Hydrogen Peroxide Resistant Peroxidases from Rhodococcus opacus 1CP. Applied Sciences. 2021; 11(17):7941. https://doi.org/10.3390/app11177941
Chicago/Turabian StyleNgo, Anna Christina R., Catleen Conrad, Álvaro Gómez Baraibar, Anke Matura, Karl-Heinz van Pée, and Dirk Tischler. 2021. "Characterization of Two Hydrogen Peroxide Resistant Peroxidases from Rhodococcus opacus 1CP" Applied Sciences 11, no. 17: 7941. https://doi.org/10.3390/app11177941
APA StyleNgo, A. C. R., Conrad, C., Gómez Baraibar, Á., Matura, A., van Pée, K.-H., & Tischler, D. (2021). Characterization of Two Hydrogen Peroxide Resistant Peroxidases from Rhodococcus opacus 1CP. Applied Sciences, 11(17), 7941. https://doi.org/10.3390/app11177941