
Phage-Phenotype Imaging of Myeloma Plasma Cells by Phage Display

 ,
,  ,
,  ,
,  , , ,
, , ,  and
and 
Abstract
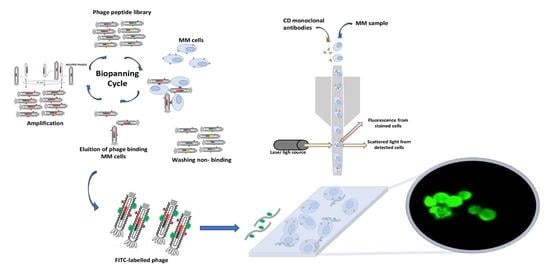
:
1. Introduction
2. Materials and Methods
2.1. Bacteriophage, Bacteria Host, and Eukaryotic Cells Line
2.2. Phage Selection and Binding Affinity
2.3. DNA Sequencing and Peptides Analysis
2.4. Labelling of Phage with FITC
2.5. Sample Collection
2.6. Isolation of Myeloma Plasma Cells from Bone Marrow
2.7. Sample Preparation for Fluorescence Imaging
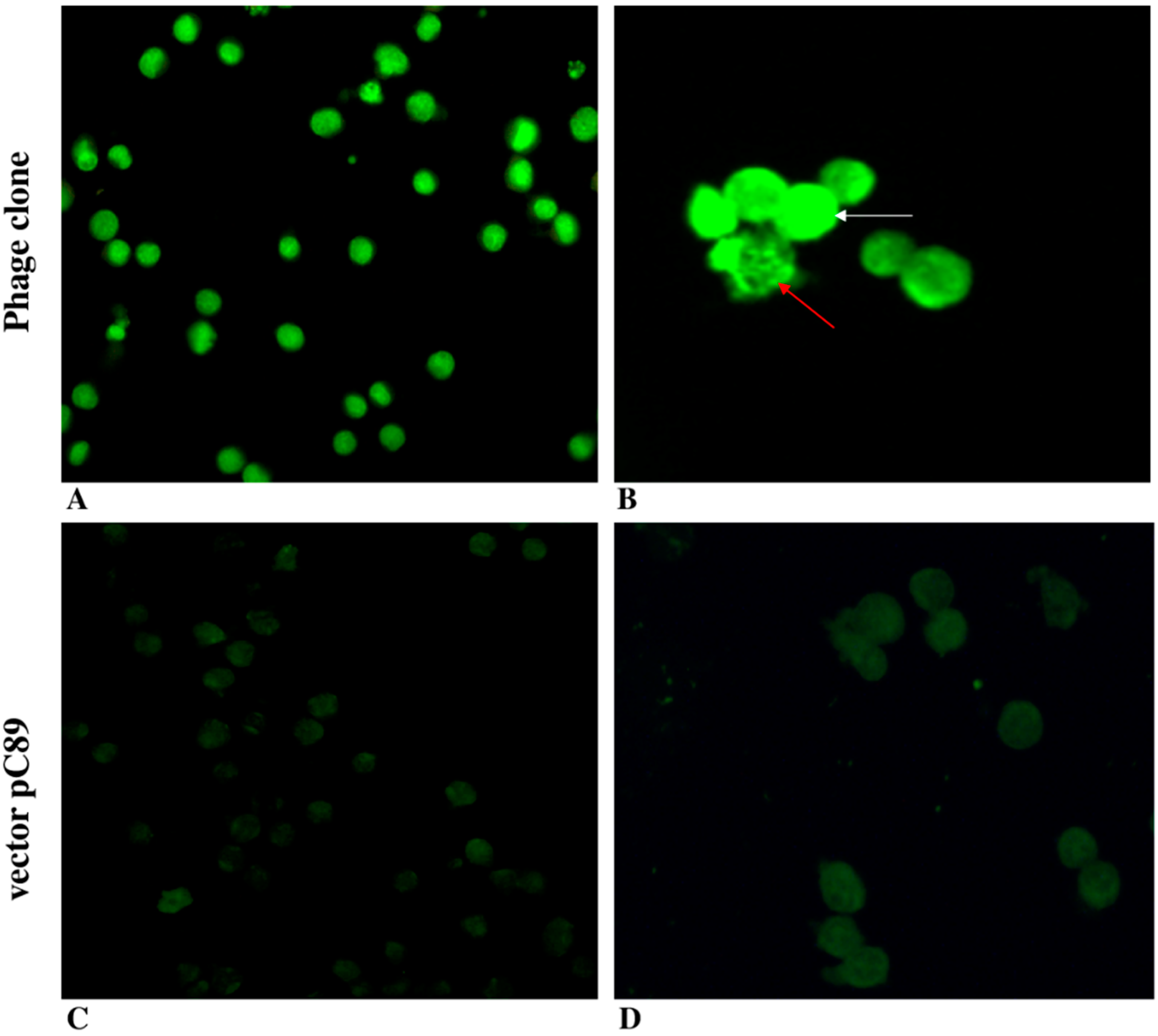
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Zagozdzon, R.; Golab, J. Cancer stem cells in haematological malignancies. Wspolczesna Onkol. 2015, 19, A1–A6. [Google Scholar] [CrossRef]
- Rawstron, A.C.; Davies, F.E.; Das Gupta, R.; Ashcroft, A.J.; Patmore, R.; Drayson, M.T.; Owen, R.G.; Jack, A.S.; Child, J.A.; Morgan, G.J. Flow cytometric disease monitoring in multiple myeloma: The relationship between normal and neoplastic plasma cells predicts outcome after transplantation. Blood 2002, 100, 3095–3100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- San Miguel, J.F.; Almeida, J.; Mateo, G.; Bladé, J.; López-Berges, C.; Caballero, D.; Hernández, J.; Moro, M.J.; Fernández-Calvo, J.; Díaz-Mediavilla, J.; et al. Immunophenotypic evaluation of the plasma cell compartment in multiple myeloma: A tool for comparing the efficacy of different treatment strategies and predicting outcome. Blood 2002, 99, 1853–1856. [Google Scholar] [CrossRef] [PubMed]
- Flores-Montero, J.; de Tute, R.; Paiva, B.; Perez, J.J.; Bottcher, S.; Wind, H.; Sanoja, L.; Puig, N.; Lecrevisse, Q.; Vidriales, M.B.; et al. Immunophenotype of normal vs. myeloma plasma cells: Toward antibody panel specifications for MRD detection in multiple myeloma. Cytom. B Clin. Cytom. 2016, 90, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Fumey, W.; Koenigsdorf, J.; Kunick, V.; Menzel, S.; Schütze, K.; Unger, M.; Schriewer, L.; Haag, F.; Adam, G.; Oberle, A.; et al. Nanobodies effectively modulate the enzymatic activity of CD38 and allow specific imaging of CD38+ tumors in mouse models in vivo. Sci. Rep. 2017, 7, 14289. [Google Scholar] [CrossRef] [PubMed]
- Akhmetzyanova, I.; McCarron, M.J.; Parekh, S.; Chesi, M.; Bergsagel, P.L.; Fooksman, D.R. Dynamic CD138 surface expression regulates switch between myeloma growth and dissemination. Leukemia 2020, 34, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Palaiologou, M.; Delladetsima, I.; Tiniakos, D. CD138 (syndecan-1) expression in health and disease. Histol. Histopathol. 2014, 29, 177–189. [Google Scholar] [CrossRef]
- Coombe, D.R. Biological implications of glycosaminoglycan interactions with haemopoietic cytokines. Immunol. Cell Biol. 2008, 86, 598–607. [Google Scholar] [CrossRef]
- Casu, B.; Naggi, A.; Torri, G. Heparin-derived heparan sulfate mimics to modulate heparan sulfate-protein interaction in inflammation and cancer. Matrix Biol. 2010, 29, 442–452. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Mahendravada, A.; Ballard, B.; Kale, B.; Ramos, C.; West, J.; Maguire, T.; McKay, K.; Lichtman, E.; Tuchman, S.; et al. Safety and efficacy of targeting CD138 with a chimeric antigen receptor for the treatment of multiple myeloma. Oncotarget 2019, 10, 2369–2383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, F.; Dalla Palma, B.; Giuliani, N. CD38 Expression by Myeloma Cells and Its Role in the Context of Bone Marrow Microenvironment: Modulation by Therapeutic Agents. Cells 2019, 8, 1632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deaglio, S.; Aydin, S.; Vaisitti, T.; Bergui, L.; Malavasi, F. CD38 at the junction between prognostic marker and therapeutic target. Trends Mol. Med. 2008, 14, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Menke, D.M.; Horny, H.P.; Griesser, H.; Atkinson, E.J.; Kaiserling, E.; Kyle, R.A. Immunophenotypic and genotypic characterisation of multiple myelomas with adverse prognosis characterised by immunohistological expression of the T cell related antigen CD45RO (UCHL-1). J. Clin. Pathol 1998, 51, 432–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rheinländer, A.; Schraven, B.; Bommhardt, U. CD45 in human physiology and clinical medicine. Immunol. Lett. 2018, 196, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Dass, J.; Arava, S.; Mishra, P.C.; Dinda, A.K.; Pati, H.P. Role of CD138, CD56, and light chain immunohistochemistry in suspected and diagnosed plasma cell myeloma: A prospective study. South Asian J. Cancer. 2019, 8, 60–64. [Google Scholar] [CrossRef]
- Gonsalves, W.I.; Timm, M.M.; Rajkumar, S.V.; Morice, W.G.; Dispenzieri, A.; Buadi, F.K.; Lacy, M.Q.; Dingli, D.; Leung, N.; Kapoor, P.; et al. The prognostic significance of CD45 expression by clonal bone marrow plasma cells in patients with newly diagnosed multiple myeloma. Leuk. Res. 2016, 44, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Rajkumar, S.V.; Kimlinger, T.; Greipp, P.R.; Witzig, T.E. CD45 expression by bone marrow plasma cells in multiple myeloma: Clinical and biological correlations. Leukemia 2015, 19, 1466–1470. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.P.; Petrenko, V.A. Phage display. Chem. Rev. 1997, 97, 391–410. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Liu, I.J.; Lu, R.M.; Wu, H.C. Advancement and applications of peptide phage display technology in biomedical science. J. Biomed. Sci. 2016, 23, 8. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, J.W.; Yang, L.; De Plano, L.M.; Stackhouse, M.A.; Petrenko, V.A. Evolution of a Landscape Phage Library in a Mouse Xenograft Model of Human Breast Cancer. Viruses 2019, 11, 988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morag, O.; Sgourakis, N.G.; Baker, D.; Goldbourt, A. The NMR-Rosetta capsid model of M13 bacteriophage reveals a quadrupled hydrophobic packing epitope. Proc. Natl. Acad. Sci. USA 2015, 112, 971–976. [Google Scholar] [CrossRef] [Green Version]
- Jaye, D.L.; Geigerman, C.M.; Fuller, R.E.; Akyildiz, A.; Parkos, C.A. Direct fluorochrome labeling of phage display library clones for studying binding specificities: Applications in flow cytometry and fluorescence microscopy. J. Immunol. Methods 2004, 295, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Kelly, K.A.; Waterman, P.; Weissleder, R. In vivo imaging of molecularly targeted phage. Neoplasia 2006, 8, 1011–1018. [Google Scholar] [CrossRef] [Green Version]
- Li, K.; Chen, Y.; Li, S.; Nguyen, H.G.; Niu, Z.; You, S.; Mello, C.M.; Lu, X.; Wang, Q. Chemical modification of M13 bacteriophage and its application in cancer cell imaging. Bioconjug. Chem. 2010, 21, 1369–1377. [Google Scholar] [CrossRef] [PubMed]
- Torrisi, L.; Guglielmino, S.; Silipigni, L.; De Plano, L.M.; Kovacik, L.; Lavrentiev, V.; Torrisi, A.; Fazio, M.; Fazio, B.; Di Marco, G. Study of gold nanoparticle transport by M13 phages towards disease tissues as targeting procedure for radiotherapy applications. Gold Bull. 2019, 52, 135–144. [Google Scholar] [CrossRef]
- Lee, L.A.; Niu, Z.; Wang, Q. Viruses and virus-like protein assemblies-Chemically programmable nanoscale building blocks. Nano Res. 2009, 2, 349–364. [Google Scholar] [CrossRef] [Green Version]
- De Plano, L.M.; Scibilia, S.; Rizzo, M.G.; Crea, S.; Franco, D.; Mezzasalma, A.M.; Guglielmino, S.P. One-step production of phage–silicon nanoparticles by PLAL as fluorescent nanoprobes for cell identification. Appl. Phys. A 2018, 124, 222. [Google Scholar] [CrossRef]
- Rakonjac, J.; Bennett, N.J.; Spagnuolo, J.; Gagic, D.; Russel, M. Filamentous bacteriophage: Biology, phage display and nanotechnology applications. Curr. Issues Mol. Biol. 2011, 13, 51–76. [Google Scholar] [PubMed]
- De Plano, L.M.; Fazio, E.; Rizzo, M.G.; Franco, D.; Carnazza, S.; Trusso, S.; Neri, F.; Guglielmino, S.P. Phage-based assay for rapid detection of bacterial pathogens in blood by Raman spectroscopy. J. Immunol. Methods 2019, 465, 45–52. [Google Scholar] [CrossRef]
- Franco, D.; De Plano, L.M.; Rizzo, M.G.; Scibilia, S.; Lentini, G.; Fazio, E.; Neri, F.; Guglielmino, S.P.P.; Mezzasalma, A.M. Bio-hybrid gold nanoparticles as SERS probe for rapid bacteria cell identification. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 224, 117394. [Google Scholar] [CrossRef] [PubMed]
- Felici, F.; Castagnoli, L.; Musacchio, A.; Jappelli, R.; Cesareni, G.J. Selection of antibody ligands from a large library of oligopeptides expressed on a multivalent exposition vector. Mol. Biol. 1991, 222, 301–310. [Google Scholar] [CrossRef]
- Lentini, G.; Fazio, E.; Calabrese, F.; De Plano, L.M.; Puliafico, M.; Franco, D.; Nicolò, M.S.; Carnazza, S.; Trusso, S.; Allegra, A.; et al. Phage–AgNPs complex as SERS probe for U937 cell identification. Biosens. Bioelectron. 2015, 74, 398–405. [Google Scholar] [CrossRef]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Gouw, M.; Michael, S.; Sámano-Sánchez, H.; Kumar, M.; Zeke, A.; Lang, B.; Bely, B.; Chemes, L.B.; Davey, N.E.; Deng, Z.; et al. The eukaryotic linear motif resource—2018 update. Nucleic Acids Res. 2018, 46, D428–D434. [Google Scholar] [CrossRef] [PubMed]
- Petrenko, V.A.; Gillespie, J.W.; Xu, H.; O’Dell, T.; De Plano, L.M. Combinatorial Avidity Selection of Mosaic Landscape Phages Targeted at Breast Cancer Cells-An Alternative Mechanism of Directed Molecular Evolution. Viruses 2019, 11, 785. [Google Scholar] [CrossRef] [Green Version]
- De Plano, L.M.; Carnazza, S.; Franco, D.; Rizzo, M.G.; Conoci, S.; Petralia, S.; Nicoletti, A.; Zappia, M.; Campolo, M.; Esposito, E.; et al. Innovative IgG biomarkers based on phage display microbial amyloid mimotope for state and stage diagnosis in Alzheimer’s disease. ACS Chem. Neurosci. 2020, 11, 1013–1026. [Google Scholar] [CrossRef]
- Herman, R.E.; Makienko, E.G.; Prieve, M.G.; Fuller, M.; Houston, M.E., Jr.; Johnson, P.H. Phage display screening of epithelial cell monolayers treated with EGTA: Identification of peptide FDFWITP that modulates tight junction activity. J. Biomol. Screen. 2007, 12, 1092–1101. [Google Scholar] [CrossRef]
- Paíno, T.; Sarasquete, M.E.; Paiva, B.; Krzeminski, P.; San-Segundo, L.; Corchete, L.A.; Redondo, A.; Garayoa, M.; García-Sanz, R.; Gutiérrez, N.C.; et al. Phenotypic, genomic and functional characterization reveals no differences between CD138++ and CD138 low subpopulations in multiple myeloma cell lines. PLoS ONE 2014, 9, e92378. [Google Scholar] [CrossRef] [PubMed]
- Greenstein, S.; Krett, N.L.; Kurosawa, Y.; Ma, C.; Chauhan, D.; Hideshima, T.; Anderson, K.C.; Rosen, S.T. Characterization of the MM.1 human multiple myeloma (MM) cell lines: A model system to elucidate the characteristics, behavior, and signaling of steroid-sensitive and -resistant MM cells. Exp. Hematol. 2003, 31, 271–282. [Google Scholar] [CrossRef]
- Chang, H.; Samiee, S.; Yi, Q.L. Prognostic relevance of CD56 expression in multiple myeloma: A study including 107 cases treated with high-dose melphalan-based chemotherapy and autologous stem cell transplant. Leuk. Lymphoma 2006, 47, 43–47. [Google Scholar] [CrossRef]
- Mateo, G.; Castellanos, M.; Rasillo, A.; Gutierrez, N.C.; Montalban, M.A.; Martin, M.L.; Hernandez, J.M.; Lopez-Berges, M.C.; Montejano, L.; Blade, J.; et al. Genetic abnormalities and patterns of antigenic expression in multiple myeloma. Clin. Cancer Res. 2005, 11, 3661–3667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palumbo, A.; Avet-Loiseau, H.; Oliva, S.; Lokhorst, H.M.; Goldschmidt, H.; Rosinol, L.; Richardson, P.; Caltagirone, S.; Lahuerta, J.J.; Facon, T.; et al. Revised International Staging System for Multiple Myeloma: A Report from International Myeloma Working Group. J. Clin. Oncol. 2015, 33, 2863–2869. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, K.; Suzuki, K. CD56 for Multiple Myeloma: Lack of CD56 May Be Associated with Worse Prognosis. Acta Haematol. 2018, 140, 40–41. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Zhu, Z.M.; Sun, K.; Lei, P.C.; Liu, Z.W.; Guo, J.M.; Yang, J.; Zang, Y.Z.; Zhang, Y. Expression of CD45 in newly diagnosed multiple myeloma and the relationship with prognosis. Zhonghua Xue Ye Xue Za Zhi 2019, 40, 744–749. (In Chinese) [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family 1 | Family 2 | Family 3 | ||||
|---|---|---|---|---|---|---|
| Sequence | * N | Sequence | * N | Sequence | * N | |
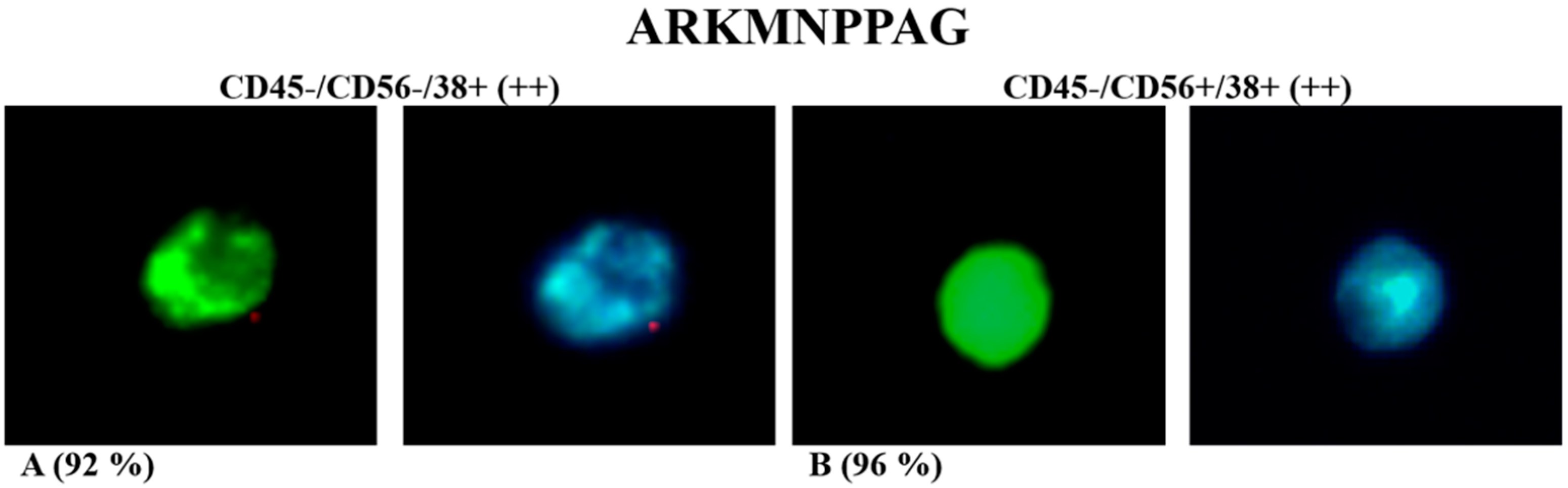
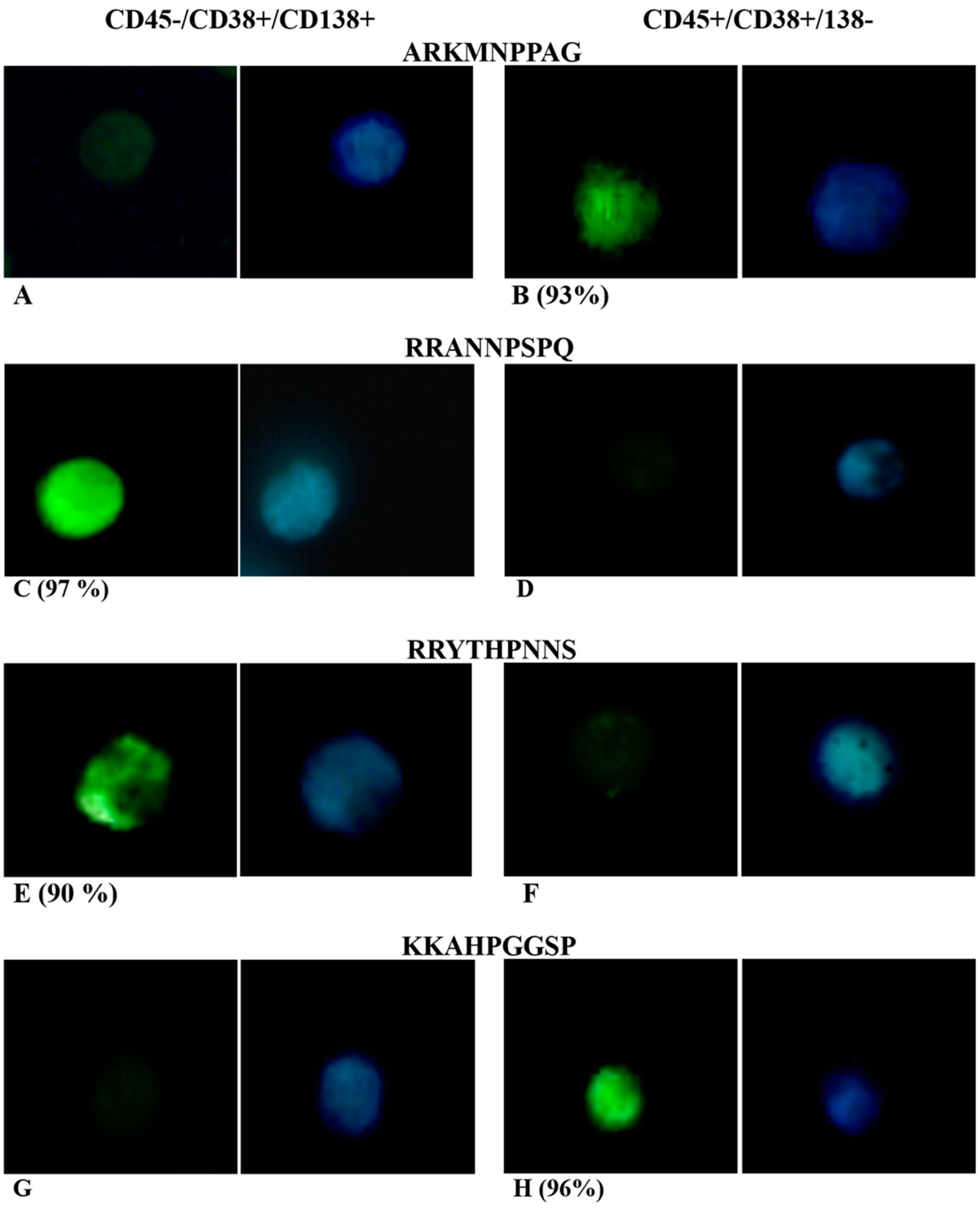
| -ARKMNPPAG | 3 | NRKIAPGNQ | 1 | -KKAHPGGSP | 3 | |
| WARKVHSPT- | 1 | NRKGTSTNL | 1 | MKKAWPGRA- | 2 | |
| -ARRTGSNPE | 1 | RRANNPSPQ | 3 | QRRAGPVPP- | 1 | |
| -ARRLLQARV | 1 | RRLFTPPRQ | 1 | -KRARHAAEG | 1 | |
| -GRKAANASS | 1 | RRYTHPNNS | 4 | |||
| Consensus | -ARK---P-- | RRA--P--Q | -KKA-PG--- | |||
| RRY--P-N- | ||||||
| Phage Clone | Cluster of Differentiation (CD) | |||
|---|---|---|---|---|
| CD45 | CD56 | CD38 | CD138 | |
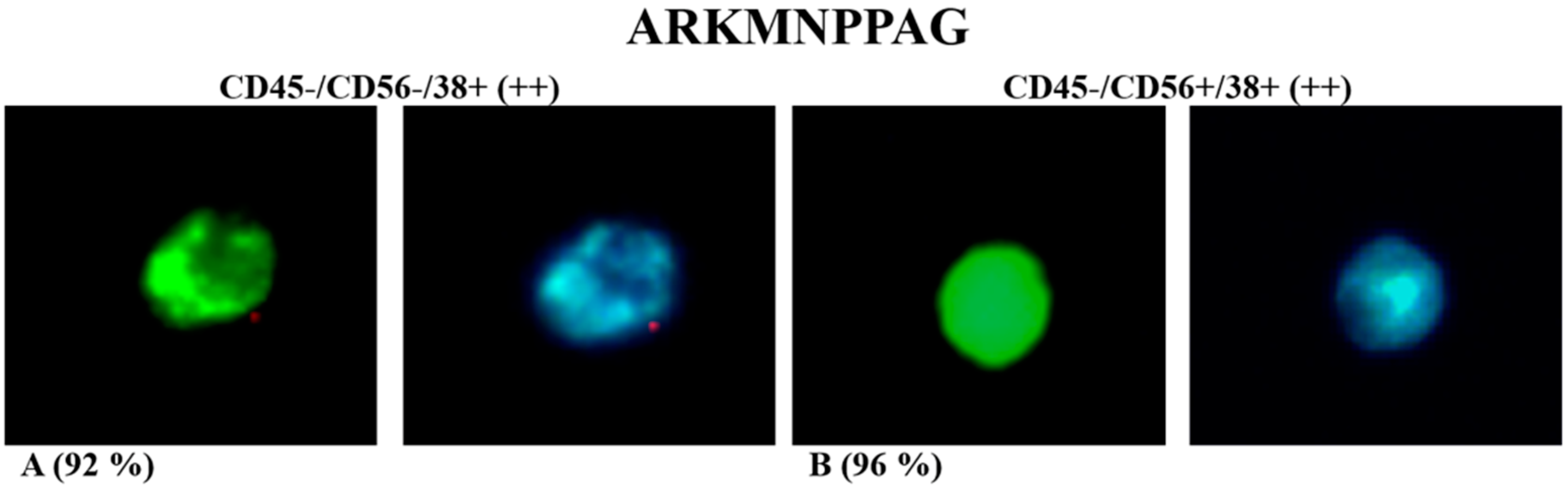
| ARKMNPPAG | + | +/− | + | − |
| RRANNPSPQ | − | nd | + | + |
| RRYTHPNNS | − | nd | + | + |
| KKAHPGGSP | + | nd | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Plano, L.M.; Franco, D.; Bonsignore, M.; Fazio, E.; Trusso, S.; Allegra, A.; Musolino, C.; Cavaliere, R.; Ferlazzo, G.; Neri, F.; et al. Phage-Phenotype Imaging of Myeloma Plasma Cells by Phage Display. Appl. Sci. 2021, 11, 7910. https://doi.org/10.3390/app11177910
De Plano LM, Franco D, Bonsignore M, Fazio E, Trusso S, Allegra A, Musolino C, Cavaliere R, Ferlazzo G, Neri F, et al. Phage-Phenotype Imaging of Myeloma Plasma Cells by Phage Display. Applied Sciences. 2021; 11(17):7910. https://doi.org/10.3390/app11177910
Chicago/Turabian StyleDe Plano, Laura M., Domenico Franco, Martina Bonsignore, Enza Fazio, Sebastiano Trusso, Alessandro Allegra, Caterina Musolino, Riccardo Cavaliere, Guido Ferlazzo, Fortunato Neri, and et al. 2021. "Phage-Phenotype Imaging of Myeloma Plasma Cells by Phage Display" Applied Sciences 11, no. 17: 7910. https://doi.org/10.3390/app11177910
APA StyleDe Plano, L. M., Franco, D., Bonsignore, M., Fazio, E., Trusso, S., Allegra, A., Musolino, C., Cavaliere, R., Ferlazzo, G., Neri, F., & Guglielmino, S. P. P. (2021). Phage-Phenotype Imaging of Myeloma Plasma Cells by Phage Display. Applied Sciences, 11(17), 7910. https://doi.org/10.3390/app11177910