
Artificial Neural Network (ANN) Modeling Analysis of Algal Blooms in an Estuary with Episodic and Anthropogenic Freshwater Inputs



Abstract
1. Introduction
2. Materials and Methods
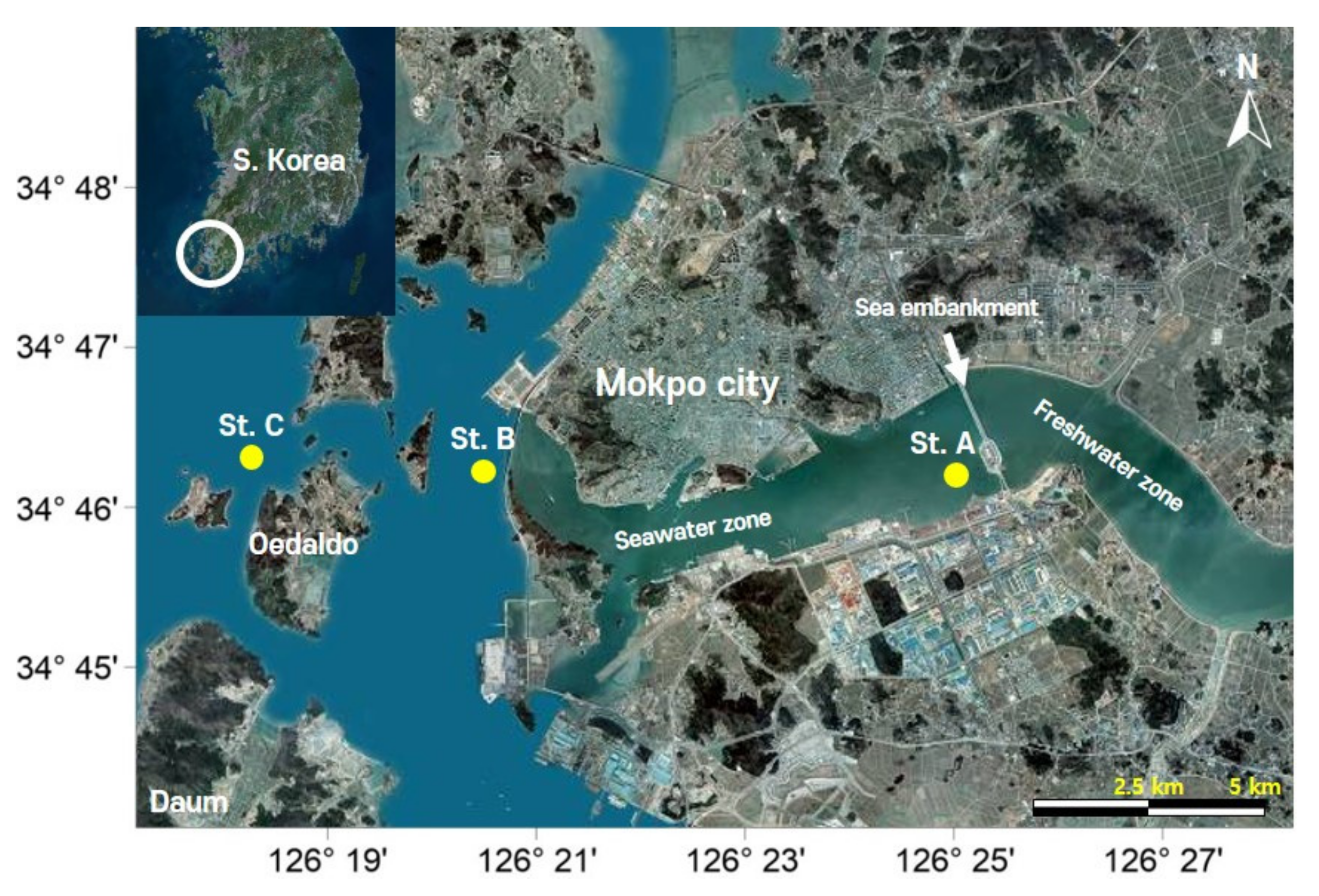
2.1. Sampling Site and Data Aquisition
2.2. Artificial Neural Network Model
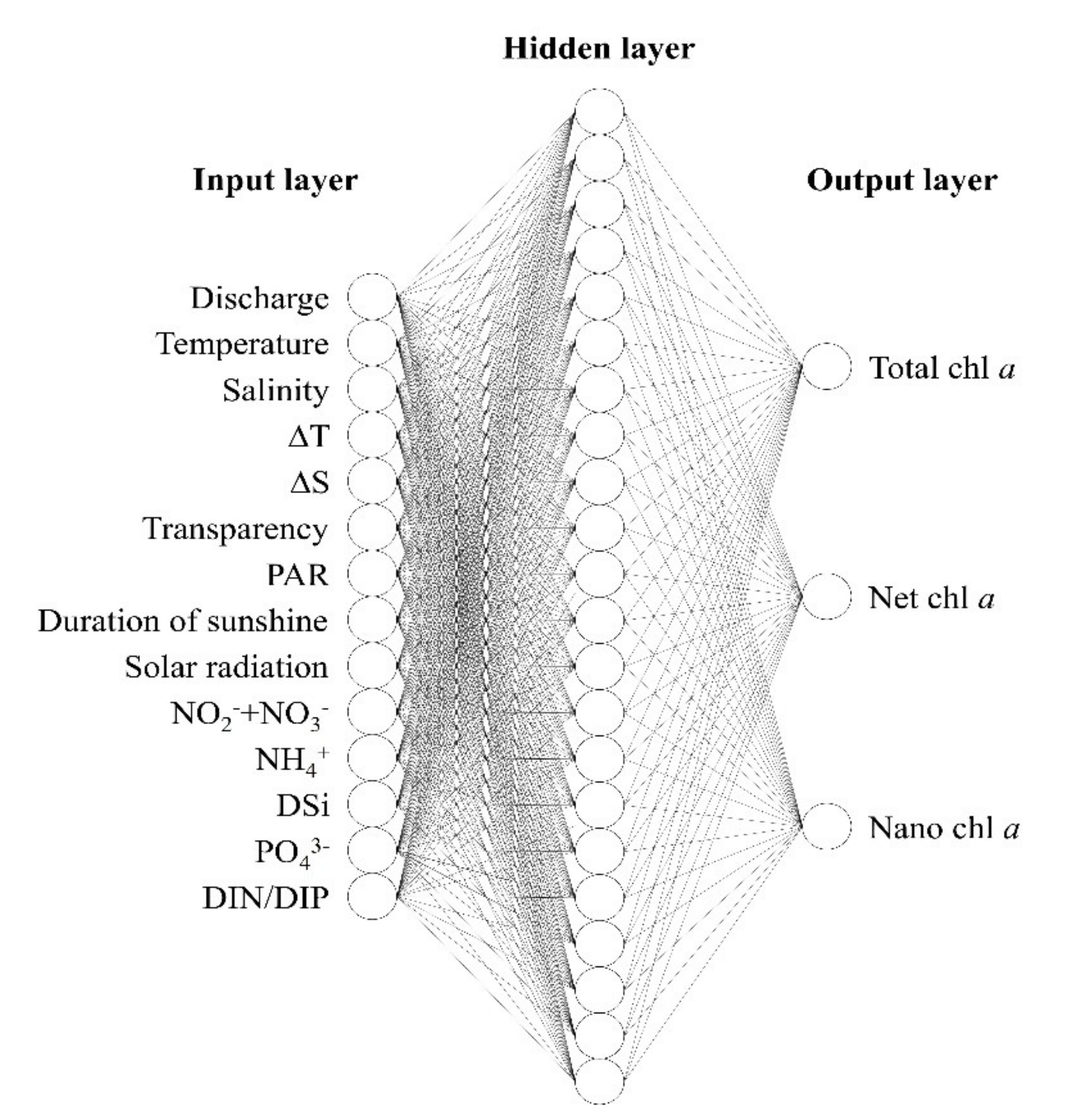
2.2.1. ANN Structure
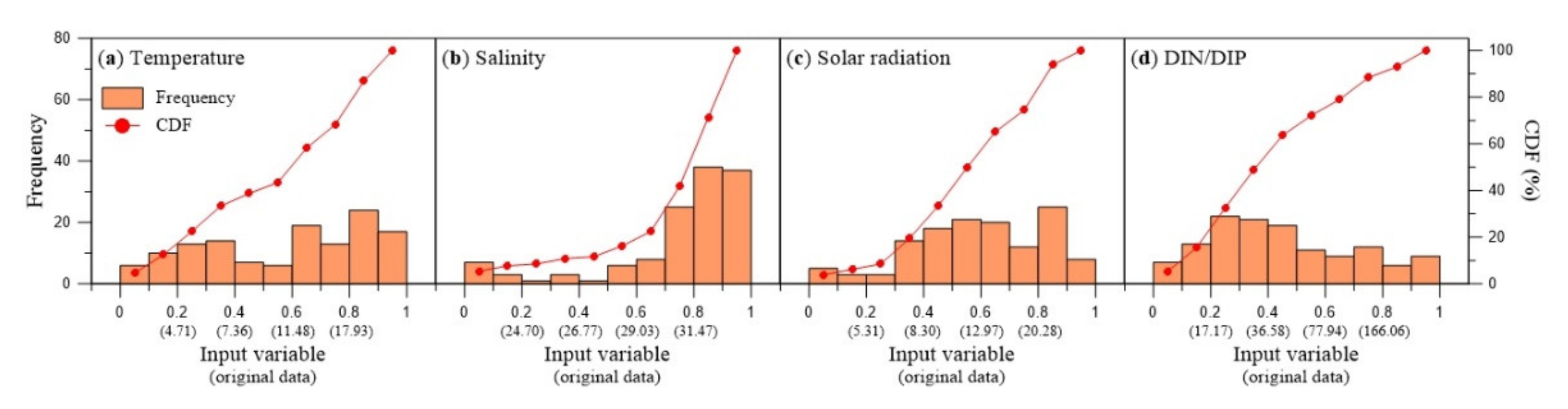
2.2.2. Learning Dataset and Normalization
- : 5% minimum value
- : 95% maximum value
- : original data
- : transformed data.
2.2.3. Statistical Validation
- : number of samples
- Xi: measurement
- Xim: mean of measurement
- Yi: output.
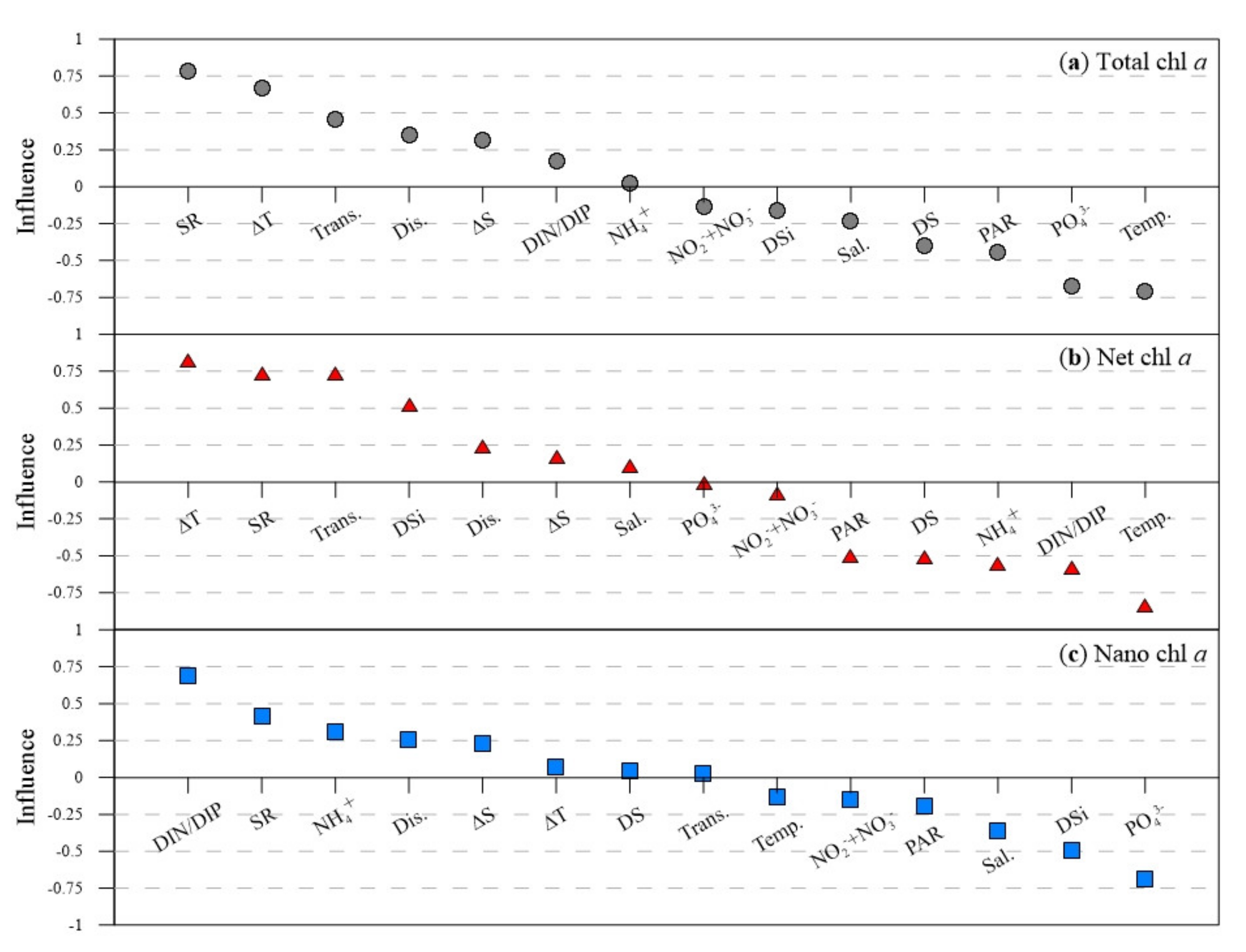
2.2.4. Environmental Impact Assessment
3. Results
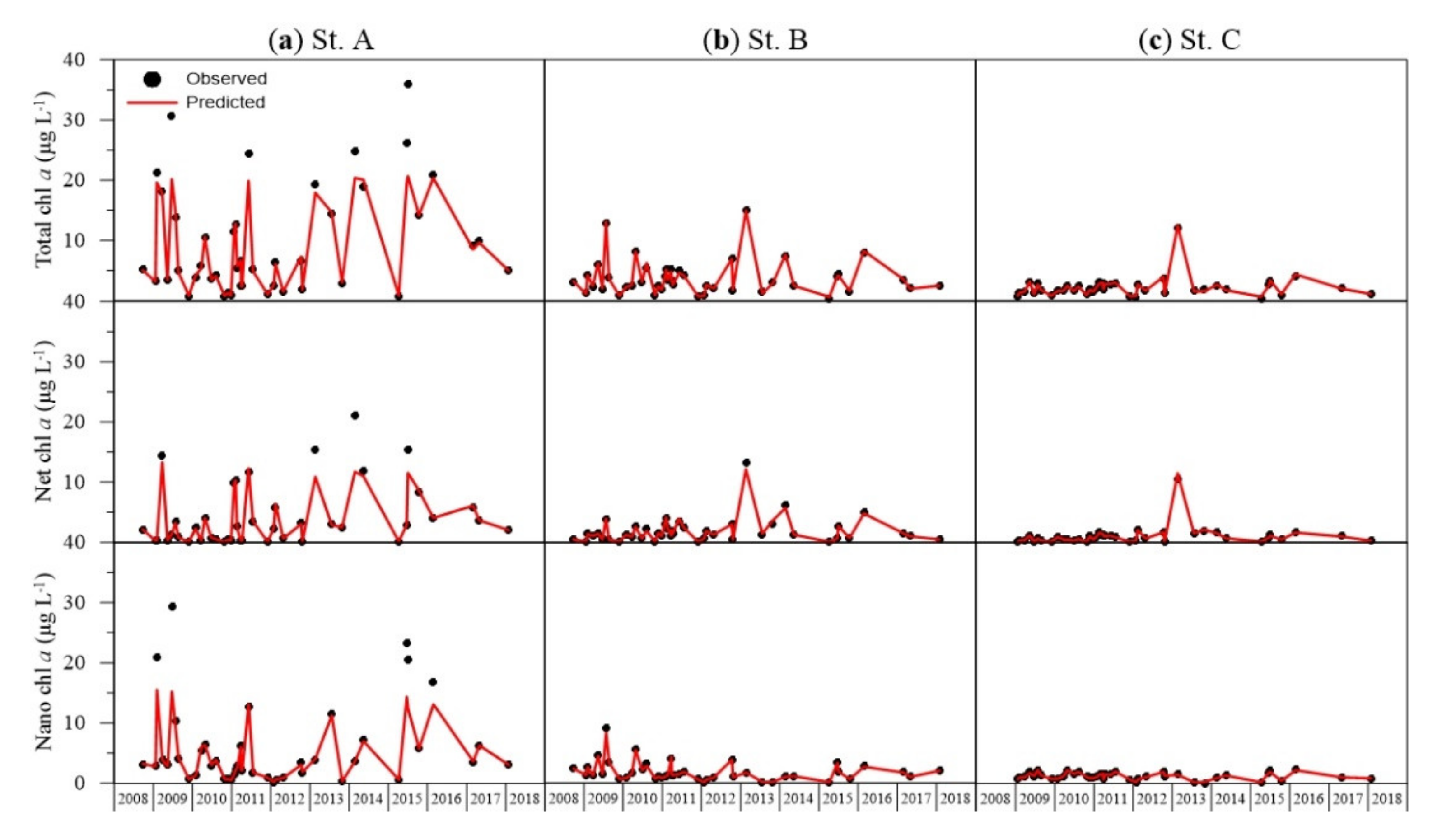
3.1. Validation of ANN Model
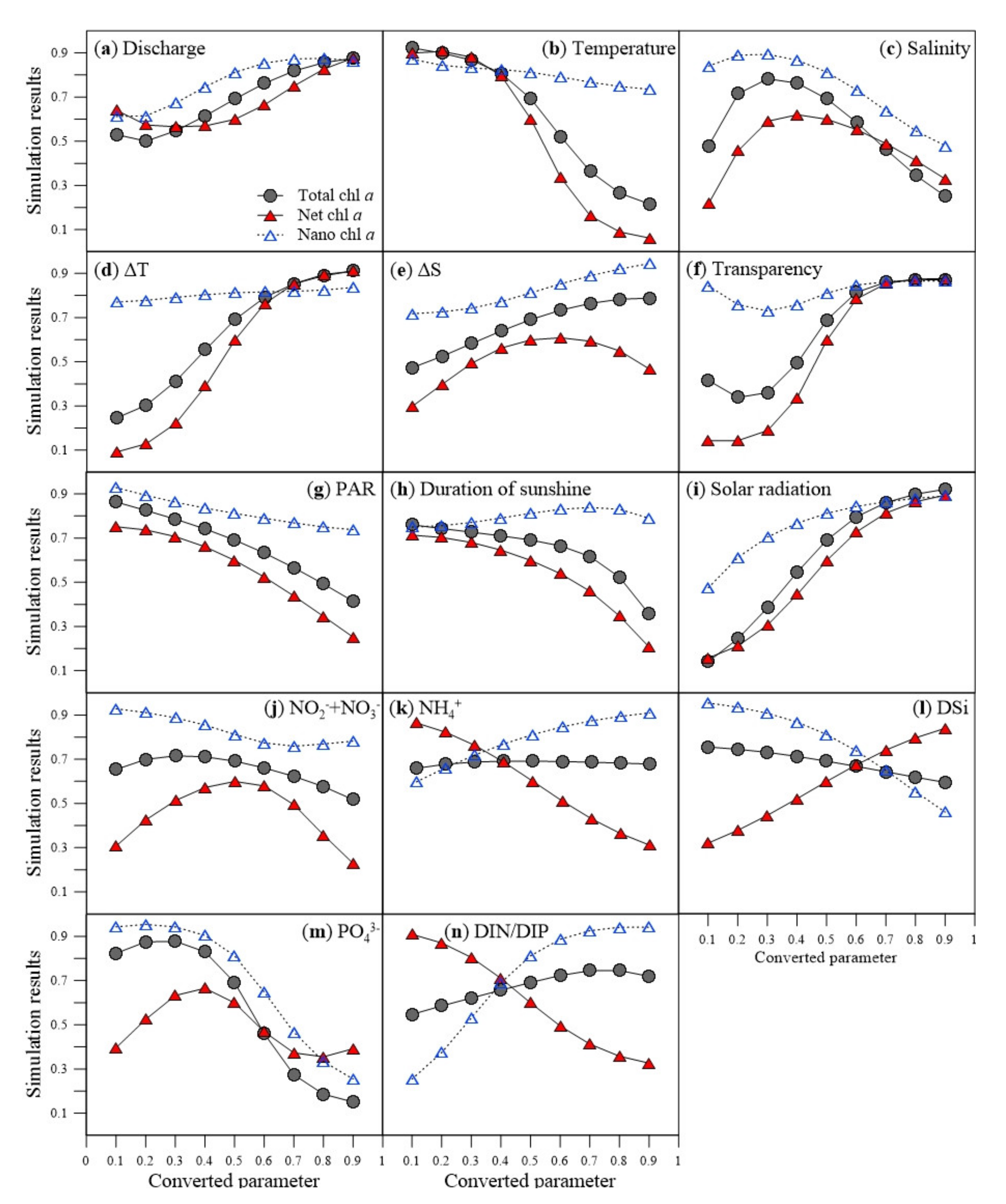
3.2. Relationship between Size-Fractionated Phytoplankton and Environmental Factors
4. Discussion
4.1. Applicability of the ANN Model to Algal Blooms in an Altered Estuary
4.2. Factors Influencing Variations in the Phytoplankton Size Structure
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kemp, M.W.; Boynton, W.R. Spatial and temporal coupling of nutrient inputs to estuarine primary production: The role of particulate transport and decomposition. Bull. Mar. Sci. 1984, 35, 522–535. [Google Scholar]
- Sundbäck, K.; Jönsson, B.; Nilsson, P.; Lindström, I. Impact of accumulating drifting macroalgae on a shallow-water sediment system: An experimental study. Mar. Ecol. Prog. Ser. 1990, 58, 261–274. Available online: https://www.jstor.org/stable/24842201 (accessed on 28 May 2021). [CrossRef]
- Landsberg, J.H. The effects of harmful algal blooms on aquatic organisms. Rev. Fish. Sci. 2002, 10, 113–390. [Google Scholar] [CrossRef]
- Harrison, P.J.; Piontkovski, S.; Al-Hashmi, K. Understanding how physical-biological coupling influences harmful algal blooms, low oxygen and fish kills in the Sea of Oman and the Western Arabian Sea. Mar. Pollut. Bull. 2017, 114, 25–34. [Google Scholar] [CrossRef]
- Rabalais, N.N.; Turner, R.E.; Wiseman Jr., W.J. Gulf of Mexico hypoxia, a.k.a. “The dead zone”. Annu. Rev. Ecol. Syst. 2002, 33, 235–263. [Google Scholar] [CrossRef]
- Finkel, Z.V.; Beardall, J.; Flynn, K.J.; Quiqq, A.; Rees, T.A.V.; Raven, J.A. Phytoplankton in a changing world: Cells size and elemental stoichiometry. J. Plankton Res. 2010, 32, 119–137. [Google Scholar] [CrossRef]
- Marañón, E. Cell size as a key determinant of phytoplankton metabolism and community structure. Annu. Rev. Mar. Sci. 2015, 7, 241–264. [Google Scholar] [CrossRef] [PubMed]
- Hansen, D.V.; Rattray, M. Gravitational circulation in straits and estuaries. J. Mar. Res. 1965, 23, 104–122. [Google Scholar]
- Ryther, J.H. Photosynthesis and fish production in the sea. Science 1969, 166, 72–76. Available online: https://www.jstor.org/stable/1727735 (accessed on 28 May 2021). [CrossRef]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Sin, Y.; Lee, H. Changes in hydrology, water, quality, and algal blooms in a freshwater system impounded with engineered structures in a temperate monsoon river estuary. J. Hydrol. Reg. Stud. 2020, 32, 100744. [Google Scholar] [CrossRef]
- Shin, Y.; Yoon, B. Change in taxonomic composition of phytoplankton and environmental factors after construction of Dike in Yeongsan River Estuary. Korean J. Environ. Biol. 2011, 29, 212–224. [Google Scholar]
- Song, E.; Cho, K.; Shin, Y. Exploring the dynamics of dissolved oxygen and vertical density structure of water column in the Youngsan Lake. J. Environ. Sci. Int. 2015, 24, 163–174. [Google Scholar] [CrossRef]
- Cho, Y.K.; Park, L.H.; Cho, C.; Lee, I.T.; Park, K.Y.; Oh, C.W. Multi-layer structure in the Youngsan estuary, Korea. Estuar. Coast. Shelf Sci. 2004, 61, 325–329. [Google Scholar] [CrossRef]
- Sin, Y.; Jeong, B. Short-term variations of phytoplankton communities in response to anthropogenic stressors in a highly altered temperate estuary. Estuar. Coast. Shelf Sci. 2015, 156, 1–9. [Google Scholar] [CrossRef]
- Sin, Y.; Yu, H. Phytoplankton community and surrounding water conditions in the Youngsan River estuary: Weekly variation in the saltwater zone. Ocean. Polar Res. 2018, 40, 191–202. [Google Scholar] [CrossRef]
- Amstrong, R.A. Grazing limitation and nutrient limitation in marine ecosystem: Steady state solution of an ecosystem model with multiple food chains. Limnol. Oceanogr. 1994, 39, 597–608. [Google Scholar] [CrossRef]
- Caraco, N.F.; Cole, J.J.; Raymond, P.A.; Strayer, D.L.; Pace, M.L.; Findlay, S.E.G.; Fischer, D.T. Zebra mussel invasion in a large, turbid river: Phytoplankton response to increased grazing. Ecology 1997, 78, 588–602. [Google Scholar] [CrossRef]
- Sin, Y.; Jeong, B.; Park, C. Semidiurnal dynamics of phytoplankton size structure and taxonomic composition in a macrotidal temperate estuary. Estuaries Coast. 2015, 38, 546–557. [Google Scholar] [CrossRef]
- Bharathi, M.D.; Sarma, V.V.S.S. Impact of monsoon-induced discharge on phytoplankton community structure in the tropical Indian estuaries. Reg. Stud. Mar. Sci. 2019, 31, 100795. [Google Scholar] [CrossRef]
- Karul, C.; Soyupak, S.; Cilesiz, A.F.; Akaby, N.; Germen, E. Case studies on the use of neural networks in eutrophication modeling. Ecol. Model. 2000, 134, 145–152. [Google Scholar] [CrossRef]
- Gevrey, M.; Dimopoulos, I.; Lek, S. Review and comparison of methods to study the contribution of variables in Artificial Neural Network models. Ecol. Model. 2003, 160, 249–264. [Google Scholar] [CrossRef]
- Taherdangkoo, R.; Taherdangkoo, M. Modified Stem Cells Algorithm-based Neural Network Applied to Bottom Hole Circulating Pressure in Underbalanced Drilling. Int. J. Pet. Eng. 2015, 1, 178–188. [Google Scholar] [CrossRef]
- Albawi, S.; Mohammed, T.A.; Al-Zawi, S. Understanding of a Convolutional Neural Network. ICET 2017. [Google Scholar] [CrossRef]
- Taherdangkoo, R.; Tatomir, A.; Taherdangkoo, M.; Qiu, P.; Sauter, M. Nonlinear Autoregressive Neural Networks to Predict Hydraulic Fracturing Fluid Leakage into Shallow Groundwater. Water 2020, 12, 841. [Google Scholar] [CrossRef]
- Mattei, F.; Franceschini, S.; Scardi, M. A depth-resolved artificial neural network model of marine phytoplankton primary production. Ecol. Model. 2018, 382, 51–62. [Google Scholar] [CrossRef]
- Joo, G.J.; Kim, H.G.; Hong, S.W.; Jeong, K.S.; Kim, D.K. Determination of sensitive variables regardless of hydrological alteration in artificial neural network model of chlorophyll a: Case study of Nakdong River. Ecol. Model. 2019, 398, 67–76. [Google Scholar] [CrossRef]
- Sin, Y.; Jeong, B. Anthropogenic disturbance of tidal variation in the water properties and phytoplankton community of an estuarine system. Estuar. Coast. 2020, 43, 547–559. [Google Scholar] [CrossRef]
- Lee, J.H.W.; Huang, Y.; Dickman, M.; Jayawardena, A.W. Neural network modelling of coastal algal blooms. Ecol. Model. 2003, 159, 179–201. [Google Scholar] [CrossRef]
- Jeffrey, S.W.; Hallegraeff, G.M. Phytoplankton ecology of Australian waters. In Biology of Marine Plants; Clayton, M.N., King, R.J., Eds.; Longman Cheshire: Melbourne, Australia, 1990; pp. 310–348. [Google Scholar]
- Tian, W.; Liao, Z.; Zhang, J. An optimization of artificial neural network model for predicting chlorophyll dynamics. Ecol. Model. 2017, 364, 42–52. [Google Scholar] [CrossRef]
- Domingues, R.B.; Guerra, C.C.; Barbosa, A.B.; Galvão, H.M. Are nutrients and light limiting summer phytoplankton in a temperate coastal lagoon? Aquat. Ecol. 2015, 49, 127–146. [Google Scholar] [CrossRef]
- González-García, C.; Forja, J.; González-Cabrera, M.C.; Jiménez, M.P.; Lubián, L.M. Annual variations of total and fractionated chlorophyll and phytoplankton groups in the Gulf of Cadiz. Sci. Total Environ. 2018, 613–614, 1551–1565. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis; Blackwell Science: Oxford, UK, 1997; p. 375. [Google Scholar]
- Taylor, G.T.; Muller-Karger, F.E.; Thunell, R.C.; Scranton, M.I.; Astor, Y.; Varela, R.; Ghinaglia, L.T.; Lorenzoni, L.; Fanning, K.A.; Hameed, S.; et al. Ecosystem responses in the southern Caribbean Sea to global climate change. Proc. Natl. Acad. Sci. USA 2012, 109, 19315–19320. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Oseji, O.; Mitra, M.; Waguespack, Y.; Chen, N. Phytoplankton pigments in Maryland Coastal Bay sediments as biomarkers of sources of organic matter to benthic community. J. Coast. Res. 2015, 32, 768–775. [Google Scholar] [CrossRef]
- Sin, Y.; Hyun, B.; Jeong, B.; Soh, H.Y. Impacts of eutrophic freshwater inputs on water quality and phytoplankton size structure in a temperate estuary altered by a sea dike. Mar. Environ. Res. 2013, 85, 54–63. [Google Scholar] [CrossRef]
- Goldman, J.C.; Ryther, J.H. Temperature-influenced species competition in mass cultures of marine phytoplankton. Biotechnol. Bioeng. 1976, 18, 1125–1144. [Google Scholar] [CrossRef]
- Sin, Y.; Lee, E.; Lee, Y.; Shin, K.H. The river-estuarine continuum of nutrients and phytoplankton communities in an estuary physically divided by a sea dike. Estuar. Coast. Shelf Sci. 2015, 163, 279–289. [Google Scholar] [CrossRef]
- Lehrter, J.C. Regulation of eutrophication susceptibility in oligohaline regions of a northern Gulf of Mexico estuary, Mobile Bay, Alabama. Mar. Pollut. Bull. 2008, 56, 1446–1460. [Google Scholar] [CrossRef] [PubMed]
- Adolf, J.E.; Yeager, C.L.; Miller, W.D.; Mallonee, M.E.; Harding, L.W., Jr. Environmental forcing of phytoplankton floral composition, biomass, and primary productivity in Chesapeake Bay, USA. Estuar. Coast. Shelf Sci. 2006, 67, 108–122. [Google Scholar] [CrossRef]
- Smith, V.H. Responses of estuarine and coastal marine phytoplankton to nitrogen and phosphorus enrichment. Limnol. Oceanogr. 2006, 51, 377–384. [Google Scholar] [CrossRef]
- Yoon, B.; Lee, E.; Kang, T.; Shin, Y. Long-term change of phytoplankton biomass (chlorophyll-a), environmental factors and freshwater discharge in Youngsan Estuary. Korean J. Ecol. Environ. 2013, 46, 205–214. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Input data | Parameter (unit) | Range | Mean ± SD |
| Discharge (×103 ton) | 1.00–52,621.00 | 7673.05 ± 13,301.20 | |
| Temperature (°C) | 2.78–26.54 | 13.05 ± 7.16 | |
| Salinity (psu) | 11.82–35.19 | 30.74 ± 3.63 | |
| ΔT (°C) | 0.45–7.69 | 2.56 ± 1.09 | |
| ΔS (psu) | 1.57–22.77 | 3.43 ± 3.25 | |
| Transparency (m) | 0.40–5.20 | 1.49 ± 0.73 | |
| PAR (µmol m−2 s−1) | 1.81–1630.00 | 395.62 ± 376.67 | |
| Duration of sunshine (h) | 0.10–12.20 | 6.43 ± 3.76 | |
| Solar radiation (MJ m−2) | 1.06–28.42 | 14.33 ± 6.79 | |
| NO2− + NO3− (µM) | 0.07–258.17 | 24.15 ± 40.35 | |
| NH4+ (µM) | 0.21–88.29 | 9.11 ± 11.30 | |
| DSi (µM) | 0.93–269.20 | 30.60 ± 33.43 | |
| PO43− (µM) | 0.02–31.23 | 1.24 ± 3.19 | |
| DIN/DIP | 0.54–1852.68 | 89.31 ± 185.31 | |
| Output data | Total chl a (µg L−1) | 0.18–35.84 | 5.25 ± 6.48 |
| Net chl a (µg L−1) | 0.02–21.02 | 2.34 ± 3.61 | |
| Nano chl a (µg L−1) | 0.02–29.22 | 2.90 ± 4.50 |
| Index | Total chl a | Net chl a | Nano chl a |
|---|---|---|---|
| SSE | 0.0003 | 0.0001 | 0.0002 |
| RMSE | 0.0173 | 0.0119 | 0.0138 |
| R2 | 0.9952 | 0.9976 | 0.9957 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.; Sin, Y. Artificial Neural Network (ANN) Modeling Analysis of Algal Blooms in an Estuary with Episodic and Anthropogenic Freshwater Inputs. Appl. Sci. 2021, 11, 6921. https://doi.org/10.3390/app11156921
Park S, Sin Y. Artificial Neural Network (ANN) Modeling Analysis of Algal Blooms in an Estuary with Episodic and Anthropogenic Freshwater Inputs. Applied Sciences. 2021; 11(15):6921. https://doi.org/10.3390/app11156921
Chicago/Turabian StylePark, Sangjun, and Yongsik Sin. 2021. "Artificial Neural Network (ANN) Modeling Analysis of Algal Blooms in an Estuary with Episodic and Anthropogenic Freshwater Inputs" Applied Sciences 11, no. 15: 6921. https://doi.org/10.3390/app11156921
APA StylePark, S., & Sin, Y. (2021). Artificial Neural Network (ANN) Modeling Analysis of Algal Blooms in an Estuary with Episodic and Anthropogenic Freshwater Inputs. Applied Sciences, 11(15), 6921. https://doi.org/10.3390/app11156921