
Automatic Actin Filament Quantification and Cell Shape Modeling of Osteoblasts on Charged Ti Surfaces

 , ,
, ,  ,
,  and
and 
Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Applied Titanium Arrays and Their Properties
2.2. Cell Biological Investigations
2.2.1. Plasma Membrane Staining and Cell Area Quantification
2.2.2. Scanning Electron Microscopy (SEM)
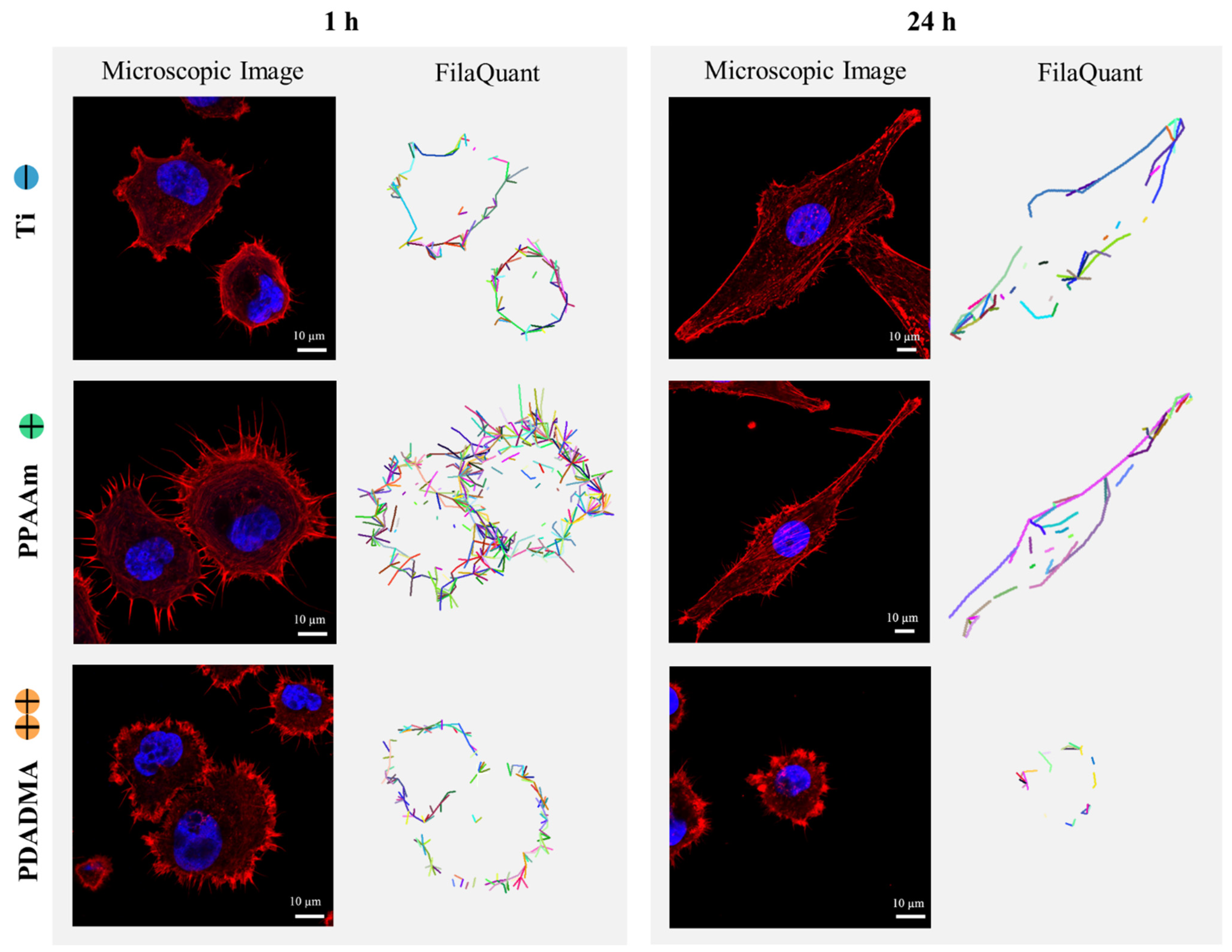
2.2.3. Actin Staining and FilaQuant Software
2.2.4. Edge Height Measurement via Scanning Ion Conductance Microscopy
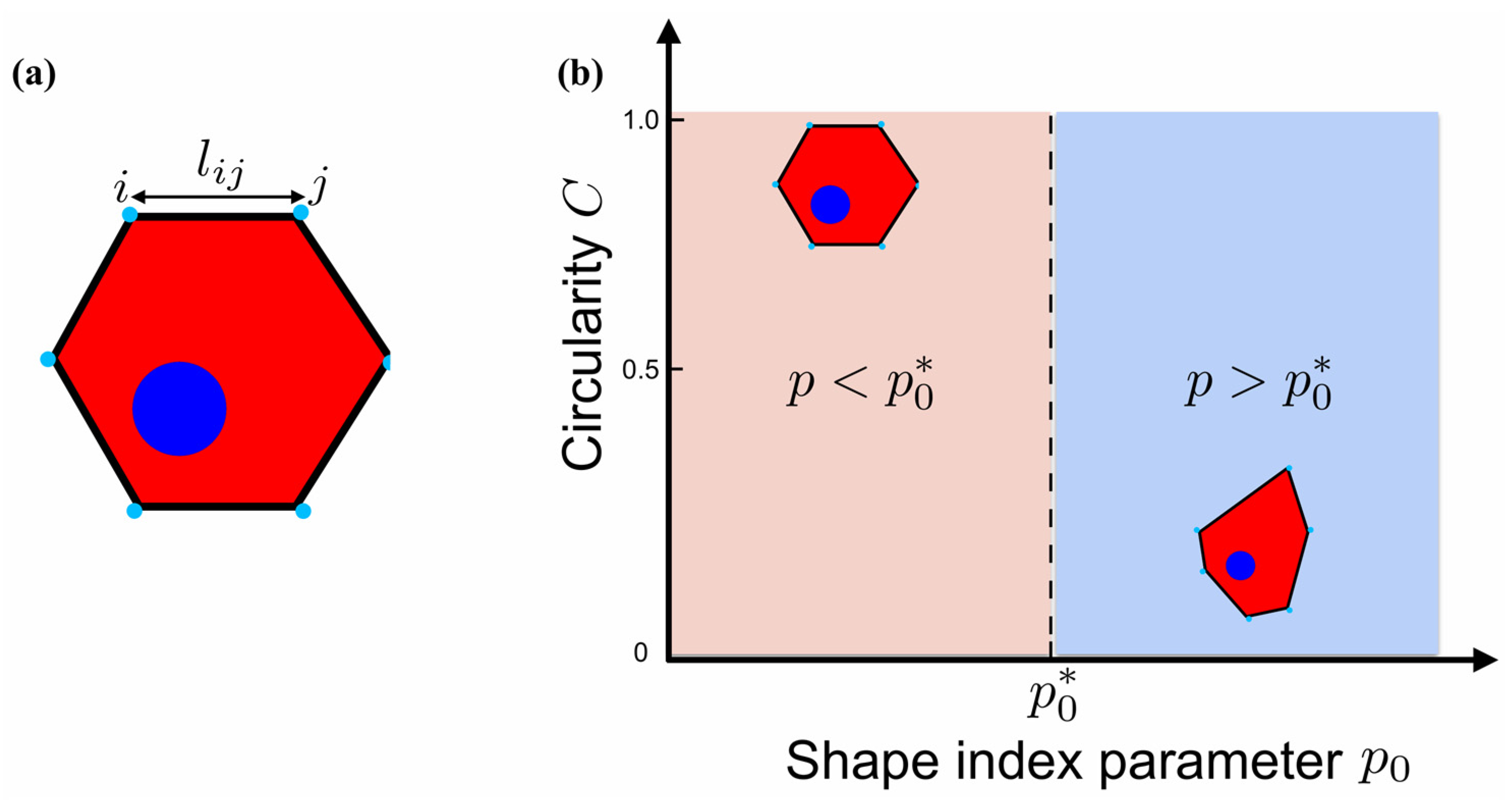
2.3. Mathematical Model for the Description of Cell Shape Changes
2.4. Statistics
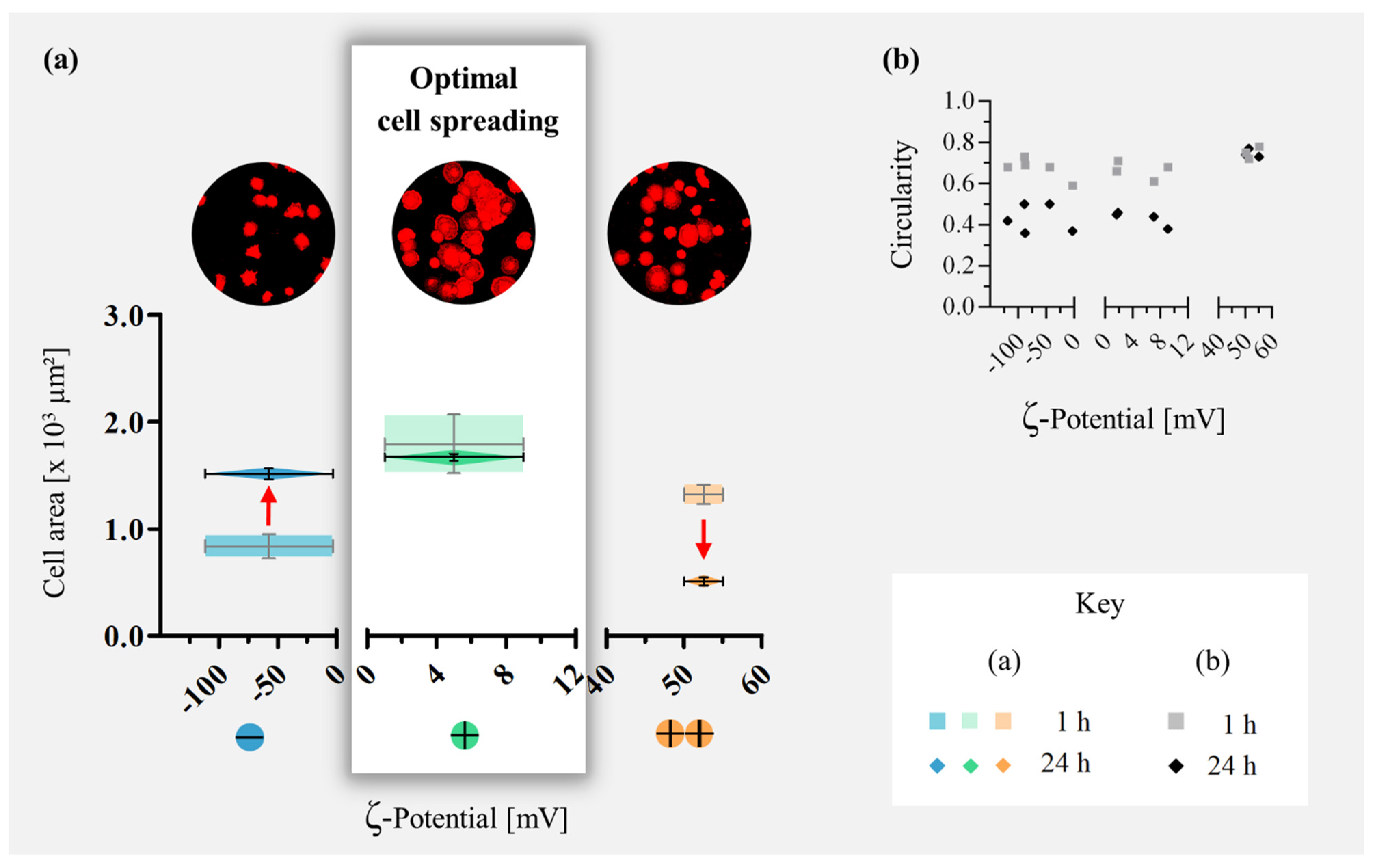
3. Results
3.1. Actin Cytoskeleton and Quantification of Filaments
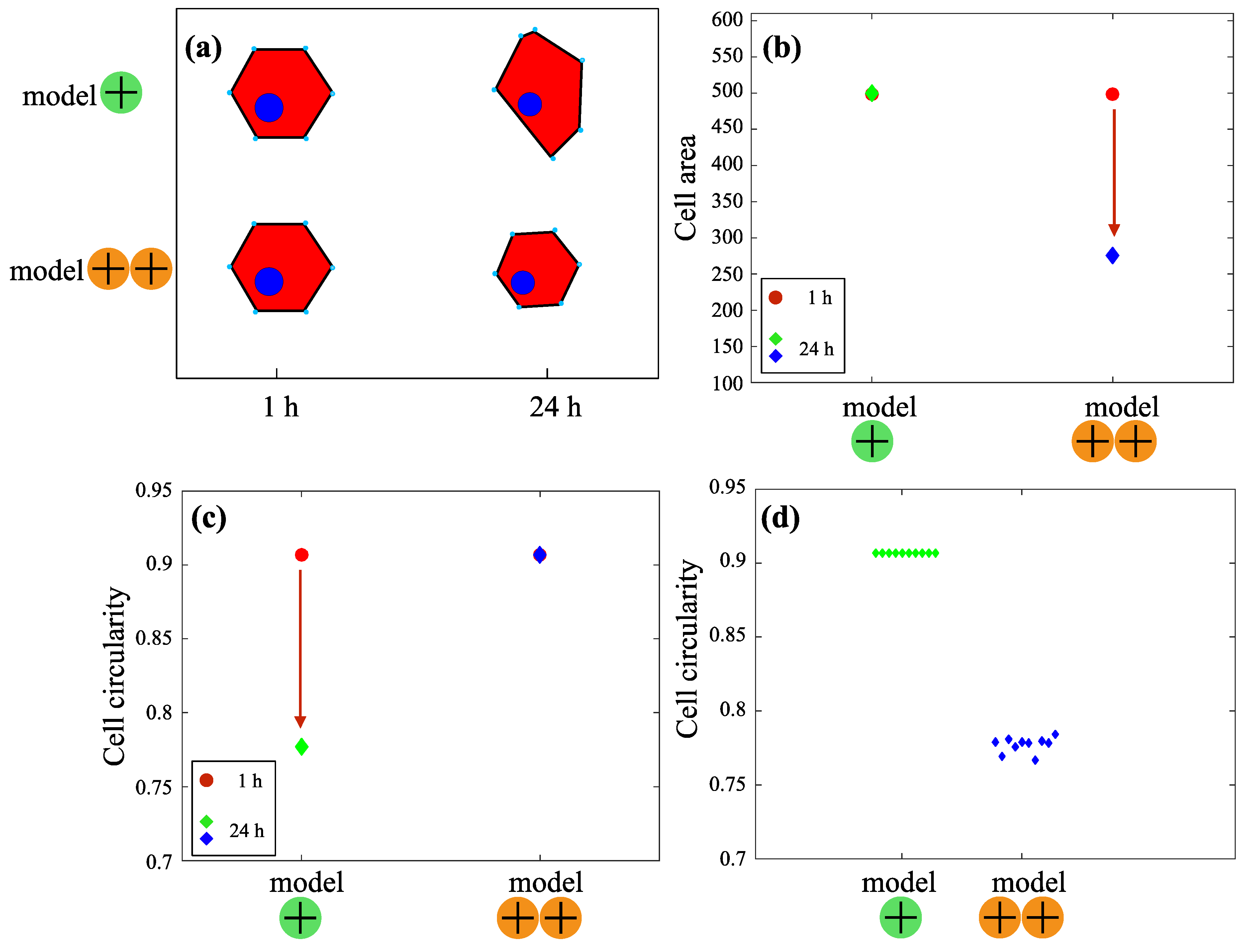
3.2. Mathematical Modeling of the Cell Shape
3.2.1. Model +: Cell Morphology on the Substrate with Moderately Positive Surface Charge
3.2.2. Model ++: Cell Morphology on a Substrate with Highly Positive Surface Charge
3.3. Edge Height of Cells on Moderately Positively Charged PPAAm
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kokubo, T.; Yamaguchi, S. Growth of Novel Ceramic Layers on Metals via Chemical and Heat Treatments for Inducing Various Biological Functions. Front. Bioeng. Biotechnol. 2015, 3, 176. [Google Scholar] [CrossRef] [PubMed]
- Muderrisoglu, C.; Saveleva, M.; Abalymov, A.; Van der Meeren, L.; Ivanova, A.; Atkin, V.; Parakhonskiy, B.; Skirtach, A.G. Nanostructured Biointerfaces Based on Bioceramic Calcium Carbonate/Hydrogel Coatings on Titanium with an Active Enzyme for Stimulating Osteoblasts Growth. Adv. Mater. Interfaces 2018, 5, 1800452. [Google Scholar] [CrossRef]
- Devgan, S.; Sidhu, S.S. Evolution of surface modification trends in bone related biomaterials: A review. Mater. Chem. Phys. 2019, 233, 68–78. [Google Scholar] [CrossRef]
- Wennerberg, A.; Albrektsson, T. Effects of titanium surface topography on bone integration: A systematic review. Clin. Oral. Implants Res. 2009, 20, 172–184. [Google Scholar] [CrossRef] [PubMed]
- Nikkhah, M.; Edalat, F.; Manoucheri, S.; Khademhosseini, A. Engineering microscale topographies to control the cell–substrate interface. Biomaterials 2012, 33, 5230–5246. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.S.; KS, S.K.; Grandhi, V.V.; Gupta, V. The Effects of Titanium Implant Surface Topography on Osseointegration: Literature Review. JMIR Biomed. Eng. 2019, 4, e13237. [Google Scholar] [CrossRef]
- Felgueiras, H.P.; Antunes, J.C.; Martins, M.C.L.; Barbosa, M.A. Fundamentals of protein and cell interactions in biomaterials. Pept. Proteins Biomater. Tissue Regen. Repair 2018, 1–27. [Google Scholar] [CrossRef]
- Ferrari, M.; Cirisano, F.; Morán, M.C. Mammalian Cell Behavior on Hydrophobic Substrates: Influence of Surface Properties. Colloids Interfaces 2019, 3, 48. [Google Scholar] [CrossRef]
- Chen, S.; Guo, Y.; Liu, R.; Wu, S.; Fang, J.; Huang, B.; Li, Z.; Chen, Z.; Chen, Z. Tuning surface properties of bone biomaterials to manipulate osteoblastic cell adhesion and the signaling pathways for the enhancement of early osseointegration. Colloid Surface B 2018, 164, 58–69. [Google Scholar] [CrossRef]
- Ekblom, P.; Timpl, R. Cell-to-cell contact and extracellular matrix. A multifaceted approach emerging. Curr. Opin. Cell Biol. 1996, 8, 599–601. [Google Scholar] [CrossRef]
- Gao, C.; Peng, S.; Feng, P.; Shuai, C. Bone biomaterials and interactions with stem cells. Bone Res. 2017, 5, 17059. [Google Scholar] [CrossRef]
- Agarwal, R.; García, A.J. Biomaterial strategies for engineering implants for enhanced osseointegration and bone repair. Adv. Drug Deliv. Rev. 2015, 94, 53–62. [Google Scholar] [CrossRef]
- Tzoneva, R.; Groth, T.; Altankov, G. Remodeling of fibrinogen by endothelial cells in dependence on fibronectin matrix assembly. Effect of substratum wettability. J. Mater. Sci. Mater. Med. 2002, 13, 1235–1244. [Google Scholar] [CrossRef] [PubMed]
- Cohen, M.; Joester, D.; Geiger, B.; Addadi, L. Spatial and Temporal Sequence of Events in Cell Adhesion: From Molecular Recognition to Focal Adhesion Assembly. ChemBioChem 2004, 5, 1393–1399. [Google Scholar] [CrossRef]
- Rebl, H.; Finke, B.; Schmidt, J.; Mohamad, H.S.; Ihrke, R.; Helm, C.A.; Nebe, J.B. Accelerated cell-surface interlocking on plasma polymer-modified porous ceramics. Mater. Sci. Engin. C 2016, 69, 1116–1124. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.M.; Johnstone, B.; Jacobson, K.; Caterson, B. The dynamic structure of the pericellular matrix on living cells. J. Cell Biol. 1993, 123, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Ercan, B.; Webster, T.J. Greater osteoblast proliferation on anodized nanotubular titanium upon electrical stimulation. Int. J. Nanomed. 2008, 3, 477–485. [Google Scholar] [CrossRef]
- Hunckler, J.; de Mel, A. A current affair: Electrotherapy in wound healing. J. Multidiscip. Healthc. 2017, 10, 179–194. [Google Scholar] [CrossRef]
- Glaser, R. Biophysics: An Introduction, 2nd ed.; Springer: Berlin, Germany, 2012; ISBN 978-366249596-4. [Google Scholar]
- Holler, F.J.; Skoog, D.A.; Crouch, S.R. Principles of Instrumental Analysis, 6th ed.; Thomson Brooks/Cole; Belmont, C.A., Ed.; Cengage Learning: Boston, MA, USA, 2007; p. 9. ISBN 978-0-495-01201-6. [Google Scholar]
- Pawlikowski, M. Electric Phenomenon in Bones as a Result of Piezoelectricity of Hydroxyapatite. Arch. Clin. Biomed. Res. 2017, 1, 132–139. [Google Scholar] [CrossRef]
- Anderson, J.C.; Eriksson, C. Electrical properties of wet collagen. Nature 1968, 21, 166–168. [Google Scholar] [CrossRef]
- Minary-Jolandan, M.; Yu, M.F. Nanoscale characterization of isolated individual type I collagen fibrils: Polarization and piezoelectricity. Nanotechnology 2009, 20, 085706. [Google Scholar] [CrossRef]
- Zheng, T.; Huang, Y.; Zhang, X.; Cai, Q.; Deng, X.; Yang, X. Mimicking the electrophysiological microenvironment of bone tissue using electroactive materials to promote its regeneration. J. Mater. Chem. B 2020, 8, 10221. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hou, Z.; Qin, Q.-H.; Fu, D.; Pan, S. Variation of Streaming Potentials with Time under Steady Fluid Pressure. Bone Appl. Sci. 2019, 9, 3726. [Google Scholar] [CrossRef]
- Balmer, T.W.; Vesztergom, S.; Broekmann, P.; Stahel, A.; Büchler, P. Characterization of the electrical conductivity of bone and its correlation to osseous structure. Sci. Rep. 2018, 8, 8601. [Google Scholar] [CrossRef]
- Hammond, C. Ionic gradients, membrane potential and ionic currents. In Cellular and Molecular Neurophysiology, 4th ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2015; pp. 39–54. [Google Scholar] [CrossRef]
- Meyer-Plath, A.A.; Schroeder, K.; Finke, B.; Ohl, A. Current trends in biomaterial surface functionalization—Nitrogen-containing plasma assisted processes with enhanced selectivity. Vacuum 2003, 71, 391–406. [Google Scholar] [CrossRef]
- Nebe, B.; Finke, B.; Luethen, F.; Bergemann, C.; Schroeder, K.; Rychly, J.; Liefeith, K.; Ohl, A. Improved initial osteoblast functions on amino-functionalized titanium surfaces. Biomol. Eng. 2007, 24, 447–454. [Google Scholar] [CrossRef]
- von der Mark, K.; Park, J. Engineering biocompatible implant surfaces: Part II: Cellular recognition of biomaterial surfaces: Lessons from cell-matrix interactions. Prog. Mater. Sci. 2013, 58, 327–381. [Google Scholar] [CrossRef]
- Khalili, A.; Ahmad, M. A Review of Cell Adhesion Studies for Biomedical and Biological Applications. Int. J. Mol. Sci. 2015, 16, 18149–18184. [Google Scholar] [CrossRef]
- Yang, S.; Kim, H.-M. The RhoA-ROCK-PTEN pathway as a molecular switch for anchorage dependent cell behavior. Biomaterials 2012, 33, 2902–2915. [Google Scholar] [CrossRef] [PubMed]
- Gruening, M.; Neuber, S.; Fricke, K.; Helm, C.A.; Nebe, B. Cell-Material Interaction - Spreading Course Correlates with Surface Charge. Am. J. Biomed. Sci. Res. 2020, 9, 4–6. [Google Scholar] [CrossRef]
- Gruening, M.; Neuber, S.; Nestler, P.; Lehnfeld, J.; Dubs, M.; Fricke, K.; Schnabelrauch, M.; Helm, C.A.; Müller, R.; Staehlke, S.; et al. Enhancement of Intracellular Calcium Ion Mobilization by Moderately but Not Highly Positive Material Surface Charges. Front. Bioeng. Biotechnol. 2020, 8, 1016. [Google Scholar] [CrossRef]
- Ingber, D.E. Tensegrity II. How structural networks influence cellular information processing networks. J. Cell Sci. 2003, 116, 1397–1408. [Google Scholar] [CrossRef] [PubMed]
- Brakebusch, C.; Fässler, R. The integrin-actin connection, an eternal love affair. EMBO J. 2003, 22, 2324–2333. [Google Scholar] [CrossRef]
- Stricker, J.; Falzone, T.; Gardel, M.L. Mechanics of the F-actin cytoskeleton. J. Biomech. 2010, 43, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Mooney, D.J.; Langer, R.; Ingber, D.E. Cytoskeletal filament assembly and the control of cell spreading and function by extracellular matrix. J. Cell Sci. 1995, 108, 2311–2320. [Google Scholar] [CrossRef]
- Kharitonova, M.A.; Vasiliev, J.M.; Rovensky, Y.A.; Vasiliev, J.M. Length control is determined by the pattern of cytoskeleton. J. Cell Sci. 2004, 117, 1955–1960. [Google Scholar] [CrossRef] [PubMed]
- Matschegewski, C.; Staehlke, S.; Birkholz, H.; Lange, R.; Beck, U.; Engel, K.; Nebe, J.B. Automatic actin filament quantification of osteoblasts and their morphometric analysis on microtextured silicon-titanium arrays. Materials 2012, 5, 1176–1195. [Google Scholar] [CrossRef]
- Lüthen, F.; Lange, R.; Becker, P.; Rychly, J.; Beck, U.; Nebe, J.B. The influence of surface roughness of titanium on β1- and β3-integrin adhesion and organization of fibronectin in human osteoblastic cells. Biomaterials 2005, 26, 2423–2440. [Google Scholar] [CrossRef] [PubMed]
- Finke, B.; Luethen, F.; Schroeder, K.; Mueller, P.D.; Bergemann, C.; Frant, M.; Ohl, A.; Nebe, B.J. The effect of positively charged plasma polymerization on initial osteoblastic focal adhesion on titanium surfaces. Biomaterials 2007, 28, 4521–4534. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, H.S.; Neuber, S.; Helm, C.A. Surface Forces of Asymmetrically Grown Polyelectrolyte Multilayers: Searching for the Charges. Langmuir 2019, 35, 15491–15499. [Google Scholar] [CrossRef] [PubMed]
- Staehlke, S.; Rebl, H.; Nebe, B. Phenotypic stability of the human MG-63 osteoblastic cell line at different passages. Cell Biol. Int. 2019, 43, 22–32. [Google Scholar] [CrossRef]
- Matschegewski, C.; Staehlke, S.; Loeffler, R.; Lange, R.; Chai, F.; Kern, D.P.; Beck, U.; Nebe, B.J. Cell architecture–cell function dependencies on titanium arrays with regular geometry. Biomaterials 2010, 31, 5729–5740. [Google Scholar] [CrossRef] [PubMed]
- Birkholz, H. Extracting the Ridge Set as a Graph for Quantification of Actin Filament Images Obtained by Confocal Laser Scanning Microscopy; Pinoli, J.-C., Debayle, J., Gavet, Y., Gruy, F., Lambert, C., Eds.; International Society for Optics and Photonics: Bellingham, WA, USA, 2011; p. 800006. [Google Scholar] [CrossRef]
- Voelkner, C.; Wendt, M.; Lange, R.; Ulbrich, M.; Gruening, M.; Staehlke, S.; Nebe, B.; Barke, I.; Speller, S. The nanomorphology of cell surfaces of adhered osteoblasts. Beilstein J. Nanotechnol. 2021, 12, 242–256. [Google Scholar] [CrossRef] [PubMed]
- Alt, S.; Ganguly, P.; Salbreux, G. Vertex models: From cell mechanics to tissue morphogenesis. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20150520. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Merkel, M.; Clark, M.J.; Jacob, A.E.; Dawson, J.E.; Manning, M.L.; Amack, J.D. Cell volume changes contribute to epithelial morphogenesis in zebrafish Kupffer’s vesicle. eLife 2018, 7, e30963. [Google Scholar] [CrossRef] [PubMed]
- Koride, S.; Loza, A.J.; Sun, S.X. Epithelial vertex models with active biochemical regulation of contractility can explain organized collective cell motility. APL Bioeng. 2018, 2, 031906. [Google Scholar] [CrossRef]
- Farhadifar, R.; Röper, J.-C.; Aigouy, B.; Eaton, S.; Jülicher, F. The influence of cell mechanics, cell-cell interactions, and proliferation on epithelial packing. Curr. Biol. 2007, 17, 2095–2104. [Google Scholar] [CrossRef]
- Kneuper, I.; Teale, W.; Dawson, J.E.; Tsugeki, R.; Katifori, E.; Palme, K.; Ditengou, F.A. Auxin biosynthesis and cellular efflux act together to regulate leaf vein patterning. J. Exp. Bot. 2021, 72, 1151–1165. [Google Scholar] [CrossRef]
- Bi, D.; Lopez, J.H.; Schwarz, J.M.; Manning, M.L. A density-independent rigidity transition in biological tissues. Nat. Phys. 2015, 11, 1074–1079. [Google Scholar] [CrossRef]
- Merkel, M.; Manning, M.L. A geometrically controlled rigidity transition in a model for confluent 3D tissues. New J. Phys. 2018, 20, 022002. [Google Scholar] [CrossRef]
- Staple, D.B.; Farhadifar, R.; Röper, J.-C.; Aigouy, B.; Eaton, S.; Jülicher, F. Mechanics and remodelling of cell packings in epithelia. Eur. Phys. J. E 2010, 33, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Raucher, D.; Sheetz, M.P. Cell spreading and lamellipodial extension rate is regulated by membrane tension. J. Cell Biol. 2000, 148, 127–136. [Google Scholar] [CrossRef]
- Mattila, P.K.; Lappalainen, P. Filopodia: Molecular architecture and cellular functions. Nat. Rev. Mol. Cell Biol. 2008, 9, 446–454. [Google Scholar] [CrossRef]
- Neoh, K.G.; Hu, X.; Zheng, D.; Kang, E.T. Balancing osteoblast functions and bacterial adhesion on functionalized titanium surfaces. Biomaterials 2012, 33, 2813–2822. [Google Scholar] [CrossRef] [PubMed]
- Small, J.V.; Rottner, K.; Kaverina, I. Functional design in the actin cytoskeleton. Curr. Opin. Cell Biol. 1999, 11, 54–60. [Google Scholar] [CrossRef]
- Small, J.V.; Stradal, T.; Vignal, E.; Rottner, K. The lamellipodium: Where motility begins. Trends Cell Biol. 2002, 12, 112–120. [Google Scholar] [CrossRef]
- Faix, J.; Breitsprecher, D.; Stradal, T.E.B.; Rottner, K. Filopodia: Complex models for simple rods. Int. J. Biochem. Cell Biol. 2009, 41, 1656–1664. [Google Scholar] [CrossRef]
- Partridge, M.A.; Marcantonio, E.E. Initiation of attachment and generation of mature focal adhesions by integrin-containing filopodia in cell spreading. Mol. Biol. Cell 2006, 17, 4237–4248. [Google Scholar] [CrossRef]
- Yamada, K.M.; Miyamoto, S. Integrin transmembrane signaling and cytoskeletal control. Curr. Opin. Cell Biol. 1995, 7, 681–689. [Google Scholar] [CrossRef]
- Clark, E.A.; Brugge, J.S. Integrins and signal transduction pathways: The road taken. Science 1995, 268, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Pollard, T.D.; Cooper, J.A. Actin, a central player in cell shape and movement. Science 2009, 326, 1208–1212. [Google Scholar] [CrossRef] [PubMed]
- Nebe, J.B.; Rebl, H.; Schlosser, M.; Staehlke, S.; Gruening, M.; Weltmann, K.-D.; Walschus, U.; Finke, B. Plasma Polymerized Allylamine—The Unique Cell-Attractive Nanolayer for Dental Implant Materials. Polymers 2019, 11, 1004. [Google Scholar] [CrossRef]
- Kunz, F.; Rebl, H.; Quade, A.; Matschegewski, C.; Finke, B.; Nebe, J. Osteoblasts with impaired spreading capacity benefit from the positive charges of plasma polymerised allylamine. Eur. Cells Mater. 2015, 29, 177–189. [Google Scholar] [CrossRef] [PubMed]
- Eisenbarth, E.; Linez, P.; Biehl, V.; Velten, D.; Breme, J.; Hildebrand, H. Cell orientation and cytoskeleton organisation on ground titanium surfaces. Biomol. Eng. 2002, 19, 233–237. [Google Scholar] [CrossRef]
- Mörke, C.; Rebl, H.; Finke, B.; Dubs, M.; Nestler, P.; Airoudj, A.; Roucoules, V.; Schnabelrauch, M.; Körtge, A.; Anselme, K.; et al. Abrogated Cell Contact Guidance on Amino-Functionalized Microgrooves. ACS Appl. Mater. Interfaces 2017, 9, 10461–10471. [Google Scholar] [CrossRef]
- Gregor, I.; Chizhik, A.; Karedla, N.; Enderlein, J. Metal-induced energy transfer. Nanophotonics 2019, 8, 1689–1699. [Google Scholar] [CrossRef]
- Kanchanawong, P.; Shtengel, G.; Pasapera, A.M.; Ramko, E.B.; Davidson, M.W.; Hess, H.F.; Waterman, C.M. Nanoscale architecture of integrin-based cell adhesions. Nature 2010, 468, 580–584. [Google Scholar] [CrossRef]
- Bekard, I.; Dunstan, D.E. Electric field induced changes in protein conformation. Soft Matter 2014, 10, 431–437. [Google Scholar] [CrossRef]
- Mecke, A.; Majoros, I.J.; Patri, A.K.; Baker, J.R.; Banaszak Holl, M.M.; Orr, B.G. Lipid Bilayer Disruption by Polycationic Polymers: The Roles of Size and Chemical Functional Group. Langmuir 2005, 21, 10348–10354. [Google Scholar] [CrossRef] [PubMed]
- Moerke, C.; Mueller, P.; Nebe, B. Attempted caveolae-mediated phagocytosis of surface-fixed micro-pillars by human osteoblasts. Biomaterials 2016, 76, 102–114. [Google Scholar] [CrossRef]
- Fröhlich, E. The role of surface charge in cellular uptake and cytotoxicity of medical nanoparticles. Int. J. Nanomed. 2012, 7, 5577. [Google Scholar] [CrossRef]
- Boudreau, N.J.; Jones, P.L. Extracellular matrix and integrin signalling: The shape of things to come. Biochem. J. 1999, 339, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Hall, A. Rho GTPases and the actin cytoskeleton. Science 1998, 279, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Moerke, C.; Mueller, P.; Nebe, J.B. Sensing of micropillars by osteoblasts involves complex intracellular signaling. J. Mater. Sci. Mater. Med. 2017, 28, 171. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model Quantities | Meaning |
|---|---|
| Dimensionless area of the cell | |
| Dimensionless preferred area of the cell | |
| Dimensionless perimeter of the cell | |
| Dimensionless preferred perimeter of the cell | |
| Dimensionless strength of resistance to area changes |
| 1 h | Ti | PPAAm | PDADMA |
| Filament number | 62.3 ± 5.1 b | 148.3 ± 10.7 a | 51.4 ± 6.3 b |
| Total filament length [µm] | 386.5 ± 35.3 b | 1050.0 ± 150.3 a | 241.6 ± 29.4 b |
| Mean filament length [µm] | 6.2 ± 0.5 | 6.6 ± 0.3 | 4.3 ± 0.3 a,b |
| Maximum filament length [µm] | 27.6 ± 2.7 | 35.6 ± 3.1 | 19.3 ± 2.6 b |
| 24 h | Ti | PPAAm | PDADMA |
| Filament number | 45.5 ± 4.9 | 48.9 ± 6.0 | 28.6 ± 4.9 b |
| Total filament length [µm] | 830.5 ± 70.0 | 863.8 ± 99.0 | 138.4 ± 33.1 a,b |
| Mean filament length [µm] | 19.0 ± 1.6 | 18.0 ± 1.9 | 4.3 ± 0.3 a,b |
| Maximum filament length [µm] | 88.8 ± 7.0 | 81.2 ± 9.1 | 14.1 ± 1.2 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gruening, M.; Dawson, J.E.; Voelkner, C.; Neuber, S.; Fricke, K.; van Rienen, U.; Speller, S.; Helm, C.A.; Nebe, J.B. Automatic Actin Filament Quantification and Cell Shape Modeling of Osteoblasts on Charged Ti Surfaces. Appl. Sci. 2021, 11, 5689. https://doi.org/10.3390/app11125689
Gruening M, Dawson JE, Voelkner C, Neuber S, Fricke K, van Rienen U, Speller S, Helm CA, Nebe JB. Automatic Actin Filament Quantification and Cell Shape Modeling of Osteoblasts on Charged Ti Surfaces. Applied Sciences. 2021; 11(12):5689. https://doi.org/10.3390/app11125689
Chicago/Turabian StyleGruening, Martina, Jonathan E. Dawson, Christian Voelkner, Sven Neuber, Katja Fricke, Ursula van Rienen, Sylvia Speller, Christiane A. Helm, and J. Barbara Nebe. 2021. "Automatic Actin Filament Quantification and Cell Shape Modeling of Osteoblasts on Charged Ti Surfaces" Applied Sciences 11, no. 12: 5689. https://doi.org/10.3390/app11125689
APA StyleGruening, M., Dawson, J. E., Voelkner, C., Neuber, S., Fricke, K., van Rienen, U., Speller, S., Helm, C. A., & Nebe, J. B. (2021). Automatic Actin Filament Quantification and Cell Shape Modeling of Osteoblasts on Charged Ti Surfaces. Applied Sciences, 11(12), 5689. https://doi.org/10.3390/app11125689