
Effects of Green Alga, Chaetomorpha aerea Extract on Non-Specific Immune Responses and Disease Resistance against Edwardsiella tarda Infection in Labeo rohita

 ,
,  ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Preparation of Extract
2.3. Experimental Design
2.4. Blood Sampling and Serum Separation
2.5. Immunological Parameters
2.5.1. Neutrophil Activity
2.5.2. Serum Lysozyme Activity
2.5.3. Myeloperoxidase Activity
2.5.4. Ceruloplasmin Activity
2.5.5. Serum Antiprotease Activity
2.5.6. Preparation of Viable Leukocytes from Peripheral Blood
2.6. Reactive Oxygen Species Production
2.7. Reactive Nitrogen Species Production
2.8. Disease Resistance
2.9. Statistical Analyses
3. Results and Discussion
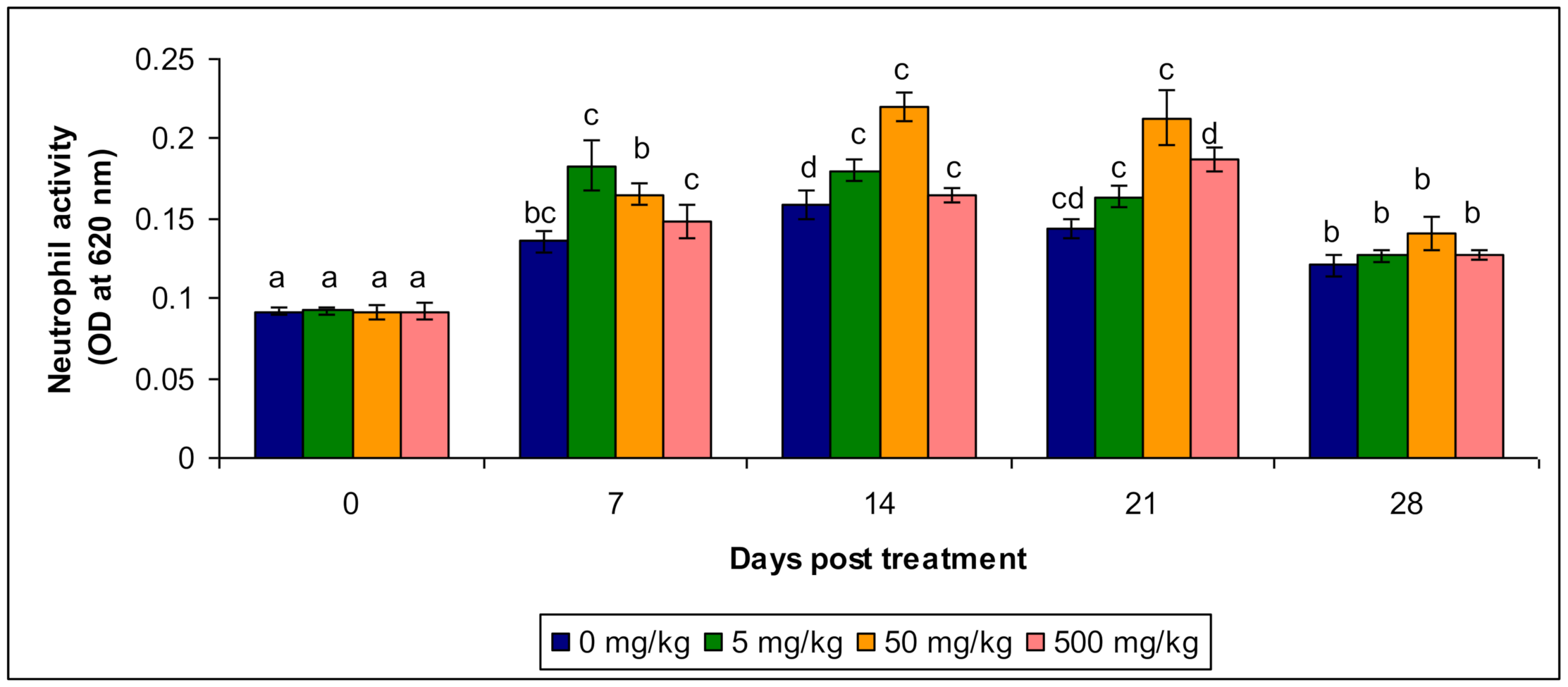
3.1. Neutrophil Activity
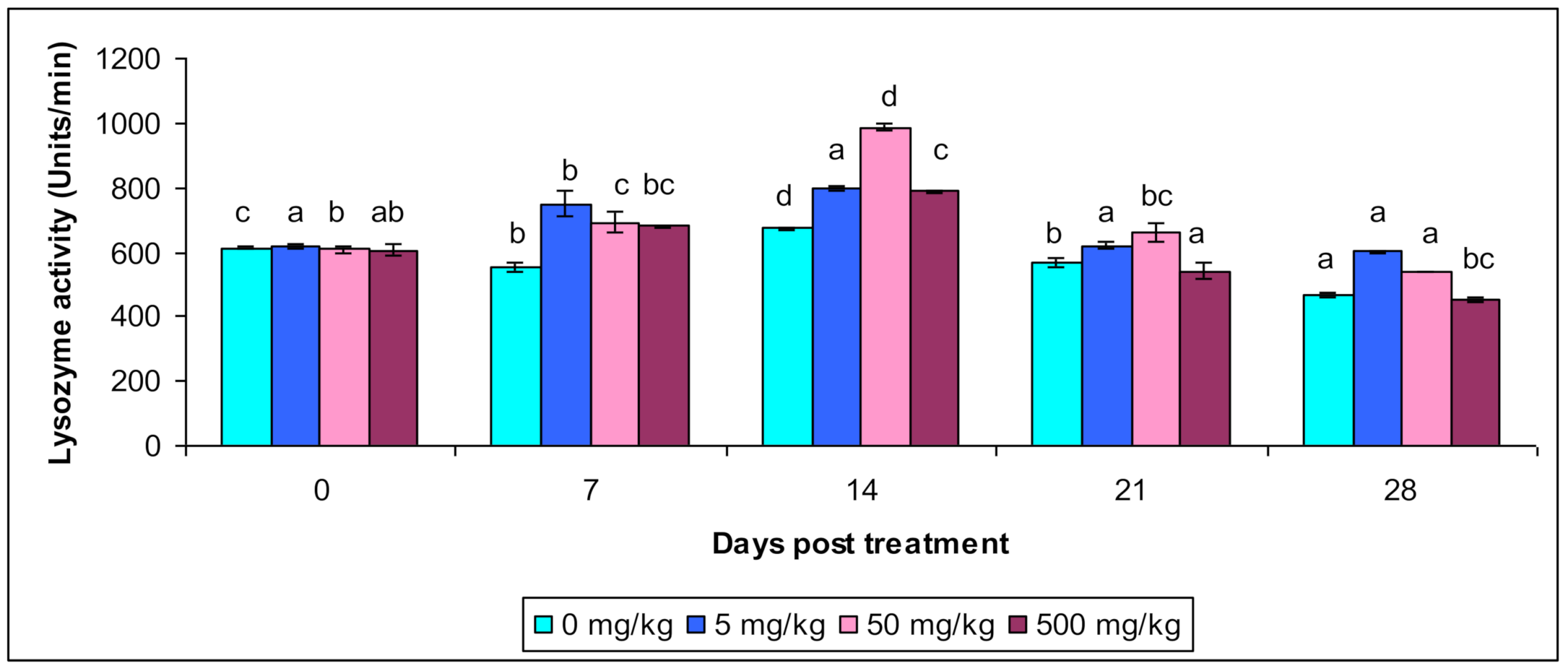
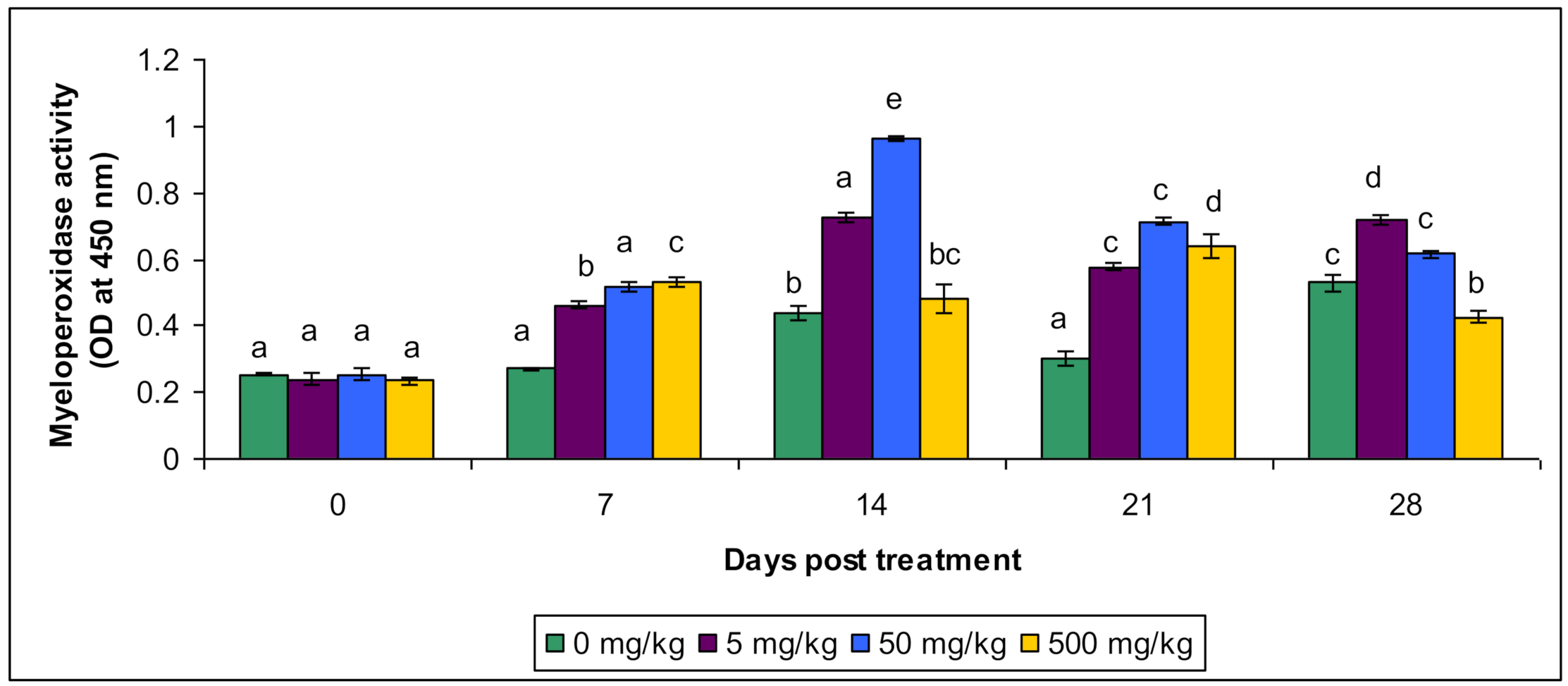
3.2. Lysozyme and Myeloperoxidase Activity
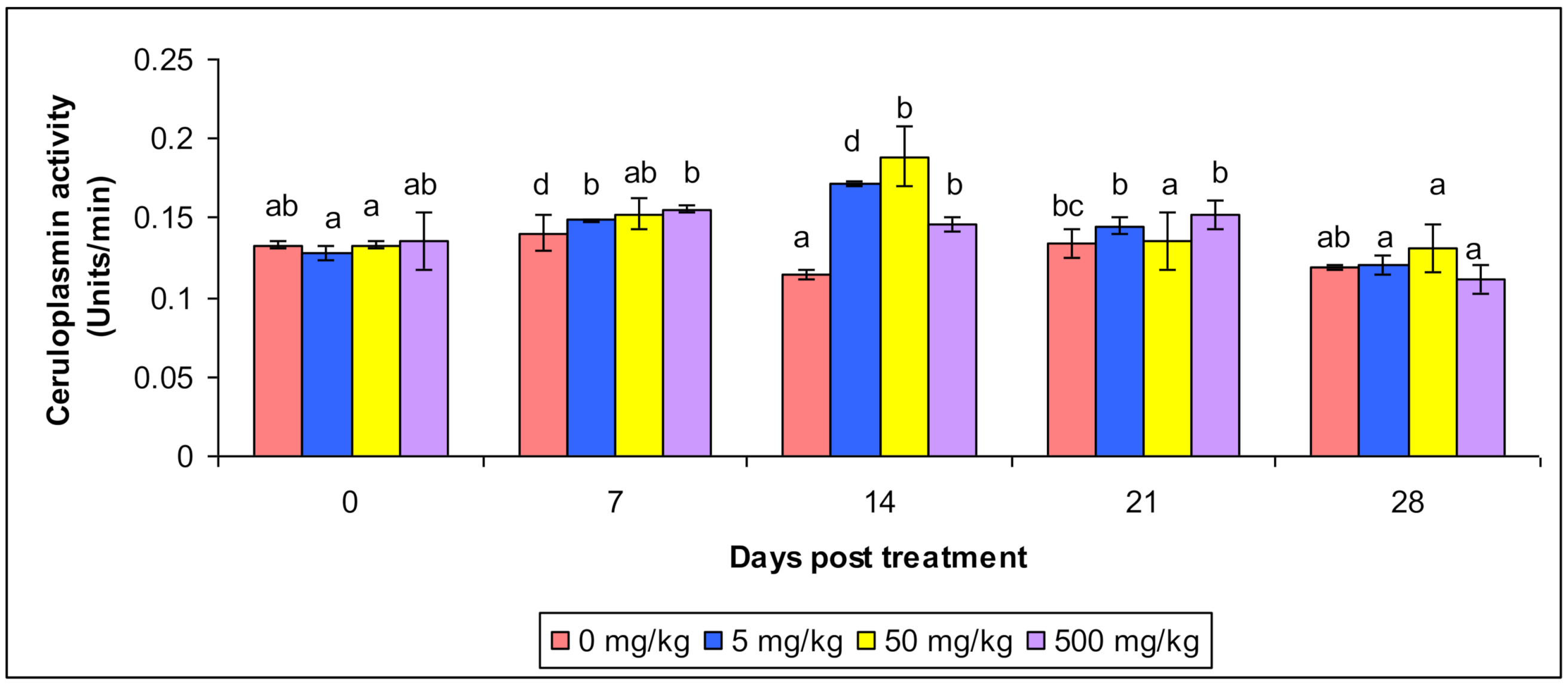
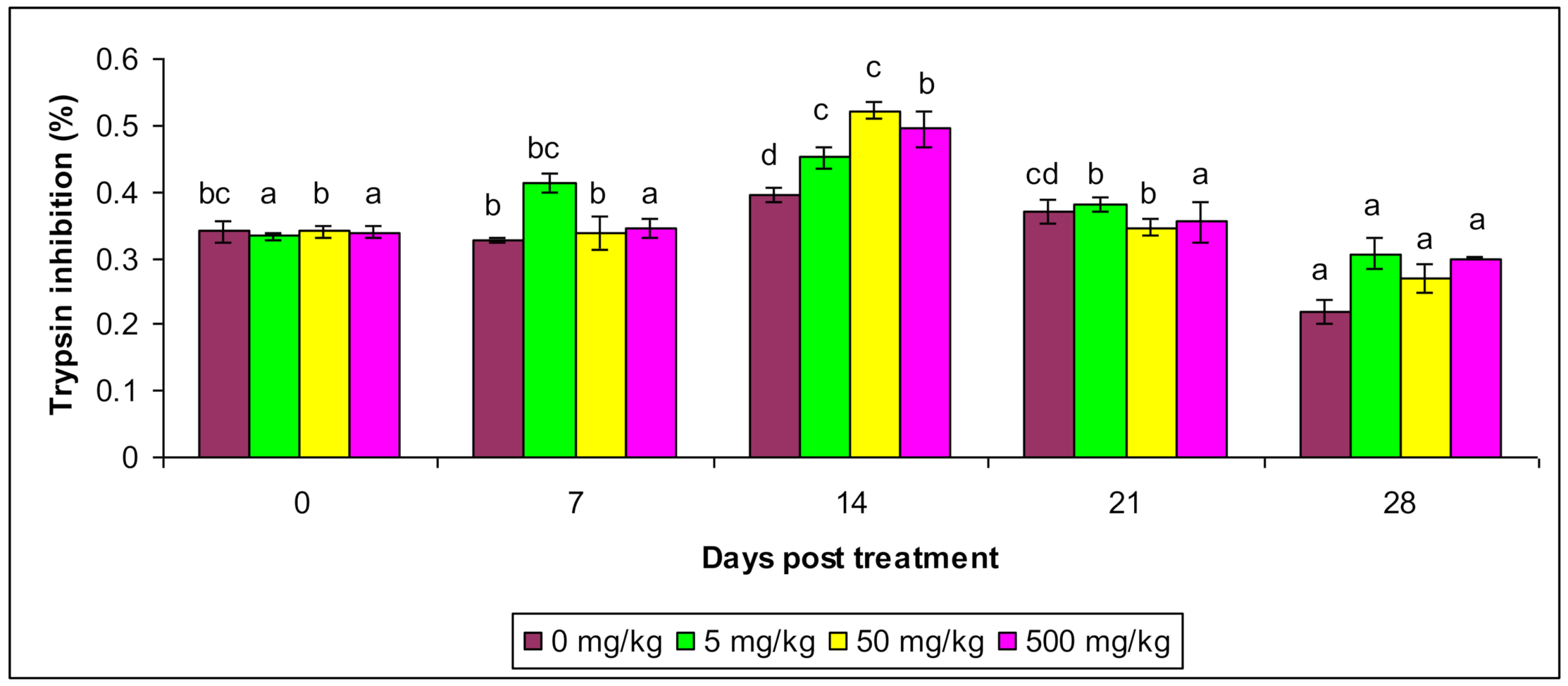
3.3. Ceruloplasmin and Antiprotease Activity
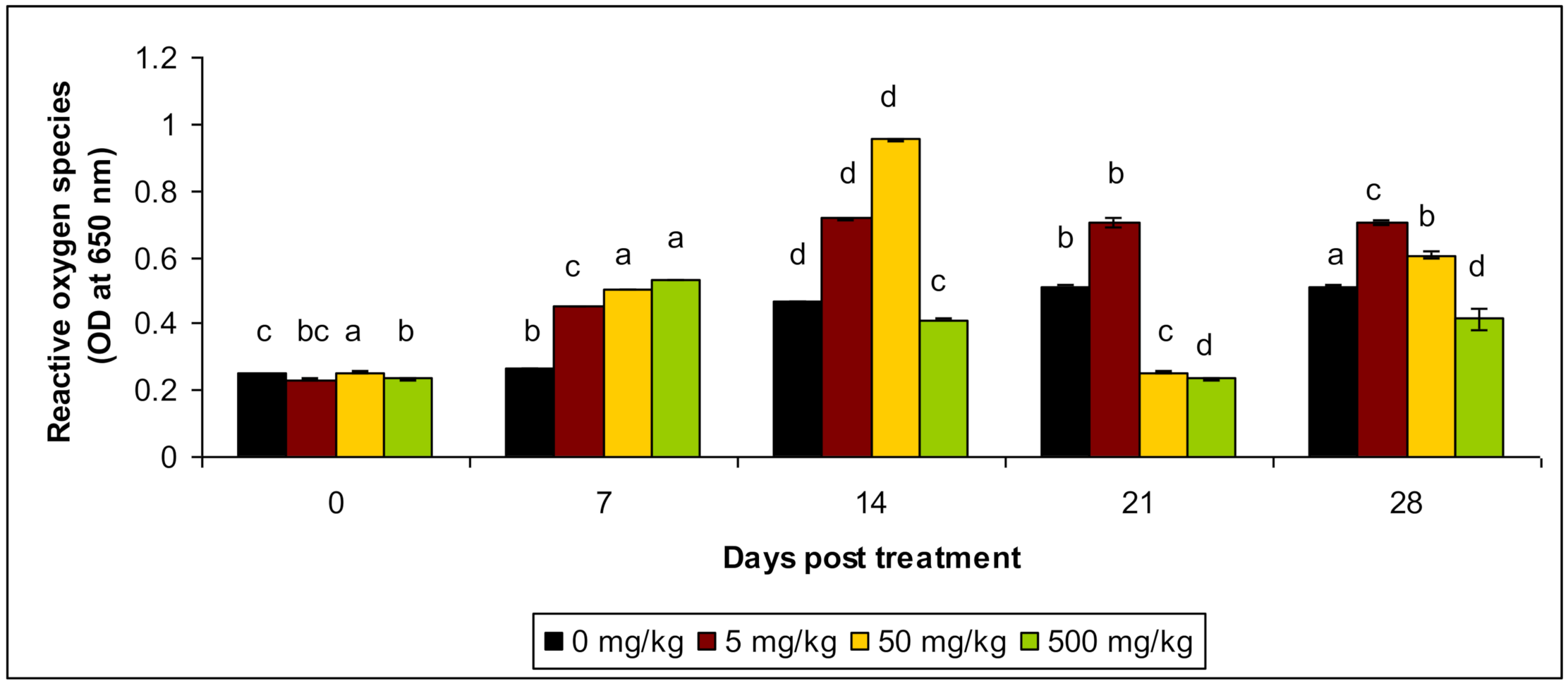
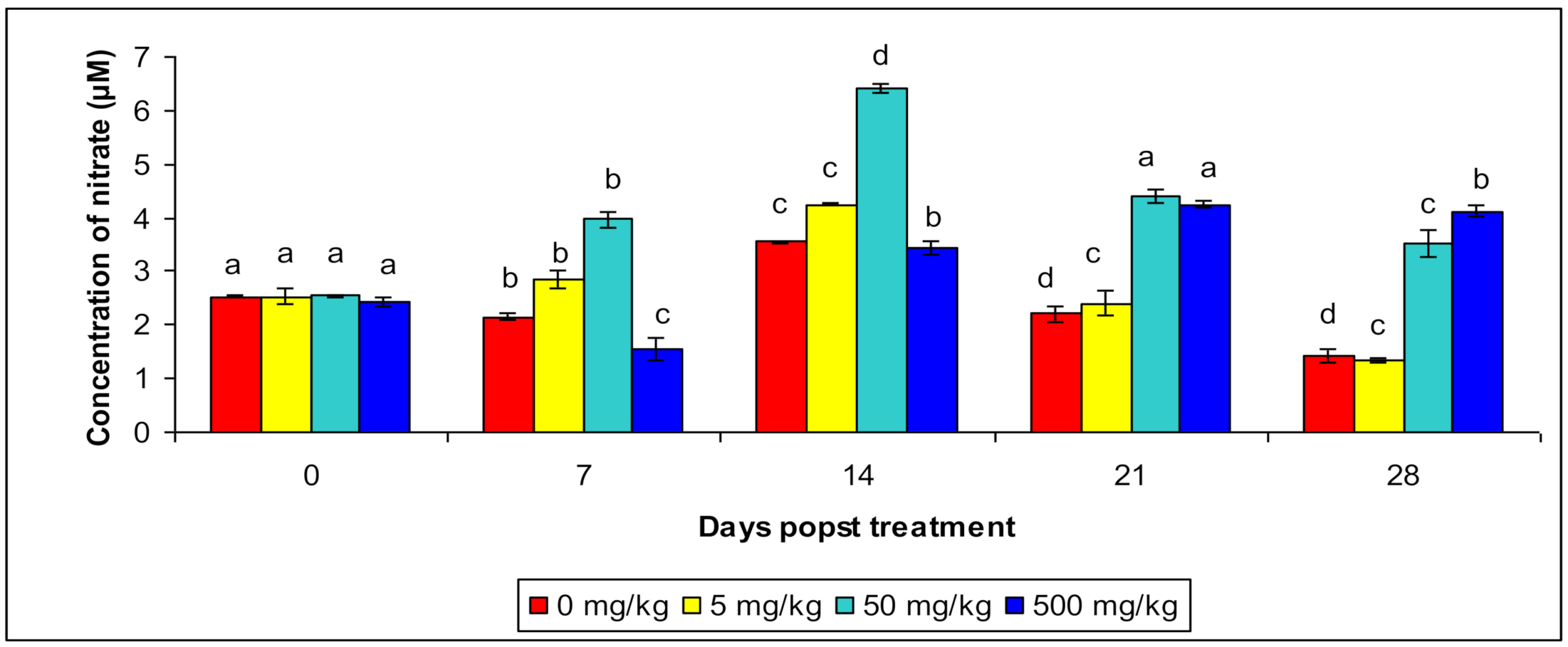
3.4. Reactive Oxygen Species (ROS) and Reactive Nitrogen Species (RNS) Production
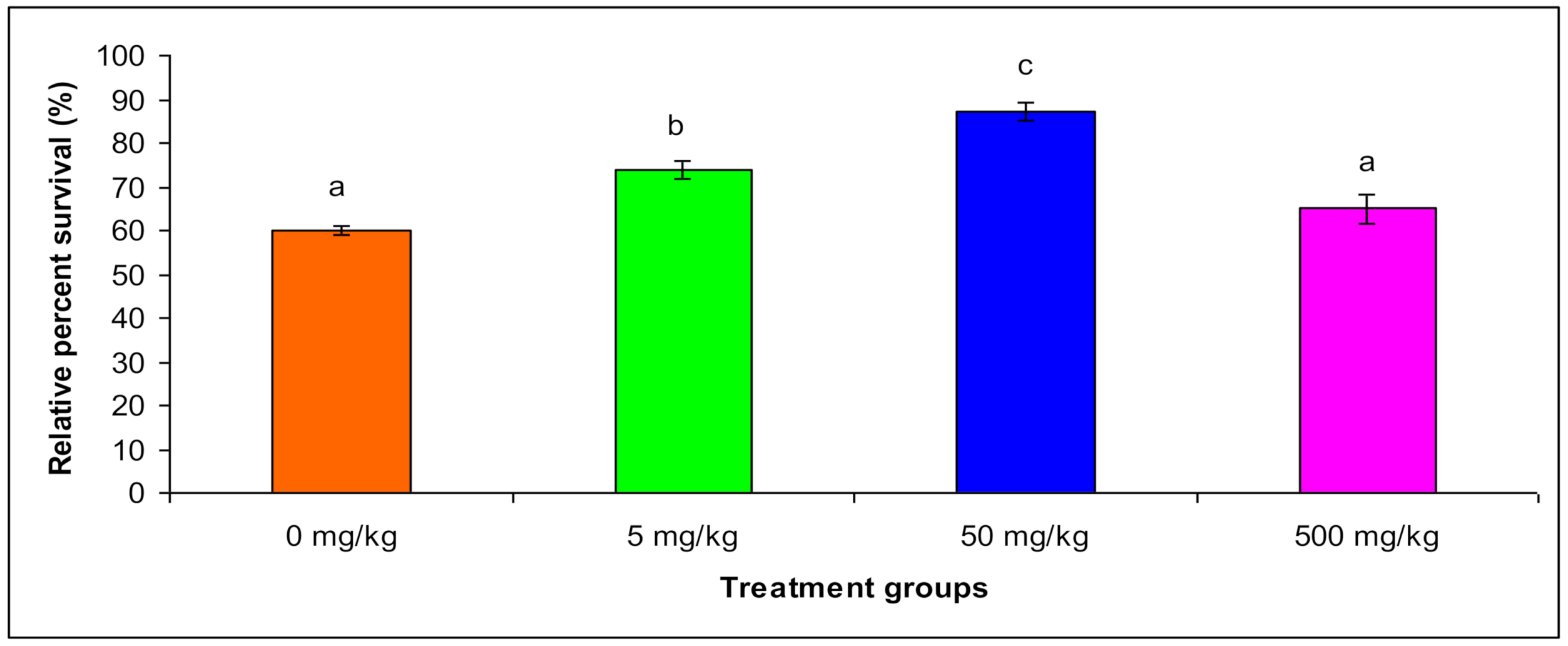
3.5. Relative Percent Survival
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ye, Y.; Barange, M.; Beveridge, M.; Garibaldi, L.; Gutierrez, N.; Anganuzzi, A.; Taconet, M. FAO’s statistic data and sustainability of fisheries and aquaculture: Comments on Pauly and Zeller (2017). Mar. Policy 2017, 81, 401–405. [Google Scholar] [CrossRef]
- Kar, B.; Mohanty, J.; Hemaprasanth, K.P.; Sahoo, P.K. The immune response in rohu, Labeo rohita (Actinopterygii: Cyprinidae) toArgulus siamensis (Branchiura: Argulidae) infection: Kinetics of immune gene expression and innate immune response. Aquac. Res. 2013, 46, 1292–1308. [Google Scholar] [CrossRef]
- Van Hai, N. The use of medicinal plants as immunostimulants in aquaculture: A review. Aquaculture 2015, 446, 88–96. [Google Scholar] [CrossRef]
- Giri, S.; Sukumaran, V.; Sen, S.; Jena, P. Effects of dietary supplementation of potential probiotic Bacillus subtilisVSG1 singularly or in combination with Lactobacillus plantarumVSG3 or/and Pseudomonas aeruginosaVSG2 on the growth, immunity and disease resistance of Labeo rohita. Aquac. Nutr. 2013, 20, 163–171. [Google Scholar] [CrossRef]
- Ninawe, A. DNA vaccination and prophylactic measures in aquaculture health management. Aquac. Asia 2006, 11, 21. [Google Scholar]
- Anderson, D.P. Immunostimulants, adjuvants, and vaccine carriers in fish: Applications to aquaculture. Annu. Rev. Fish Dis. 1992, 2, 281–307. [Google Scholar] [CrossRef]
- Swain, P.; Dash, S.; Sahoo, P.K.; Routray, P.; Sahoo, S.K.; Gupta, S.D.; Meher, P.K.; Sarangi, N. Non-specific immune parameters of brood Indian major carp Labeo rohita and their seasonal variations. Fish Shellfish. Immunol. 2007, 22, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Harikrishnan, R.; Balasundaram, C.; Heo, M.-S. Impact of plant products on innate and adaptive immune system of cultured finfish and shellfish. Aquaculture 2011, 317, 1–15. [Google Scholar] [CrossRef]
- Vaseeharan, B.; Thaya, R. Medicinal plant derivatives as immunostimulants: An alternative to chemotherapeutics and antibiotics in aquaculture. Aquac. Int. 2014, 22, 1079–1091. [Google Scholar] [CrossRef]
- Yuan, C.; Li, D.; Chen, W.; Sun, F.; Wu, G.; Gong, Y.; Tang, J.; Shen, M.; Han, X. Administration of a herbal immunoregulation mixture enhances some immune parameters in carp (Cyprinus carpio). Fish Physiol. Biochem. 2007, 33, 93–101. [Google Scholar] [CrossRef]
- Divyagnaneswari, M.; Christybapita, D.; Michael, R.D. Enhancement of nonspecific immunity and disease resistance in Oreochromis mossambicus by Solanum trilobatum leaf fractions. Fish Shellfish. Immunol. 2007, 23, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Citarasu, T.; Venkatramalingam, K.; Babu, M.M.; Sekar, R.R.J.; Petermarian, M. Influence of the antibacterial herbs, Solanum trilobatum, Andrographis paniculata and Psoralea corylifolia on the survival, growth and bacterial load of Penaeus monodon post larvae. Aquac. Int. 2003, 11, 581–595. [Google Scholar] [CrossRef]
- Prasad, G.; Mukthiraj, S. Effect of methanolic extract of Andrographis paniculata (Nees) on growth and haematology of Oreochromis mossambicus (Peters). World J. Fish Mar. Sci. 2011, 3, 473–479. [Google Scholar]
- Sahoo, P.K.; Mukherjee, S.C. The effect of dietary immunomodulation upon Edwardsiella tarda vaccination in healthy and immunocompromised Indian major carp (Labeo rohita). Fish Shellfish. Immunol. 2002, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Shobana, S.; Kumar, G.; Bakonyi, P.; Saratale, G.D.; Al-Muhtaseb, A.H.; Nemestóthy, N.; Bélafi-Bakó, K.; Xia, A.; Chang, J.-S. A review on the biomass pretreatment and inhibitor removal methods as key-steps towards efficient macroalgae-based biohydrogen production. Bioresour. Technol. 2017, 244, 1341–1348. [Google Scholar] [CrossRef]
- Daneshvar, E.; Zarrinmehr, M.J.; Kousha, M.; Hashtjin, A.M.; Saratale, G.D.; Maiti, A.; Vithanage, M.; Bhatnagar, A. Hexavalent chromium removal from water by microalgal-based materials: Adsorption, desorption and recovery studies. Bioresour. Technol. 2019, 293, 122064. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Leung, T.L.F.; Bates, A.E. More rapid and severe disease outbreaks for aquaculture at the tropics: Implications for food security. J. Appl. Ecol. 2012, 50, 215–222. [Google Scholar] [CrossRef]
- Peeler, E.J.; Taylor, N.G. The application of epidemiology in aquatic animal health -opportunities and challenges. Vet. Res. 2011, 42, 94. [Google Scholar] [CrossRef]
- Swaminathan, T.R.; Kumar, R.; Dharmaratnam, A.; Basheer, V.S.; Sood, N.; Pradhan, P.K.; Sanil, N.K.; Vijayagopal, P.; Jena, J. Emergence of carp edema virus in cultured ornamental koi carp, Cyprinus carpio koi, in India. J. Gen. Virol. 2016, 97, 3392–3399. [Google Scholar] [CrossRef]
- Sattanathan, G.; Tamizhazhagan, V.; Padmapriya, S.; Liu, W.-C.; Balasubramanian, B. Effect of Green Algae Chaetomorpha antennina Extract on Growth, Modulate Immunity, and Defenses against Edwardsiella tarda Infection in Labeo rohita. Animals 2020, 10, 2033. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M.; Snader, K.M. Natural Products as Sources of New Drugs over the Period 1981−2002. J. Nat. Prod. 2003, 66, 1022–1037. [Google Scholar] [CrossRef]
- Bricknell, I.; Dalmo, R.A. The use of immunostimulants in fish larval aquaculture. Fish Shellfish. Immunol. 2005, 19, 457–472. [Google Scholar] [CrossRef] [PubMed]
- Sattanathan, G.; Palanisamy, T.; Padmapriya, S.; Arumugam, V.A.; Park, S.; Kim, I.H.; Balasubramanian, B. Influences of dietary inclusion of algae Chaetomorpha aerea enhanced growth performance, immunity, haematological response and disease resistance of Labeo rohita challenged with Aeromonas hydrophila. Aquac. Rep. 2020, 17, 100353. [Google Scholar] [CrossRef]
- Talwar, P.K. Inland Fishes of India and adjacent Countries; CRC Press: New Delhi, India, 1991; Volume 2. [Google Scholar]
- Plumb, J. Edwardsiella Septicaemias. In Viral, Bacterial and Fungal Infections; Woo, P.T.K., Bruno, D.W., Eds.; Fish Disease and Disorders Volume 3; CABI Publishing: Wallingford, UK, 1999. [Google Scholar]
- Sattanathan, G.; Sournamanikam, V. Cow urine distillate as an eco safe and economical feed additive for enhancing growth, food utilization and survival rate in rohu, labeo rohita (Hamilton). J. Fish. Aquat. Sci. 2017, 12, 241–252. [Google Scholar] [CrossRef]
- Kirubakaran, C.J.W.; Alexander, C.P.; Michael, R.D. Enhancement of non-specific immune responses and disease resistance on oral administration of Nyctanthes arbortristis seed extract in Oreochromis mossambicus (Peters). Aquac. Res. 2010, 41, 1630–1639. [Google Scholar] [CrossRef]
- Dinakaran Michael, R.; Srinivas, S.; Sailendri, K.; Muthukkaruppan, V. A rapid method for repetitive bleeding in fish. Indian J. Exp. Biol. 1994, 32, 838. [Google Scholar]
- Stasiack, A.; Bauman, C. Netrophil activity as a potential indicator immunomodulator Ecteinascida turbinata extract on Edwardsiella ictaluri infection of channel catfish. J. Aquat. Anim. Health 1996, 7, 141–146. [Google Scholar]
- Parry, R.M., Jr.; Chandan, R.C.; Shahani, K.M. A Rapid and Sensitive Assay of Muramidase. Proc. Soc. Exp. Biol. Med. 1965, 119, 384–386. [Google Scholar] [CrossRef]
- Sahoo, P.; Rauta, P.; Mohanty, B.; Mahapatra, K.; Saha, J.; Rye, M.; Eknath, A. Selection for improved resistance to Aeromonas hydrophila in Indian major carp Labeo rohita: Survival and innate immune responses in first generation of resistant and susceptible lines. Fish Shellfish. Immunol. 2011, 31, 432–438. [Google Scholar] [CrossRef]
- Dautremepuits, C.; Betoulle, S.; Paris-Palacios, S.; Vernet, G. Humoral immune factors modulated by copper and chitosan in healthy or parasitised carp (Cyprinus carpio L.) by Ptychobothrium sp. (Cestoda). Aquat. Toxicol. 2004, 68, 325–338. [Google Scholar] [CrossRef]
- Rao, Y.V.; Chakrabarti, R. Enhanced anti-proteases in Labeo rohita fed with diet containing herbal ingredients. Indian J. Clin. Biochem. 2004, 19, 132–134. [Google Scholar] [CrossRef]
- Strober, W. Trypan blue exclusion test of cell viability. Curr. Protoc. Immunol. 1997, 21, A.3B.1–A.3B.2. [Google Scholar]
- Secombes, C.J. Isolation of salmonid macrophages and analysis of their killing activity. Tech. Fish Immunol. 1990, 1, 137–154. [Google Scholar]
- Palić, D.; Andreasen, C.B.; Menzel, B.W.; Roth, J.A. A rapid, direct assay to measure degranulation of primary granules in neutrophils from kidney of fathead minnow (Pimephales promelas Rafinesque, 1820). Fish Shellfish. Immunol. 2005, 19, 217–227. [Google Scholar] [CrossRef]
- Amend, D.F. Potency testing of fish vaccines. Fish Biol. Serodiagn. Vaccines 1981, 49, 447–454. [Google Scholar]
- Gora, A.H.; Sahu, N.P.; Sahoo, S.; Rehman, S.; Dar, S.A.; Ahmad, I.; Agarwal, D. Effect of dietary Sargassum wightii and its fucoidan-rich extract on growth, immunity, disease resistance and antimicrobial peptide gene expression in Labeo rohita. Int. Aquat. Res. 2018, 10, 115–131. [Google Scholar] [CrossRef]
- Salton, M.R.J.; Ghuysen, J.M. The structure of di- and tetra-saccharides released from cell walls by lysozyme and streptomyces F1 enzyme and the β (1→4) N-acetylhexosaminidase activity of these enzymes. Biochim. Biophys. Acta Bioenerg. 1959, 36, 552–554. [Google Scholar] [CrossRef]
- Glynn, A.A. The complement lysozyme sequence in immune bacteriolysis. Immunology 1969, 16, 463–471. [Google Scholar]
- Ghiasi, F.; Mirzargar, S.S.; Badakhshan, H.; Shamsi, S. Leukocyte Count and Phagocytic Index in Cyprinus carpio under the Wintering Conditions. J. Fish. Aquat. Sci. 2010, 5, 113–119. [Google Scholar] [CrossRef]
- Sivaram, V.; Babu, M.; Immanuel, G.; Murugadass, S.; Citarasu, T.; Marian, M. Growth and immune response of juvenile greasy groupers (Epinephelus tauvina) fed with herbal antibacterial active principle supplemented diets against Vibrio harveyi infections. Aquaculture 2004, 237, 9–20. [Google Scholar] [CrossRef]
- Rao, Y.V.; Das, B.; Jyotyrmayee, P.; Chakrabarti, R. Effect of Achyranthes aspera on the immunity and survival of Labeo rohita infected with Aeromonas hydrophila. Fish Shellfish. Immunol. 2006, 20, 263–273. [Google Scholar] [CrossRef]
- Amar, E.C.; Kiron, V.; Satoh, S.; Watanabe, T. Enhancement of innate immunity in rainbow trout (Oncorhynchus mykiss Walbaum) associated with dietary intake of carotenoids from natural products. Fish Shellfish. Immunol. 2004, 16, 527–537. [Google Scholar] [CrossRef]
- Amat-García, E.C.; Amat-García, G.D.; Henaom, L.G. Diversidad taxonómica y ecológica de la entomofauna micófaga en un bosque altoandino de la cordillera oriental de Colombia. Ecología 2004, 28, 223–231. [Google Scholar]
- Bilen, S.; Ünal, S.; Güvensoy, H. Effects of oyster mushroom (Pleurotus ostreatus) and nettle (Urtica dioica) methanolic extracts on immune responses and resistance to Aeromonas hydrophila in rainbow trout (Oncorhynchus mykiss). Aquaculture 2016, 454, 90–94. [Google Scholar] [CrossRef]
- Olivier, G.; Evelyn, T.; Lallier, R. Immunity to aeromonas salmonicida in coho salmon (oncorhynchus kisutch) inducfd by modified freund’s complete adjuvant: Its non-specific nature and the probable role of macrophages in the phenomenon. Dev. Comp. Immunol. 1985, 9, 419–432. [Google Scholar] [CrossRef]
- Sahoo, P.; Mahapatra, K.D.; Saha, J.; Barat, A.; Sahoo, M.; Mohanty, B.; Gjerde, B.; Ødegård, J.; Rye, M.; Salte, R. Family association between immune parameters and resistance to Aeromonas hydrophila infection in the Indian major carp, Labeo rohita. Fish Shellfish. Immunol. 2008, 25, 163–169. [Google Scholar] [CrossRef]
- Ellis, A. Innate host defense mechanisms of fish against viruses and bacteria. Dev. Comp. Immunol. 2001, 25, 827–839. [Google Scholar] [CrossRef]
- Pradhan, J.; Das, S.; Das, B.K. Antibacterial activity of freshwater microalgae: A review. Afr. J. Pharm. Pharmacol. 2014, 8, 809–818. [Google Scholar]
- Mohanty, B.; Sahoo, P.; Mahapatra, K.; Saha, J. Innate immune responses in families of Indian major carp, Labeo rohita, differing in their resistance to Edwardsiella tarda infection. Curr. Sci. 2007, 92, 1270–1274. [Google Scholar]
- Miyazaki, T. A Simple Method to Evaluate Respiratory Burst Activity of Blood Phagocytes from Japanese Flounder. Fish Pathol. 1998, 33, 141–142. [Google Scholar] [CrossRef]
- Jang, S.I.; Marsden, M.J.; Secombes, C.J.; Choi, M.S.; Kim, Y.G.; Kim, K.J.; Chung, H.-T. Effect of glycyrrhizin on rainbow trout Oncorhynchus mykiss leukocyte responses. J. Korean Soc. Microbiol. 1998, 33, 263–271. [Google Scholar]
- Peddie, S.; Zou, J.; Secombes, C.J. Immunostimulation in the rainbow trout (Oncorhynchus mykiss) following intraperitoneal administration of Ergosan. Vet. Immunol. Immunopathol. 2002, 86, 101–113. [Google Scholar] [CrossRef]
- Sakai, M. Current research status of fish immunostimulants. Aquaculture 1999, 172, 63–92. [Google Scholar] [CrossRef]
- Citarasu, T.; Sivaram, V.; Immanuel, G.; Rout, N.; Murugan, V. Influence of selected Indian immunostimulant herbs against white spot syndrome virus (WSSV) infection in black tiger shrimp, Penaeus monodon with reference to haematological, biochemical and immunological changes. Fish Shellfish. Immunol. 2006, 21, 372–384. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Do Jeong, H. Enhanced resistance against Edwardsiella tarda infection in tilapia (Oreochromis niloticus) by administration of protein-bound polysaccharide. Aquaculture 1996, 143, 135–143. [Google Scholar] [CrossRef]
- Jacobson, D.M.; Andersen, R.A. The discovery of mixotrophy in photosynthetic species of Dinophysis (Dinophyceae): Light and electron microscopical observations of food vacuoles in Dinophysis acuminata, D. norvegica and two heterotrophic dinophysoid dinoflagellates. Phycologia 1994, 33, 97–110. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Groups | Days | ||||
|---|---|---|---|---|---|---|
| 0 Days | 7 Days | 14 Days | 21 Days | 28 Days | ||
| Neutrophil Activity | Control | 0.098 a ± 0.013 | 0.134 bc ± 0.00632 | 0.156 d ± 0.00272 | 0.143 c ± 0.0034 | 0.123 b ± 0.0014 |
| T1 | 0.093 a ± 0.00033 | 0.185 c ± 0.0022 | 0.187 c ± 0.00323 | 0.164 b ± 0.0035 | 0.125 b ± 0.0014 | |
| T2 | 0.096 a ± 0.0012 | 0.166 b ± 0.0023 | 0.226 c ± 0.00214 | 0.201 a ± 0.0033 | 0.143 b ± 0.00434 | |
| T3 | 0.091 a ± 0.00134 | 0.145 c ± 0.00343 | 0.168 c ± 0.013 | 0.184 b ± 0.001854 | 0.132 b ± 0.00524 | |
| Lysozyme Activity | Control | 609.4 c ± 0.39 | 565.4 b ± 0.43 | 678.3 d ± 0.582 | 545.3 b ± 0.34 | 464.3 a ± 0.212 |
| T1 | 612.4 a ± 0.423 | 745.54 b ± 0.48 | 798.45 a ± 0.743 | 612.43 a ± 0.85 | 600.3 a ± 0.88 | |
| T2 | 603.5 b ± 0.65 | 689.3 c ± 0.24 | 995.35 d ± 0.17 | 690.43 bc ± 0.43 | 536.4 a ± 0.34 | |
| T3 | 603.4 ab ± 0.64 | 678.3 bc ± 0.83 | 789.32 c ± 0.13 | 546.43 a ± 0.54 | 453.2 bc ± 0.18 | |
| Myeloperoxidase Activity | Control | 0.253 a ± 0.083 | 0.267 a ± 0.072 | 0.467 b ± 0.068 | 0.332 a ± 0.064 | 0.512 c ± 0.094 |
| T1 | 0.233 a ± 0.059 | 0.454 b ± 0.083 | 0.716 a ± 0.0783 | 0.564 c ± 0.085 | 0.703 b ± 0.084 | |
| T2 | 0.254 a ± 0.082 | 0.503 a ± 0.074 | 0.954 e ± 0.074 | 0.696 c ± 0.093 | 0.605 c ± 0.0564 | |
| T3 | 0.234 a ± 0.0734 | 0.534 c ± 0.0833 | 0.412 bc ± 0.083 | 0.624 d ± 0.0854 | 0.414 b ± 0.0824 | |
| Ceruloplasmin Activity | Control | 0.137 ab ± 0.0083 | 0.129 d ± 0.0082 | 0.12 a ± 0.0079 | 0.139 bc ± 0.0084 | 0.119 ab ± 0.0094 |
| T1 | 0.133 a ± 0.0089 | 0.149 b ± 0.0093 | 0.173 d ± 0.0083 | 0.145 b ± 0.005 | 0.127 a ± 0.0034 | |
| T2 | 0.135 a ± 0.0012 | 0.154 ab ± 0.0074 | 0.189 b ± 0.0057 | 0.138 a ± 0.0053 | 0.106 a ± 0.00634 | |
| T3 | 0.133 ab ± 0.0086 | 0.159 b ± 0.0089 | 0.144 b ± 0.013 | 0.161 b ± 0.0077 | 0.111 a ± 0.00724 | |
| Antiprotease Activity | Control | 0.353 bc ± 0.083 | 0.327 b ± 0.055 | 0.396 d ± 0.0538 | 0.395 d ± 0.0053 | 0.209 a ± 0.0084 |
| T1 | 0.334 a ± 0.0089 | 0.404 bc ± 0.0043 | 0.452 c ± 0.00623 | 0.365 b ± 0.05 | 0.308 a ± 0.034 | |
| T2 | 0.332 b ± 0.008 | 0.35 b ± 0.0034 | 0.546 c ± 0.033 | 0.336 b ± 0.053 | 0.274 a ± 0.003 | |
| T3 | 0.353 a ± 0.00454 | 0.367 a ± 0.0034 | 0.505 b ± 0.0153 | 0.353 a ± 0.00143 | 0.307 a ± 0.00254 | |
| Reactive Oxygen Species | Control | 0.253 c ± 0.0123 | 0.267 b ± 0.0532 | 0.467 d ± 0.0332 | 0.332 b ± 0.0432 | 0.512 a ± 0.0412 |
| T1 | 0.233 bc ± 0.0123 | 0.454 c ± 0.043 | 0.716 d ± 0.0543 | 0.564 b ± 0.035 | 0.703 c ± 0.0154 | |
| T2 | 0.254 a ± 0.015 | 0.503 a ± 0.024 | 0.954 d ± 0.0214 | 0.696 c ± 0.0143 | 0.605 b ± 0.0234 | |
| T3 | 0.234 b ± 0.0244 | 0.534 a ± 0.0143 | 0.412 c ± 0.0213 | 0.624 d ± 0.0254 | 0.414 d ± 0.0224 | |
| Reactive Nitrogen Species | Control | 2.53 a ± 0.203 | 2.15 b ± 0.12 | 3.53 c ± 0.134 | 2.151 d ± 0.156 | 1.523 d ± 0.387 |
| T1 | 2.65 a ± 0.21 | 2.88 b ± 0.54 | 4.22 c ± 0.21 | 2.04 c ± 0.23 | 1.35 c ± 0.219 | |
| T2 | 2.54 a ± 0.25 | 2.98 b ± 0.18 | 6.43 d ± 0.245 | 4.32 a ± 0.383 | 3.13 c ± 0.384 | |
| T3 | 2.43 a ± 0.084 | 1.54 c ± 0.12 | 3.25 b ± 0.213 | 4.33 a ± 0.215 | 4.25 b ± 0.214 | |
| Relative Percent Survival | Control | - | - | - | - | 58.4 a ± 4 |
| T1 | - | - | - | - | 70.54 b ± 7 | |
| T2 | - | - | - | - | 85.3 c ± 12 | |
| T3 | - | - | - | - | 60.43 a ± 16 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sattanathan, G.; Tamizhazhagan, V.; Raza, N.; Shah, S.Q.A.; Hussain, M.Z.; Kim, K.-H. Effects of Green Alga, Chaetomorpha aerea Extract on Non-Specific Immune Responses and Disease Resistance against Edwardsiella tarda Infection in Labeo rohita. Appl. Sci. 2021, 11, 4325. https://doi.org/10.3390/app11104325
Sattanathan G, Tamizhazhagan V, Raza N, Shah SQA, Hussain MZ, Kim K-H. Effects of Green Alga, Chaetomorpha aerea Extract on Non-Specific Immune Responses and Disease Resistance against Edwardsiella tarda Infection in Labeo rohita. Applied Sciences. 2021; 11(10):4325. https://doi.org/10.3390/app11104325
Chicago/Turabian StyleSattanathan, Govindharajan, Vairakannu Tamizhazhagan, Nadeem Raza, Syed Qaswar Ali Shah, Muhammad Zubair Hussain, and Ki-Hyun Kim. 2021. "Effects of Green Alga, Chaetomorpha aerea Extract on Non-Specific Immune Responses and Disease Resistance against Edwardsiella tarda Infection in Labeo rohita" Applied Sciences 11, no. 10: 4325. https://doi.org/10.3390/app11104325
APA StyleSattanathan, G., Tamizhazhagan, V., Raza, N., Shah, S. Q. A., Hussain, M. Z., & Kim, K.-H. (2021). Effects of Green Alga, Chaetomorpha aerea Extract on Non-Specific Immune Responses and Disease Resistance against Edwardsiella tarda Infection in Labeo rohita. Applied Sciences, 11(10), 4325. https://doi.org/10.3390/app11104325