An Updated Review of Smac Mimetics, LCL161, Birinapant, and GDC-0152 in Cancer Treatment



Abstract
1. Introduction
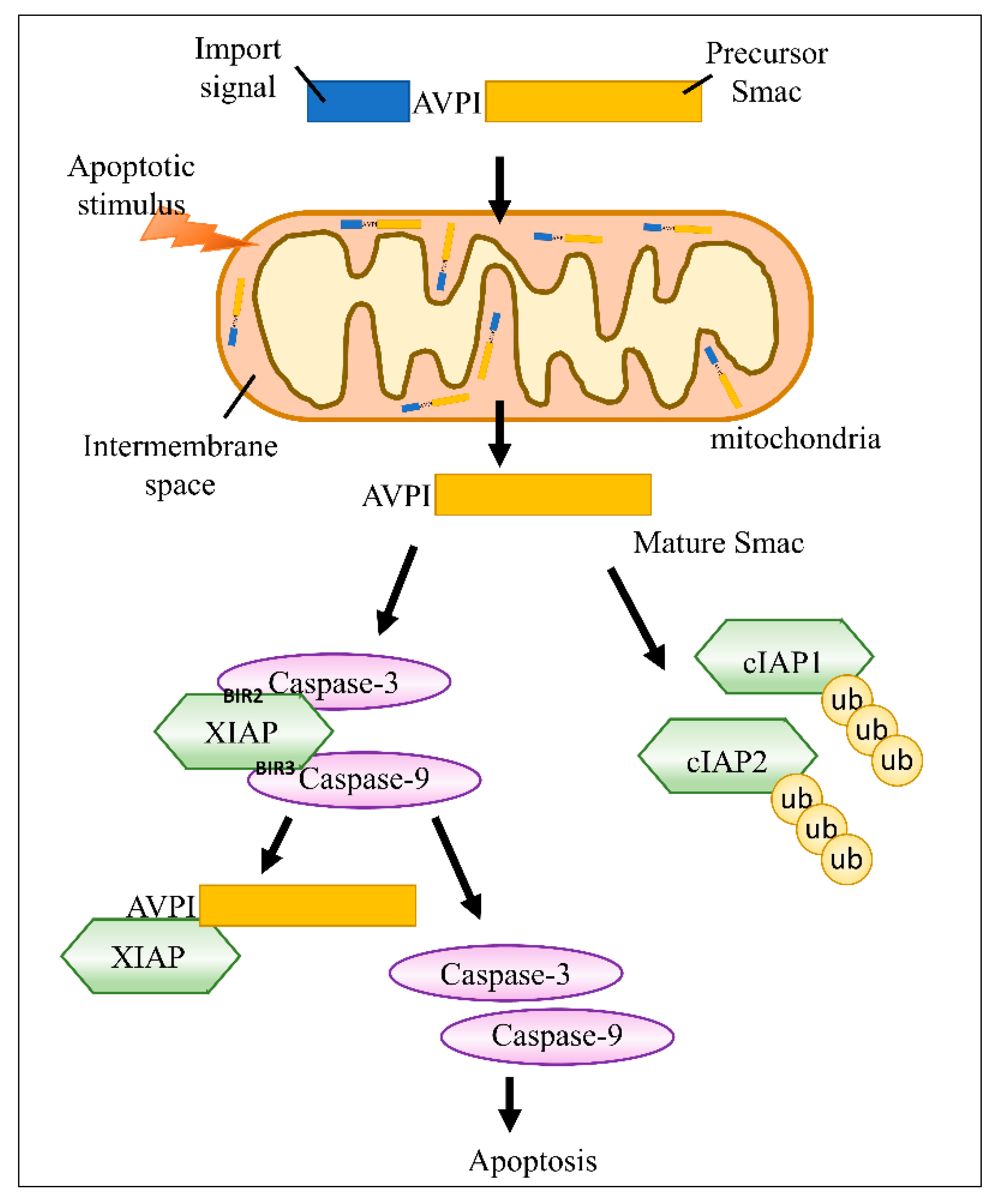
2. Smac/DIABLO Inhibits IAPs in Cancer
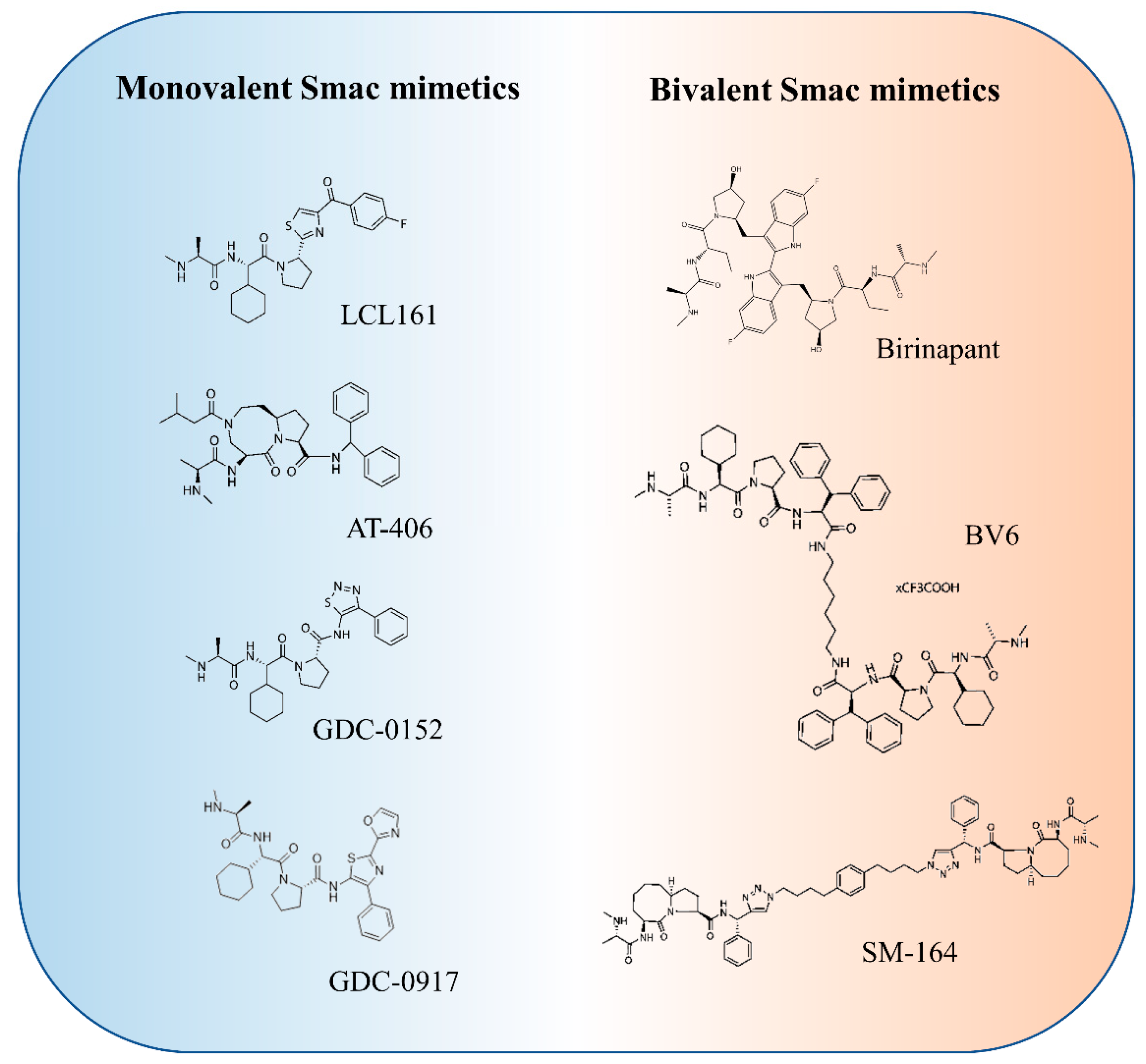
3. Smac Mimetics for Cancer Treatment
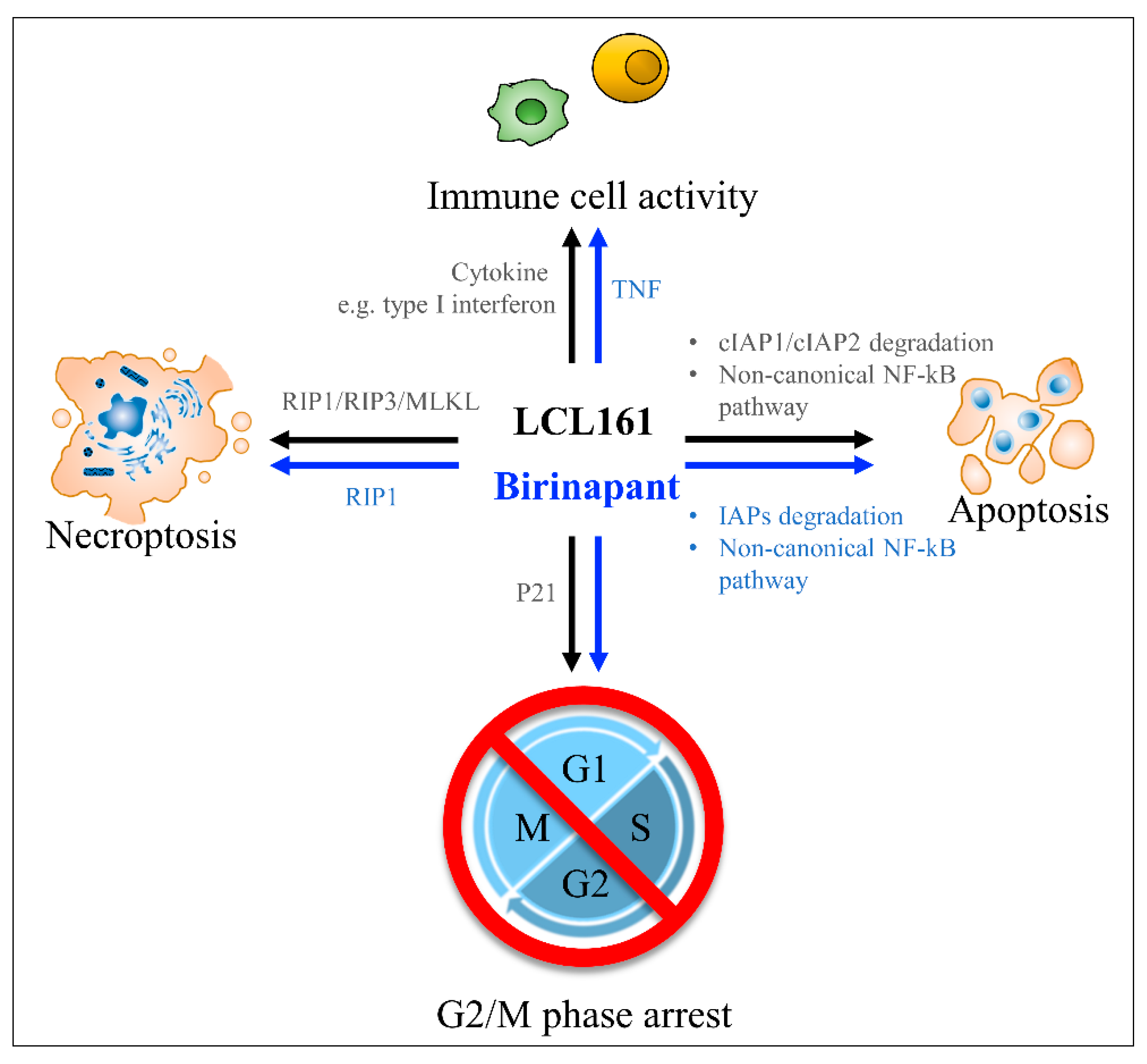
4. Anti-Cancer Molecular Mechanism of LCL161
5. LCL161 Combination Treatment
6. Current Status of LCL161 in Clinical Trials
7. Anti-Cancer Molecular Mechanisms of Birinapant (TL32711)
8. Combination Therapy with Birinapant for Cancer Treatment
9. Current Status of Birinapant in Clinical Trials
10. GDC-0152 for Cancer Treatment
11. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Fulda, S.; Vucic, D. Targeting IAP proteins for therapeutic intervention in cancer. Nat. Rev. Drug Discov. 2012, 11, 109–124. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Fan, T.; Yu, M. Inhibitor of apoptosis proteins and apoptosis. Acta Biochim. Biophys. Sin. 2008, 40, 278–288. [Google Scholar] [CrossRef] [PubMed]
- Eckelman, B.P.; Drag, M.; Snipas, S.J.; Salvesen, G.S. The mechanism of peptide-binding specificity of IAP BIR domains. Cell Death Differ. 2008, 15, 920–928. [Google Scholar] [CrossRef] [PubMed]
- Shiozaki, E.N.; Chai, J.; Rigotti, D.J.; Riedl, S.J.; Li, P.; Srinivasula, S.M.; Alnemri, E.S.; Fairman, R.; Shi, Y. Mechanism of XIAP-mediated inhibition of caspase-9. Mol. Cell 2003, 11, 519–527. [Google Scholar] [CrossRef]
- Lin, T.Y.; Chan, H.H.; Chen, S.H.; Sarvagalla, S.; Chen, P.S.; Coumar, M.S.; Cheng, S.M.; Chang, Y.C.; Lin, C.H.; Leung, E.; et al. BIRC5/Survivin is a novel ATG12-ATG5 conjugate interactor and an autophagy-induced DNA damage suppressor in human cancer and mouse embryonic fibroblast cells. Autophagy 2020, 16, 1296–1313. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Debatin, K.M. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 2006, 25, 4798–4811. [Google Scholar] [CrossRef] [PubMed]
- Gyrd-Hansen, M.; Meier, P. IAPs: From caspase inhibitors to modulators of NF-kappaB, inflammation and cancer. Nat. Rev. Cancer 2010, 10, 561–574. [Google Scholar] [CrossRef] [PubMed]
- Wertz, I.E.; Dixit, V.M. Regulation of death receptor signaling by the ubiquitin system. Cell Death Differ. 2010, 17, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Dogan, T.; Harms, G.S.; Hekman, M.; Karreman, C.; Oberoi, T.K.; Alnemri, E.S.; Rapp, U.R.; Rajalingam, K. X-linked and cellular IAPs modulate the stability of C-RAF kinase and cell motility. Nat. Cell Biol. 2008, 10, 1447–1455. [Google Scholar] [CrossRef] [PubMed]
- Uren, A.G.; Beilharz, T.; O’Connell, M.J.; Bugg, S.J.; van Driel, R.; Vaux, D.L.; Lithgow, T. Role for yeast inhibitor of apoptosis (IAP)-like proteins in cell division. Proc. Natl. Acad. Sci. USA 1999, 96, 10170–10175. [Google Scholar] [CrossRef]
- Ebner, P.; Poetsch, I.; Deszcz, L.; Hoffmann, T.; Zuber, J.; Ikeda, F. The IAP family member BRUCE regulates autophagosome-lysosome fusion. Nat. Commun. 2018, 9, 599. [Google Scholar] [CrossRef] [PubMed]
- Sauer, M.; Reiners, K.S.; Hansen, H.P.; Engert, A.; Gasser, S.; von Strandmann, E.P. Induction of the DNA damage response by IAP inhibition triggers natural immunity via upregulation of NKG2D ligands in Hodgkin lymphoma in vitro. Biol. Chem. 2013, 394, 1325–1331. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.M.; Chang, Y.C.; Liu, C.Y.; Lee, J.Y.; Chan, H.H.; Kuo, C.W.; Lin, K.Y.; Tsai, S.L.; Chen, S.H.; Li, C.F.; et al. YM155 down-regulates survivin and XIAP, modulates autophagy and induces autophagy-dependent DNA damage in breast cancer cells. Br. J. Pharmacol. 2015, 172, 214–234. [Google Scholar] [CrossRef] [PubMed]
- Ghobrial, I.M.; Witzig, T.E.; Adjei, A.A. Targeting apoptosis pathways in cancer therapy. CA Cancer J. Clin. 2005, 55, 178–194. [Google Scholar] [CrossRef] [PubMed]
- Mehrotra, S.; Languino, L.R.; Raskett, C.M.; Mercurio, A.M.; Dohi, T.; Altieri, D.C. IAP regulation of metastasis. Cancer Cell 2010, 17, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, D.; Luo, W.; Yu, Y.; Yu, J.; Li, J.; Zhang, X.; Zhang, B.; Chen, J.; Wu, X.R.; et al. X-linked inhibitor of apoptosis protein (XIAP) mediates cancer cell motility via Rho GDP dissociation inhibitor (RhoGDI)-dependent regulation of the cytoskeleton. J. Biol. Chem. 2011, 286, 15630–15640. [Google Scholar] [CrossRef]
- Tran, J.; Rak, J.; Sheehan, C.; Saibil, S.D.; LaCasse, E.; Korneluk, R.G.; Kerbel, R.S. Marked induction of the IAP family antiapoptotic proteins survivin and XIAP by VEGF in vascular endothelial cells. Biochem. Biophys. Res. Commun. 1999, 264, 781–788. [Google Scholar] [CrossRef]
- Rathore, R.; McCallum, J.E.; Varghese, E.; Florea, A.M.; Busselberg, D. Overcoming chemotherapy drug resistance by targeting inhibitors of apoptosis proteins (IAPs). Apoptosis 2017, 22, 898–919. [Google Scholar] [CrossRef]
- Ward, G.A.; Lewis, E.J.; Ahn, J.S.; Johnson, C.N.; Lyons, J.F.; Martins, V.; Munck, J.M.; Rich, S.J.; Smyth, T.; Thompson, N.T.; et al. ASTX660, a Novel Non-peptidomimetic Antagonist of cIAP1/2 and XIAP, Potently Induces TNFalpha-Dependent Apoptosis in Cancer Cell Lines and Inhibits Tumor Growth. Mol. Cancer Ther. 2018, 17, 1381–1391. [Google Scholar] [CrossRef]
- Ahn, K.S.; Sethi, G.; Aggarwal, B.B. Embelin, an inhibitor of X chromosome-linked inhibitor-of-apoptosis protein, blocks nuclear factor-kappaB (NF-kappaB) signaling pathway leading to suppression of NF-kappaB-regulated antiapoptotic and metastatic gene products. Mol. Pharmacol. 2007, 71, 209–219. [Google Scholar] [CrossRef]
- Nakahara, T.; Kita, A.; Yamanaka, K.; Mori, M.; Amino, N.; Takeuchi, M.; Tominaga, F.; Hatakeyama, S.; Kinoyama, I.; Matsuhisa, A.; et al. YM155, a novel small-molecule survivin suppressant, induces regression of established human hormone-refractory prostate tumor xenografts. Cancer Res. 2007, 67, 8014–8021. [Google Scholar] [CrossRef] [PubMed]
- Talbot, D.C.; Ranson, M.; Davies, J.; Lahn, M.; Callies, S.; Andre, V.; Kadam, S.; Burgess, M.; Slapak, C.; Olsen, A.L.; et al. Tumor survivin is downregulated by the antisense oligonucleotide LY2181308: A proof-of-concept, first-in-human dose study. Clin. Cancer Res. 2010, 16, 6150–6158. [Google Scholar] [CrossRef] [PubMed]
- Allensworth, J.L.; Sauer, S.J.; Lyerly, H.K.; Morse, M.A.; Devi, G.R. Smac mimetic Birinapant induces apoptosis and enhances TRAIL potency in inflammatory breast cancer cells in an IAP-dependent and TNF-alpha-independent mechanism. Breast Cancer Res. Treat. 2013, 137, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Houghton, P.J.; Kang, M.H.; Reynolds, C.P.; Morton, C.L.; Kolb, E.A.; Gorlick, R.; Keir, S.T.; Carol, H.; Lock, R.; Maris, J.M.; et al. Initial testing (stage 1) of LCL161, a SMAC mimetic, by the Pediatric Preclinical Testing Program. Pediatr. Blood Cancer 2012, 58, 636–639. [Google Scholar] [CrossRef] [PubMed]
- Zhen, M.C.; Wang, F.Q.; Wu, S.F.; Zhao, Y.L.; Liu, P.G.; Yin, Z.Y. Identification of mTOR as a primary resistance factor of the IAP antagonist AT406 in hepatocellular carcinoma cells. Oncotarget 2017, 8, 9466–9475. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Du, C.; Fang, M.; Li, Y.; Li, L.; Wang, X. Smac, a mitochondrial protein that promotes cytochrome c-dependent caspase activation by eliminating IAP inhibition. Cell 2000, 102, 33–42. [Google Scholar] [CrossRef]
- Verhagen, A.M.; Ekert, P.G.; Pakusch, M.; Silke, J.; Connolly, L.M.; Reid, G.E.; Moritz, R.L.; Simpson, R.J.; Vaux, D.L. Identification of DIABLO, a mammalian protein that promotes apoptosis by binding to and antagonizing IAP proteins. Cell 2000, 102, 43–53. [Google Scholar] [CrossRef]
- Kominsky, D.J.; Bickel, R.J.; Tyler, K.L. Reovirus-induced apoptosis requires mitochondrial release of Smac/DIABLO and involves reduction of cellular inhibitor of apoptosis protein levels. J. Virol. 2002, 76, 11414–11424. [Google Scholar] [CrossRef]
- Scott, F.L.; Denault, J.B.; Riedl, S.J.; Shin, H.; Renatus, M.; Salvesen, G.S. XIAP inhibits caspase-3 and -7 using two binding sites: Evolutionarily conserved mechanism of IAPs. EMBO J. 2005, 24, 645–655. [Google Scholar] [CrossRef]
- Darding, M.; Feltham, R.; Tenev, T.; Bianchi, K.; Benetatos, C.; Silke, J.; Meier, P. Molecular determinants of Smac mimetic induced degradation of cIAP1 and cIAP2. Cell Death Differ. 2011, 18, 1376–1386. [Google Scholar] [CrossRef]
- Yang, Q.H.; Du, C. Smac/DIABLO selectively reduces the levels of c-IAP1 and c-IAP2 but not that of XIAP and livin in HeLa cells. J. Biol. Chem. 2004, 279, 16963–16970. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Jin, Y.; Arend, L.J. Smac3, a novel Smac/DIABLO splicing variant, attenuates the stability and apoptosis-inhibiting activity of X-linked inhibitor of apoptosis protein. J. Biol. Chem. 2003, 278, 52660–52672. [Google Scholar] [CrossRef] [PubMed]
- Tenev, T.; Bianchi, K.; Darding, M.; Broemer, M.; Langlais, C.; Wallberg, F.; Zachariou, A.; Lopez, J.; MacFarlane, M.; Cain, K.; et al. The Ripoptosome, a signaling platform that assembles in response to genotoxic stress and loss of IAPs. Mol. Cell 2011, 43, 432–448. [Google Scholar] [CrossRef] [PubMed]
- Feoktistova, M.; Geserick, P.; Kellert, B.; Dimitrova, D.P.; Langlais, C.; Hupe, M.; Cain, K.; MacFarlane, M.; Hacker, G.; Leverkus, M. cIAPs block Ripoptosome formation, a RIP1/caspase-8 containing intracellular cell death complex differentially regulated by cFLIP isoforms. Mol. Cell 2011, 43, 449–463. [Google Scholar] [CrossRef]
- Akara-Amornthum, P.; Lomphithak, T.; Choksi, S.; Tohtong, R.; Jitkaew, S. Key necroptotic proteins are required for Smac mimetic-mediated sensitization of cholangiocarcinoma cells to TNF-alpha and chemotherapeutic gemcitabine-induced necroptosis. PLoS ONE 2020, 15, e0227454. [Google Scholar] [CrossRef] [PubMed]
- Mizutani, Y.; Nakanishi, H.; Yamamoto, K.; Li, Y.N.; Matsubara, H.; Mikami, K.; Okihara, K.; Kawauchi, A.; Bonavida, B.; Miki, T. Downregulation of Smac/DIABLO expression in renal cell carcinoma and its prognostic significance. J. Clin. Oncol. 2005, 23, 448–454. [Google Scholar] [CrossRef]
- Kempkensteffen, C.; Hinz, S.; Christoph, F.; Krause, H.; Magheli, A.; Schrader, M.; Schostak, M.; Miller, K.; Weikert, S. Expression levels of the mitochondrial IAP antagonists Smac/DIABLO and Omi/HtrA2 in clear-cell renal cell carcinomas and their prognostic value. J. Cancer Res. Clin. Oncol. 2008, 134, 543–550. [Google Scholar] [CrossRef]
- Endo, K.; Kohnoe, S.; Watanabe, A.; Tashiro, H.; Sakata, H.; Morita, M.; Kakeji, Y.; Maehara, Y. Clinical significance of Smac/DIABLO expression in colorectal cancer. Oncol. Rep. 2009, 21, 351–355. [Google Scholar] [CrossRef]
- Mizutani, Y.; Katsuoka, Y.; Bonavida, B. Prognostic significance of second mitochondria-derived activator of caspase (Smac/DIABLO) expression in bladder cancer and target for therapy. Int. J. Oncol. 2010, 37, 503–508. [Google Scholar] [CrossRef][Green Version]
- Mizutani, Y.; Katsuoka, Y.; Bonavida, B. Low circulating serum levels of second mitochondria-derived activator of caspase (Smac/DIABLO) in patients with bladder cancer. Int. J. Oncol. 2012, 40, 1246–1250. [Google Scholar] [CrossRef]
- Sekimura, A.; Konishi, A.; Mizuno, K.; Kobayashi, Y.; Sasaki, H.; Yano, M.; Fukai, I.; Fujii, Y. Expression of Smac/DIABLO is a novel prognostic marker in lung cancer. Oncol. Rep. 2004, 11, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Bao, S.T.; Gui, S.Q.; Lin, M.S. Relationship between expression of Smac and Survivin and apoptosis of primary hepatocellular carcinoma. Hepatobiliary Pancreat Dis. Int. 2006, 5, 580–583. [Google Scholar] [PubMed]
- Kempkensteffen, C.; Jager, T.; Bub, J.; Weikert, S.; Hinz, S.; Christoph, F.; Krause, H.; Schostak, M.; Miller, K.; Schrader, M. The equilibrium of XIAP and Smac/DIABLO expression is gradually deranged during the development and progression of testicular germ cell tumours. Int. J. Androl. 2007, 30, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Liao, Y.; Zhang, J.; Huang, Q.; Luo, W.; Yu, J.; Gong, J.; Zhou, Y.; Li, X.; Tang, B.; et al. Heat shock factor 1 inhibits the mitochondrial apoptosis pathway by regulating second mitochondria-derived activator of caspase to promote pancreatic tumorigenesis. J. Exp. Clin. Cancer Res. 2017, 36, 64. [Google Scholar] [CrossRef]
- Arellano-Llamas, A.; Garcia, F.J.; Perez, D.; Cantu, D.; Espinosa, M.; De la Garza, J.G.; Maldonado, V.; Melendez-Zajgla, J. High Smac/DIABLO expression is associated with early local recurrence of cervical cancer. BMC Cancer 2006, 6, 256. [Google Scholar] [CrossRef] [PubMed]
- Chai, J.; Du, C.; Wu, J.W.; Kyin, S.; Wang, X.; Shi, Y. Structural and biochemical basis of apoptotic activation by Smac/DIABLO. Nature 2000, 406, 855–862. [Google Scholar] [CrossRef]
- Weisberg, E.; Ray, A.; Barrett, R.; Nelson, E.; Christie, A.L.; Porter, D.; Straub, C.; Zawel, L.; Daley, J.F.; Lazo-Kallanian, S.; et al. Smac mimetics: Implications for enhancement of targeted therapies in leukemia. Leukemia 2010, 24, 2100–2109. [Google Scholar] [CrossRef]
- Cai, Q.; Sun, H.; Peng, Y.; Lu, J.; Nikolovska-Coleska, Z.; McEachern, D.; Liu, L.; Qiu, S.; Yang, C.Y.; Miller, R.; et al. A potent and orally active antagonist (SM-406/AT-406) of multiple inhibitor of apoptosis proteins (IAPs) in clinical development for cancer treatment. J. Med. Chem. 2011, 54, 2714–2726. [Google Scholar] [CrossRef]
- Flygare, J.A.; Beresini, M.; Budha, N.; Chan, H.; Chan, I.T.; Cheeti, S.; Cohen, F.; Deshayes, K.; Doerner, K.; Eckhardt, S.G.; et al. Discovery of a potent small-molecule antagonist of inhibitor of apoptosis (IAP) proteins and clinical candidate for the treatment of cancer (GDC-0152). J. Med. Chem. 2012, 55, 4101–4113. [Google Scholar] [CrossRef]
- Wong, H.; Gould, S.E.; Budha, N.; Darbonne, W.C.; Kadel, E.E., III; La, H.; Alicke, B.; Halladay, J.S.; Erickson, R.; Portera, C.; et al. Learning and confirming with preclinical studies: Modeling and simulation in the discovery of GDC-0917, an inhibitor of apoptosis proteins antagonist. Drug Metab. Dispos. 2013, 41, 2104–2113. [Google Scholar] [CrossRef]
- Varfolomeev, E.; Blankenship, J.W.; Wayson, S.M.; Fedorova, A.V.; Kayagaki, N.; Garg, P.; Zobel, K.; Dynek, J.N.; Elliott, L.O.; Wallweber, H.J.; et al. IAP antagonists induce autoubiquitination of c-IAPs, NF-kappaB activation, and TNFalpha-dependent apoptosis. Cell 2007, 131, 669–681. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Bai, L.; Sun, H.; Nikolovska-Coleska, Z.; McEachern, D.; Qiu, S.; Miller, R.S.; Yi, H.; Shangary, S.; Sun, Y.; et al. SM-164: A novel, bivalent Smac mimetic that induces apoptosis and tumor regression by concurrent removal of the blockade of cIAP-1/2 and XIAP. Cancer Res. 2008, 68, 9384–9393. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S. Promises and Challenges of Smac Mimetics as Cancer Therapeutics. Clin. Cancer Res. 2015, 21, 5030–5036. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, V.; Painuly, U.; Kimlinger, T.; Haug, J.; Rajkumar, S.V.; Kumar, S. Inhibitor of apoptosis proteins as therapeutic targets in multiple myeloma. Leukemia 2014, 28, 1519–1528. [Google Scholar] [CrossRef]
- Brands, R.C.; Herbst, F.; Hartmann, S.; Seher, A.; Linz, C.; Kubler, A.C.; Muller-Richter, U.D.A. Cytotoxic effects of SMAC-mimetic compound LCL161 in head and neck cancer cell lines. Clin. Oral. Investig. 2016, 20, 2325–2332. [Google Scholar] [CrossRef]
- Ren, K.; Ma, L.; Chong, D.; Zhang, Z.; Zhou, C.; Liu, H.; Zhao, S. Effects of LCL161, a Smac mimetic on the proliferation and apoptosis in hepatocellular carcinoma cells. Zhong Nan Da Xue Xue Bao Yi Xue Ban 2016, 41, 898–904. [Google Scholar]
- Shekhar, T.M.; Miles, M.A.; Gupte, A.; Taylor, S.; Tascone, B.; Walkley, C.R.; Hawkins, C.J. IAP antagonists sensitize murine osteosarcoma cells to killing by TNFalpha. Oncotarget 2016, 7, 33866–33886. [Google Scholar] [CrossRef]
- Jin, G.; Lan, Y.; Han, F.; Sun, Y.; Liu, Z.; Zhang, M.; Liu, X.; Zhang, X.; Hu, J.; Liu, H.; et al. Smac mimeticinduced caspaseindependent necroptosis requires RIP1 in breast cancer. Mol. Med. Rep. 2016, 13, 359–366. [Google Scholar] [CrossRef]
- Jin, G.; Liu, Y.; Xu, P.; Jin, G. Induction of Necroptosis in Human Breast Cancer Drug-Resistant Cells by SMAC Analog LCL161 After Caspase Inhibition Requires RIP3. Pharmazie 2019, 74, 363–368. [Google Scholar]
- Chen, S.M.; Lin, T.K.; Tseng, Y.Y.; Tu, C.H.; Lui, T.N.; Huang, S.F.; Hsieh, L.L.; Li, Y.Y. Targeting inhibitors of apoptosis proteins suppresses medulloblastoma cell proliferation via G2/M phase arrest and attenuated neddylation of p21. Cancer Med. 2018, 7, 3988–4003. [Google Scholar] [CrossRef]
- Langemann, D.; Trochimiuk, M.; Appl, B.; Hundsdoerfer, P.; Reinshagen, K.; Eschenburg, G. Sensitization of neuroblastoma for vincristine-induced apoptosis by Smac mimetic LCL161 is attended by G2 cell cycle arrest but is independent of NFkappaB, RIP1 and TNF-alpha. Oncotarget 2017, 8, 87763–87772. [Google Scholar] [CrossRef] [PubMed]
- Dougan, M.; Dougan, S.; Slisz, J.; Firestone, B.; Vanneman, M.; Draganov, D.; Goyal, G.; Li, W.; Neuberg, D.; Blumberg, R.; et al. IAP inhibitors enhance co-stimulation to promote tumor immunity. J. Exp. Med. 2010, 207, 2195–2206. [Google Scholar] [CrossRef] [PubMed]
- Knights, A.J.; Fucikova, J.; Pasam, A.; Koernig, S.; Cebon, J. Inhibitor of apoptosis protein (IAP) antagonists demonstrate divergent immunomodulatory properties in human immune subsets with implications for combination therapy. Cancer Immunol. Immunother. 2013, 62, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Muller-Sienerth, N.; Dietz, L.; Holtz, P.; Kapp, M.; Grigoleit, G.U.; Schmuck, C.; Wajant, H.; Siegmund, D. SMAC mimetic BV6 induces cell death in monocytes and maturation of monocyte-derived dendritic cells. PLoS ONE 2011, 6, e21556. [Google Scholar] [CrossRef] [PubMed]
- Chesi, M.; Mirza, N.N.; Garbitt, V.M.; Sharik, M.E.; Dueck, A.C.; Asmann, Y.W.; Akhmetzyanova, I.; Kosiorek, H.E.; Calcinotto, A.; Riggs, D.L.; et al. IAP antagonists induce anti-tumor immunity in multiple myeloma. Nat. Med. 2016, 22, 1411–1420. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Kondapuram, S.K.; Yang, T.H.; Syed, S.B.; Cheng, S.M.; Lin, T.Y.; Lin, Y.C.; Coumar, M.S.; Chang, J.Y.; Leung, E.; et al. The SMAC mimetic LCL161 is a direct ABCB1/MDR1-ATPase activity modulator and BIRC5/Survivin expression down-regulator in cancer cells. Toxicol. Appl. Pharmacol. 2020, 401, 115080. [Google Scholar] [CrossRef] [PubMed]
- Tian, A.; Wilson, G.S.; Lie, S.; Wu, G.; Hu, Z.; Hebbard, L.; Duan, W.; George, J.; Qiao, L. Synergistic effects of IAP inhibitor LCL161 and paclitaxel on hepatocellular carcinoma cells. Cancer Lett. 2014, 351, 232–241. [Google Scholar] [CrossRef]
- Scheurer, M.J.J.; Seher, A.; Steinacker, V.; Linz, C.; Hartmann, S.; Kubler, A.C.; Muller-Richter, U.D.A.; Brands, R.C. Targeting inhibitors of apoptosis in oral squamous cell carcinoma in vitro. J. Craniomaxillofac. Surg. 2019, 47, 1589–1599. [Google Scholar] [CrossRef]
- Najem, S.; Langemann, D.; Appl, B.; Trochimiuk, M.; Hundsdoerfer, P.; Reinshagen, K.; Eschenburg, G. Smac mimetic LCL161 supports neuroblastoma chemotherapy in a drug class-dependent manner and synergistically interacts with ALK inhibitor TAE684 in cells with ALK mutation F1174L. Oncotarget 2016, 7, 72634–72653. [Google Scholar] [CrossRef][Green Version]
- Ramakrishnan, V.; Gomez, M.; Prasad, V.; Kimlinger, T.; Painuly, U.; Mukhopadhyay, B.; Haug, J.; Bi, L.; Rajkumar, S.V.; Kumar, S. Smac mimetic LCL161 overcomes protective ER stress induced by obatoclax, synergistically causing cell death in multiple myeloma. Oncotarget 2016, 7, 56253–56265. [Google Scholar] [CrossRef]
- Yang, L.; Kumar, B.; Shen, C.; Zhao, S.; Blakaj, D.; Li, T.; Romito, M.; Teknos, T.N.; Williams, T.M. LCL161, a SMAC-mimetic, Preferentially Radiosensitizes Human Papillomavirus-negative Head and Neck Squamous Cell Carcinoma. Mol. Cancer Ther. 2019, 18, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Qin, Q.; Zuo, Y.; Yang, X.; Lu, J.; Zhan, L.; Xu, L.; Zhang, C.; Zhu, H.; Liu, J.; Liu, Z.; et al. Smac mimetic compound LCL161 sensitizes esophageal carcinoma cells to radiotherapy by inhibiting the expression of inhibitor of apoptosis protein. Tumour. Biol. 2014, 35, 2565–2574. [Google Scholar] [CrossRef] [PubMed]
- Hass, C.; Belz, K.; Schoeneberger, H.; Fulda, S. Sensitization of acute lymphoblastic leukemia cells for LCL161-induced cell death by targeting redox homeostasis. Biochem. Pharmacol. 2016, 105, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Infante, J.R.; Dees, E.C.; Olszanski, A.J.; Dhuria, S.V.; Sen, S.; Cameron, S.; Cohen, R.B. Phase I dose-escalation study of LCL161, an oral inhibitor of apoptosis proteins inhibitor, in patients with advanced solid tumors. J. Clin. Oncol. 2014, 32, 3103–3110. [Google Scholar] [CrossRef]
- Bardia, A.; Parton, M.; Kummel, S.; Estevez, L.G.; Huang, C.S.; Cortes, J.; Ruiz-Borrego, M.; Telli, M.L.; Martin-Martorell, P.; Lopez, R.; et al. Paclitaxel With Inhibitor of Apoptosis Antagonist, LCL161, for Localized Triple-Negative Breast Cancer, Prospectively Stratified by Gene Signature in a Biomarker-Driven Neoadjuvant Trial. J. Clin. Oncol. 2018, JCO2017748392. [Google Scholar] [CrossRef] [PubMed]
- Condon, S.M.; Mitsuuchi, Y.; Deng, Y.; LaPorte, M.G.; Rippin, S.R.; Haimowitz, T.; Alexander, M.D.; Kumar, P.T.; Hendi, M.S.; Lee, Y.H.; et al. Birinapant, a smac-mimetic with improved tolerability for the treatment of solid tumors and hematological malignancies. J. Med. Chem. 2014, 57, 3666–3677. [Google Scholar] [CrossRef] [PubMed]
- Benetatos, C.A.; Mitsuuchi, Y.; Burns, J.M.; Neiman, E.M.; Condon, S.M.; Yu, G.; Seipel, M.E.; Kapoor, G.S.; Laporte, M.G.; Rippin, S.R.; et al. Birinapant (TL32711), a bivalent SMAC mimetic, targets TRAF2-associated cIAPs, abrogates TNF-induced NF-kappaB activation, and is active in patient-derived xenograft models. Mol. Cancer Ther. 2014, 13, 867–879. [Google Scholar] [CrossRef]
- Po Yee Mak, D.H.M.; Ruvolo, V.; Jacamo, R.; Steven, M.K.; Andreeff, M.; Bing, Z. Carter, Apoptosis suppressor ARC modulates SMAC mimetic-induced cell death through BIRC2/MAP3K14 signalling in acute myeloid leukaemia. Br. Kournal Haematol. 2014, 163, 9. [Google Scholar]
- Eytan, D.F.; Snow, G.E.; Carlson, S.G.; Schiltz, S.; Chen, Z.; Van Waes, C. Combination effects of SMAC mimetic birinapant with TNFalpha, TRAIL, and docetaxel in preclinical models of HNSCC. Laryngoscope 2015, 125, E118–E124. [Google Scholar] [CrossRef]
- Krepler, C.; Chunduru, S.K.; Halloran, M.B.; He, X.; Xiao, M.; Vultur, A.; Villanueva, J.; Mitsuuchi, Y.; Neiman, E.M.; Benetatos, C.; et al. The novel SMAC mimetic birinapant exhibits potent activity against human melanoma cells. Clin. Cancer Res. 2013, 19, 1784–1794. [Google Scholar] [CrossRef]
- Ding, J.; Qin, D.; Zhang, Y.; Li, Q.; Li, Y.; Li, J. SMAC mimetic birinapant inhibits hepatocellular carcinoma growth by activating the cIAP1/TRAF3 signaling pathway. Mol. Med. Rep. 2020, 21, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, Z.; Tivnan, A.; Flanagan, L.; Murray, D.W.; Salvucci, M.; Stringer, B.W.; Day, B.W.; Boyd, A.W.; Kogel, D.; Rehm, M.; et al. Patient-derived glioblastoma cells show significant heterogeneity in treatment responses to the inhibitor-of-apoptosis-protein antagonist birinapant. Br. J. Cancer 2016, 114, 188–198. [Google Scholar] [CrossRef] [PubMed]
- McComb, S.; Aguade-Gorgorio, J.; Harder, L.; Marovca, B.; Cario, G.; Eckert, C.; Schrappe, M.; Stanulla, M.; von Stackelberg, A.; Bourquin, J.P.; et al. Activation of concurrent apoptosis and necroptosis by SMAC mimetics for the treatment of refractory and relapsed ALL. Sci. Transl. Med. 2016, 8, 339ra70. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.; Salvucci, M.; Hellwig, C.T.; Lincoln, F.A.; Mooney, R.E.; O’Connor, C.L.; Prehn, J.H.; Longley, D.B.; Rehm, M. Simulating and predicting cellular and in vivo responses of colon cancer to combined treatment with chemotherapy and IAP antagonist Birinapant/TL32711. Cell Death Differ. 2018, 25, 1952–1966. [Google Scholar] [CrossRef] [PubMed]
- Hamacher, R.; Schmid, R.M.; Saur, D.; Schneider, G. Apoptotic pathways in pancreatic ductal adenocarcinoma. Mol. Cancer 2008, 7, 64. [Google Scholar] [CrossRef] [PubMed]
- Carter, B.Z.; Mak, P.Y.; Mak, D.H.; Shi, Y.; Qiu, Y.; Bogenberger, J.M.; Mu, H.; Tibes, R.; Yao, H.; Coombes, K.R.; et al. Synergistic targeting of AML stem/progenitor cells with IAP antagonist birinapant and demethylating agents. J. Natl. Cancer Inst. 2014, 106, djt440. [Google Scholar] [CrossRef]
- Mak, D.Y.; Fraser, I.; Ferris, R.; James, K.; Liu, M.; Thomas, S.D.; McKenzie, M.; Lefresne, S. Comparison of Rapid to Standard Volumetric Modulated Arc Therapy for Palliative Radiotherapy in Lung Cancer Patients. Cureus 2020, 12, e10055. [Google Scholar] [CrossRef]
- Eytan, D.F.; Snow, G.E.; Carlson, S.; Derakhshan, A.; Saleh, A.; Schiltz, S.; Cheng, H.; Mohan, S.; Cornelius, S.; Coupar, J.; et al. SMAC Mimetic Birinapant plus Radiation Eradicates Human Head and Neck Cancers with Genomic Amplifications of Cell Death Genes FADD and BIRC2. Cancer Res. 2016, 76, 5442–5454. [Google Scholar] [CrossRef]
- Brumatti, G.; Ma, C.; Lalaoui, N.; Nguyen, N.Y.; Navarro, M.; Tanzer, M.C.; Richmond, J.; Ghisi, M.; Salmon, J.M.; Silke, N.; et al. The caspase-8 inhibitor emricasan combines with the SMAC mimetic birinapant to induce necroptosis and treat acute myeloid leukemia. Sci. Transl. Med. 2016, 8, 339ra69. [Google Scholar] [CrossRef]
- Kearney, C.J.; Lalaoui, N.; Freeman, A.J.; Ramsbottom, K.M.; Silke, J.; Oliaro, J. PD-L1 and IAPs co-operate to protect tumors from cytotoxic lymphocyte-derived TNF. Cell Death Differ. 2017, 24, 1705–1716. [Google Scholar] [CrossRef]
- Zhao, L.; Yang, G.; Bai, H.; Zhang, M.; Mou, D. NCTD promotes Birinapant-mediated anticancer activity in breast cancer cells by downregulation of c-FLIP. Oncotarget 2017, 8, 26886–26895. [Google Scholar] [CrossRef] [PubMed]
- Lalaoui, N.; Merino, D.; Giner, G.; Vaillant, F.; Chau, D.; Liu, L.; Kratina, T.; Pal, B.; Whittle, J.R.; Etemadi, N.; et al. Targeting triple-negative breast cancers with the Smac-mimetic birinapant. Cell Death Differ. 2020, 27, 2768–2780. [Google Scholar] [CrossRef] [PubMed]
- Orozco, C.A.; Martinez-Bosch, N.; Guerrero, P.E.; Vinaixa, J.; Dalotto-Moreno, T.; Iglesias, M.; Moreno, M.; Djurec, M.; Poirier, F.; Gabius, H.J.; et al. Targeting galectin-1 inhibits pancreatic cancer progression by modulating tumor-stroma crosstalk. Proc. Natl. Acad. Sci. USA 2018, 115, E3769–E3778. [Google Scholar] [CrossRef] [PubMed]
- Brands, R.C.; Scheurer, M.J.J.; Hartmann, S.; Seher, A.; Kubler, A.C.; Muller-Richter, U.D.A. Apoptosis-sensitizing activity of birinapant in head and neck squamous cell carcinoma cell lines. Oncol. Lett. 2018, 15, 4010–4016. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Shen, X.; Qu, J.; Straubinger, R.M.; Jusko, W.J. Proteomic Analysis of Combined Gemcitabine and Birinapant in Pancreatic Cancer Cells. Front. Pharmacol. 2018, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Straubinger, R.M.; Jusko, W.J. Mechanism-based mathematical modeling of combined gemcitabine and birinapant in pancreatic cancer cells. J. Pharmacokinet. Pharmacodyn. 2015, 42, 477–496. [Google Scholar] [CrossRef]
- Cekay, M.J.; Roesler, S.; Frank, T.; Knuth, A.K.; Eckhardt, I.; Fulda, S. Smac mimetics and type II interferon synergistically induce necroptosis in various cancer cell lines. Cancer Lett. 2017, 410, 228–237. [Google Scholar] [CrossRef]
- Michie, J.; Beavis, P.A.; Freeman, A.J.; Vervoort, S.J.; Ramsbottom, K.M.; Narasimhan, V.; Lelliott, E.J.; Lalaoui, N.; Ramsay, R.G.; Johnstone, R.W.; et al. Antagonism of IAPs Enhances CAR T-cell Efficacy. Cancer Immunol. Res. 2019, 7, 183–192. [Google Scholar] [CrossRef]
- Amaravadi, R.K.; Schilder, R.J.; Martin, L.P.; Levin, M.; Graham, M.A.; Weng, D.E.; Adjei, A.A. A Phase I Study of the SMAC-Mimetic Birinapant in Adults with Refractory Solid Tumors or Lymphoma. Mol. Cancer Ther. 2015, 14, 2569–2575. [Google Scholar] [CrossRef]
- Senzer, N.N.; LoRusso, P.; Martin, L.P.; Schilder, R.J.; Amaravadi, R.K.; Papadopoulos, K.P.; Segota, Z.E.; Weng, D.E.; Graham, M.; Adjei, A.A. Phase II clinical activity and tolerability of the SMAC-mimetic birinapant (TL32711) plus irinotecan in irinotecan-relapsed/refractory metastatic colorectal cancer. J. Clin. Oncol. 2013, 31 (Suppl. 15), 3621. [Google Scholar] [CrossRef]
- Hu, R.; Li, J.; Liu, Z.; Miao, M.; Yao, K. GDC-0152 induces apoptosis through down-regulation of IAPs in human leukemia cells and inhibition of PI3K/Akt signaling pathway. Tumour Biol. 2015, 36, 577–584. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Shu, T.; Liang, Y.; Gu, W.; Wang, C.; Song, X.; Fan, C.; Wang, W. GDC-0152 attenuates the malignant progression of osteosarcoma promoted by ANGPTL2 via PI3K/AKT but not p38MAPK signaling pathway. Int. J. Oncol. 2015, 46, 1651–1658. [Google Scholar] [CrossRef] [PubMed]
- Shekhar, T.M.; Burvenich, I.J.G.; Harris, M.A.; Rigopoulos, A.; Zanker, D.; Spurling, A.; Parker, B.S.; Walkley, C.R.; Scott, A.M.; Hawkins, C.J. Smac mimetics LCL161 and GDC-0152 inhibit osteosarcoma growth and metastasis in mice. BMC Cancer 2019, 19, 924. [Google Scholar] [CrossRef] [PubMed]
- Tchoghandjian, A.; Souberan, A.; Tabouret, E.; Colin, C.; Denicolai, E.; Jiguet-Jiglaire, C.; El-Battari, A.; Villard, C.; Baeza-Kallee, N.; Figarella-Branger, D. Inhibitor of apoptosis protein expression in glioblastomas and their in vitro and in vivo targeting by SMAC mimetic GDC-0152. Cell Death Dis. 2016, 7, e2325. [Google Scholar] [CrossRef]
- Mantik, P.; Xie, M.; Wong, H.; La, H.; Steigerwalt, R.W.; Devanaboyina, U.; Ganem, G.; Shih, D.; Flygare, J.A.; Fairbrother, W.J.; et al. Cyclodextrin Reduces Intravenous Toxicity of a Model Compound. J. Pharm. Sci. 2019, 108, 1934–1943. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.G.; Jones, S.A.; Murakami, S.C.; Budha, N.; Ware, J.; Wong, H.; Buonarati, M.H.; Dean, B.; Hop, C.E. Validation and application of a liquid chromatography-tandem mass spectrometric method for the determination of GDC-0152 in human plasma using solid-phase extraction. Biomed. Chromatogr. 2013, 27, 102–110. [Google Scholar] [CrossRef]
- Nikkhoo, A.; Rostami, N.; Farhadi, S.; Esmaily, M.; Moghadaszadeh Ardebili, S.; Atyabi, F.; Baghaei, M.; Haghnavaz, N.; Yousefi, M.; Aliparasti, M.R.; et al. Codelivery of STAT3 siRNA and BV6 by carboxymethyl dextran trimethyl chitosan nanoparticles suppresses cancer cell progression. Int. J. Pharm. 2020, 581, 119236. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| ClinicalTrials.Gov Identifier | Phase | Condition or Disease (in Patients) | Combination Therapy |
|---|---|---|---|
| NCT01968915 | Phase 1 (completed) | Neoplasms | - |
| NCT01955434 | Phase 2 (completed) | Recurrent plasma cell myeloma Refractory plasma cell myeloma | Cyclophophamide |
| NCT01934634 | Phase 1 | Metastatic pancreatic cancer | Gemcitabine Nab-Paclitaxel |
| NCT02098161 | Phase 2 | Polycythemia vera, post-polycythemic myelofibrosis phase Primary myelofibrosis Secondary myelofibrosis | - |
| NCT02649673 | Phase 1/2 | Small cell lung cancer Ovarian cancer | Topotecan Pegylated granulocyte colony stimulating factor |
| NCT01617668 | Phase 2 (completed) | Breast cancer | Paclitaxel |
| NCT01240655 | Phase 1 (completed) | Solid tumors | Paclitaxel |
| NCT01098838 | Phase 1 (completed) | Advanced solid tumors (lung, skin, colon, pancreas, and others) | - |
| NCT03111992 | Phase 1 (completed) | Multiple myeloma | PDR001 CJM112 |
| NCT02890069 | Phase 1 | Colorectal cancer Non-small cell lung carcinoma (Adenocarcinoma) Triple-negative breast cancer Renal cell carcinoma | PDR001 Evrolimus Panobinostat QBM076 HDM201 |
| ClinicalTrials.Gov Identifier | Phase | Condition or Disease (in Patients) | Combination Therapy |
|---|---|---|---|
| NCT03803774 | Phase 1 | Head and neck squamous cell carcinoma | Radiation therapy |
| NCT02587962 | Phase 1/2 (terminated) | Solid tumor | Pembrolizumab |
| NCT02147873 | Phase 2 (terminated) | Myelodysplastic syndrome Chronic myelomonocytic leukemia | Azacitidine Placebo |
| NCT01940172 | Phase 1 (completed) | Relapsed epithelial ovarian cancer Relapsed primary peritoneal cancer Relapsed fallopian tube cancer | Conatumumab |
| NCT01188499 | Phase 1/2 (completed) | Cancer | Carboplatin Paclitaxel Irinotecan Docetaxel Gemcitabine Liposomal Doxorubicin |
| NCT01681368 | Phase 2 (terminated) | Epithelial ovarian cancer Peritoneal neoplasms Fallopian tube neoplasms | - |
| NCT00993239 | Phase 1 (completed) | Cancer | - |
| NCT01573780 | Phase 1 (terminated) | Adult solid tumor | Gemcitabine |
| NCT01486784 | Phase 1/2 (terminated) | Acute myelogenous leukemia | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.-C.; Cheung, C.H.A. An Updated Review of Smac Mimetics, LCL161, Birinapant, and GDC-0152 in Cancer Treatment. Appl. Sci. 2021, 11, 335. https://doi.org/10.3390/app11010335
Chang Y-C, Cheung CHA. An Updated Review of Smac Mimetics, LCL161, Birinapant, and GDC-0152 in Cancer Treatment. Applied Sciences. 2021; 11(1):335. https://doi.org/10.3390/app11010335
Chicago/Turabian StyleChang, Yung-Chieh, and Chun Hei Antonio Cheung. 2021. "An Updated Review of Smac Mimetics, LCL161, Birinapant, and GDC-0152 in Cancer Treatment" Applied Sciences 11, no. 1: 335. https://doi.org/10.3390/app11010335
APA StyleChang, Y.-C., & Cheung, C. H. A. (2021). An Updated Review of Smac Mimetics, LCL161, Birinapant, and GDC-0152 in Cancer Treatment. Applied Sciences, 11(1), 335. https://doi.org/10.3390/app11010335