Trichoderma: Evaluation of Its Degrading Abilities for the Bioremediation of Hydrocarbon Complex Mixtures

 , ,
, ,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Used Engine Oil
2.2. Biological Material
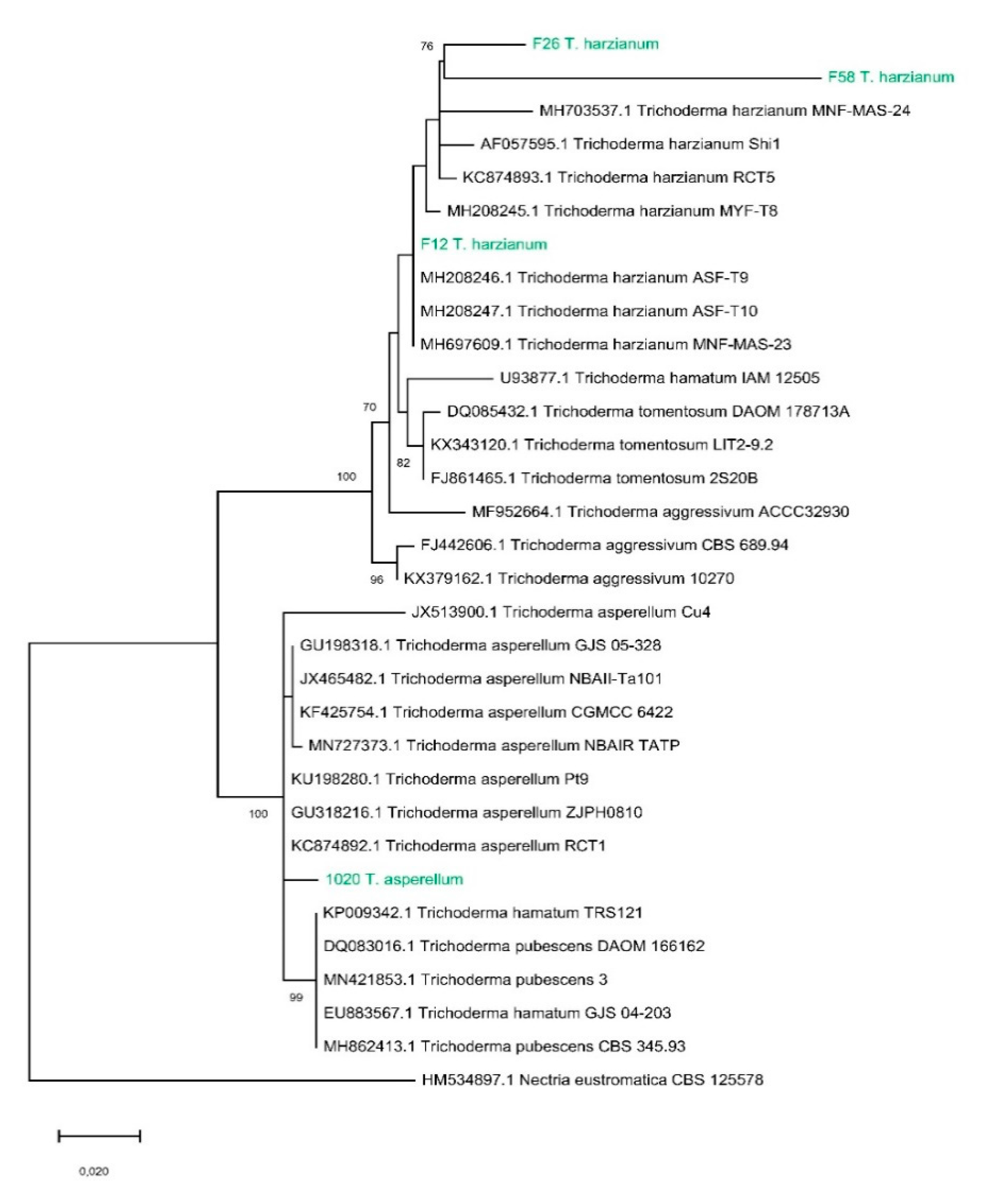
Trichoderma Strains and Their Phylogenetic Relationships
2.3. Engine Oil Tolerance Test on the Fungal Strains
2.4. Analysis of Used Engine Oil Degradation
2.5. Colorimetric Screening for the Detection of Ligninolytic Enzymatic Activities
2.6. Statistical Analysis
3. Results
3.1. Identification of Trichoderma Strains and Analysis of Their Phylogenetic Relationships
3.2. Used Engine Oil Tolerance Test on Trichoderma Strains
3.3. Used Engine Oil Composition
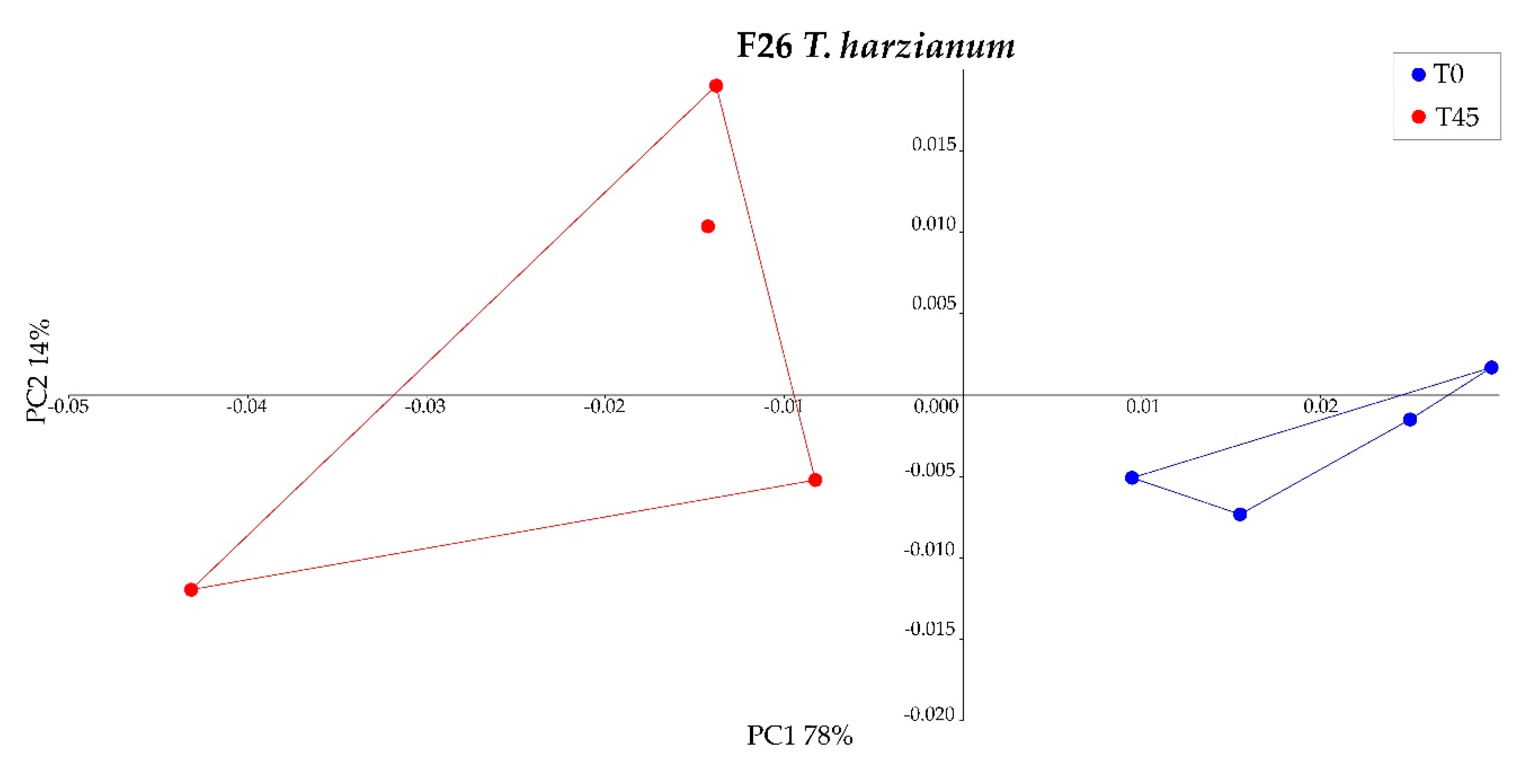
3.4. Analysis of Used Engine Oil Fungal Degradation
3.5. Results of the Colorimetric Screening for the Detection of Fungal Enzymatic Activities
4. Discussion
- The efficiency of degradative activities seems to vary greatly depending on different species and single strains.
- The use of strains isolated from contaminated environments provides an advantage in bioremediation activities. This may depend on the adaptation of fungi to stress conditions that limit their growth and on improved enzymatic response to the degradation of recalcitrant substances.
- T. harzianum F26 should be subject to further studies, including field studies, to deepen and enhance its potential as a bioremediation agent.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chaîneau, C.H.; Rougeux, G.; Yéprémian, C.; Oudot, J. Effects of nutrient concentration on the biodegradation of crude oil and associated microbial populations in the soil. Soil Biol. Biochem. 2005, 37, 1490–1497. [Google Scholar] [CrossRef]
- Wang, Z.; Fingas, M.; Blenkinsopp, S.; Sergy, G.; Landriault, M. Comparison of oil composition changes due to biodegradation and physical weathering in different oils. J. Chromatogr. A 1998, 809, 89–107. [Google Scholar] [CrossRef]
- Edewor, T.I.; Adelowo, O.O.; Afolabi, T.J. Preliminary studies into the biological activities of a broad spectrum disinfectant formulated from used engine oil. Pollut. Res. 2004, 234, 581–586. [Google Scholar]
- Wu, Y.; Lou, Y.; Zou, D.; Ni, J.; Liu, W.; Teng, Y.; Li, Z. Bioremediation of polycyclic aromatic hydrocarbons contaminated soil with Molilinia sp.: Degradation and microbial community analysis. Biodegradation 2008, 19, 247–257. [Google Scholar] [CrossRef]
- Mackay, D.; Fraser, A. Bioaccumulation of persistent organic chemicals: Mechanisms and models. Environ. Pollut. 2000, 111, 375–391. [Google Scholar] [CrossRef]
- Lloyd, A.C.; Cackette, T.A. Diesel engines: Environmental impact and control. J. Air Waste Manag. Assoc. 2001, 51, 809–847. [Google Scholar] [CrossRef]
- Mishra, S.; Jyot, J.; Kuhad, R.C.; Lal, B. Evaluation of inoculum addition to stimulate in situ bioremediation of oily-sludge-contaminated soil. Appl. Environ. Microbiol. 2001, 67, 1675–1681. [Google Scholar] [CrossRef]
- Amadi, A.; Abbey, S.D.; Nma, A. Chronic effects of oil spill on soil properties and microflora of a rainforest ecosystem in Nigeria. Water. Air Soil Pollut. 1996, 86, 1–11. [Google Scholar] [CrossRef]
- Asemoloye, M.D.; Ahmad, R.; Jonathan, S.G. Synergistic action of rhizospheric fungi with Megathyrsus maximus root speeds up hydrocarbon degradation kinetics in oil polluted soil. Chemosphere 2017, 187, 1–10. [Google Scholar] [CrossRef]
- Spini, G.; Spina, F.; Poli, A.; Blieux, A.L.; Regnier, T.; Gramellini, C.; Varese, G.C.; Puglisi, E. Molecular and microbiological insights on the enrichment procedures for the isolation of petroleum degrading bacteria and fungi. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Daccò, C.; Girometta, C.; Asemoloye, M.D.; Carpani, G.; Picco, A.M.; Tosi, S. Key fungal degradation patterns, enzymes and their applications for the removal of aliphatic hydrocarbons in polluted soils: A review. Int. Biodeterior. Biodegrad. 2020, 147. [Google Scholar] [CrossRef]
- Cerniglia, C.E.; Sutherland, G.R. Degradation of polycyclic aromatic hydrocarbons by fungi. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin, Germany, 2010; pp. 2079–2110. [Google Scholar]
- Magan, N. Fungi in extreme environment. In The Mycota Environmental and Microbial Relationships; Esser, K., Lemke, P.A., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 2007; Volume IV, pp. 99–114. [Google Scholar]
- Deshmukh, R.; Khardenavis, A.A.; Purohit, H.J. Diverse metabolic capacities of fungi for bioremediation. Indian J. Microbiol. 2016, 56, 247–264. [Google Scholar] [CrossRef] [PubMed]
- Lieckfeldt, E.; Samuels, G.J.; Nirenberg, H.; Petrini, O. A morphological and molecular perspective of Trichoderma viride: Is it one or two species? Appl. Environ. Microbiol. 1999, 65, 2418–2428. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Druzhinina, I.; Kubick, C.P.; Xu, T. Trichoderma biodiversity in China: Evidence for a north to southern distribution of species in East Asia. FEMS Microbiol. Lett. 2005, 251, 251–257. [Google Scholar] [CrossRef]
- Ezzi, M.I.; Lynch, J.M. Biodegradation of cyanide by Trichoderma spp. and Fusarium spp. Enzyme Microb. Technol. 2005, 36, 849–854. [Google Scholar] [CrossRef]
- Tripathi, P.; Singh, P.C.; Mishra, A.; Chauhan, P.S.; Dwivedi, S.; Bais, R.T.; Tripathi, R.D. Trichoderma: A potential bioremediator for environmental cleanup. Clean Technol. Environ. Policy 2013, 15, 541–550. [Google Scholar] [CrossRef]
- Ávila-Miranda, M.E.; Herrera-Estrella, A.; Peña-Cabriales, J.J. Colonization of the rhizosphere, rhizoplane and endorhiza of garlic (Allium sativum L.) by strains of Trichoderma harzianum and their capacity to control allium white-rot under field conditions. Soil Biol. Biochem. 2006, 38, 1823–1830. [Google Scholar] [CrossRef]
- Rojo, F.G.; Reynoso, M.M.; Ferez, M.; Chulze, S.N.; Torres, A.M. Biological control by Trichoderma species of Fusarium solani causing peanut brown root rot under field conditions. Crop Prot. 2007, 26, 549–555. [Google Scholar] [CrossRef]
- Chaîneau, C.H.; Morel, J.; Dupont, J.; Bury, E.; Oudot, J. Comparison of the fuel oil biodegradation potential of hydrocarbon-assimilating microorganisms isolated from a temperate agricultural soil. Sci. Total Environ. 1999, 227, 237–247. [Google Scholar] [CrossRef]
- Ravelet, C.; Krivobok, S.; Sage, L.; Steiman, R. Biodegradation of pyrene by sediment fungi. Chemosphere 2000, 40, 557–563. [Google Scholar] [CrossRef]
- Saraswathy, A.; Hallberg, R. Degradation of pyrene by indigenous fungi from a former gasworks site. FEMS Microbiol. Lett. 2002, 210, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Hadibarata, T.; Tachibana, S. Microbial degradation of n-eicosane by filamentous fungi. In Interdisciplinary Studies on Environmental Chemistry e Environmental Research in Asia; Obayashi, Y., Isobe, T., Subramanian, A., Suzuki, S., Tanabe, S., Eds.; Terrapub: Tokyo, Japan, 2009; pp. 323–329. [Google Scholar]
- Atagana, H.I. Biodegradation of PAHs by fungi in contaminated soil containing cadmium and nickel ions. Afr. J. Biotechnol. 2009, 21, 5780–5789. [Google Scholar] [CrossRef]
- Zafra, G.; Moreno-Montano, A.; Absalon, A.E.; Cortes-Espinosa, D.V. Degradation of polycyclic aromatic hydrocarbons in soil by a tolerant strain of Trichoderma asperellum. Environ. Sci. Pollut. Res. 2015, 22, 1034–1042. [Google Scholar] [CrossRef] [PubMed]
- Argumedo-Delira, R.; Alarcon, A.; Ferrera-Cerrato, R.; Almaraz, J.J.; Pena-Cabriales, J.J. Tolerance and growth of 11 Trichoderma strains to crude oil, naphthalene, phenanthrene and benzo[a]pyrene. J. Environ. Manag. 2012, 95, S291–S299. [Google Scholar] [CrossRef] [PubMed]
- Cazares-Garcia, S.V.; Vazquez-Garciduenas, M.S.; Vazquez-Marrufo, G. Structural and phylogenetic analysis of laccases from Trichoderma: A bioinformatic approach. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Cristica, M.; Manoliu, A.; Barbaneagra, T.; Ciornea, E. Compared analysis of catalase and peroxidase activity in cellulolytic fungus Trichoderma reesei grown on medium with different concentrations of grinded wheat and barley straws. Sci. Ann. Alexandru Ioan Cuza Univ. Iaşi N. Ser. Sect. IIA Genet. Mol. Biol. 2011, 12, 89. [Google Scholar]
- Hadibarata, T.; Tachibana, S.; Itoh, K. Biodegradation of phenanthrene by fungi screened from nature. Pak. J. Biol. Sci. 2007, 10, 2535–2543. [Google Scholar] [CrossRef]
- Marchand, C.; St-Arnaud, M.; Hogland, W.; Bell, T.H.; Hijri, M. Petroleum biodegradation capacity of bacteria and fungi isolated from petroleum-contaminated soil. Int. Biodeterior. Biodegrad. 2017, 116, 48–57. [Google Scholar] [CrossRef]
- Ameen, F.; Moslem, M.; Hadi, S.; Al-Sabri, A.E. Biodegradation of diesel fuel hydrocarbons by mangrove fungi from Red Sea Coast of Saudi Arabia. Saudi J. Biol. Sci. 2016, 23, 211–218. [Google Scholar] [CrossRef]
- Yao, L.; Teng, Y.; Luo, Y.; Christie, P.; Ma, W.; Liu, F.; Wu, Y.; Luo, Y.; Li, Z. Biodegradation of polycyclic aromatic hydrocarbons (PAHs) by Trichoderma reesei FS10-C and effect of bioaugmentation on an aged PAH-contaminated soil. Bioremediat. J. 2015, 19, 9–17. [Google Scholar] [CrossRef]
- Martin, K.J.; Rygiewicz, P.T. Fungal-specific PCR primers developed for analysis of the ITS region of environmental DNA extracts. BMC Microbiol. 2005, 5. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Gams, W.; Bissett, J. Morphology and identification of Trichoderma. In Trichoderma and Gliocladium. Basic Biology, Taxonomy and Genetics; Kubicek, C.P., Harman, G.E., Eds.; Taylor & Francis: London, UK, 1998; Volume 1, pp. 3–34. [Google Scholar]
- Scharf, M. Termites as targets and models for biotechnology. Ann. Rev. Entomol. 2015, 60, 77–102. [Google Scholar] [CrossRef] [PubMed]
- Yanto, D.H.Y.; Tachibana, S. Biodegradation of petroleum hydrocarbons by a newly isolated Pestalotiopsis sp. NG007. Int. Biodeterior. Biodegrad. 2013, 85, 438–450. [Google Scholar] [CrossRef]
- Shleev, S.V.; Morozova, O.V.; Nikitina, O.V.; Gorshina, E.S.; Rusinova, T.V.; Serezhenkov, V.A.; Burbaev, D.S.; Gazaryan, I.G.; Yaropolov, A.I. Comparison of physico-chemical characteristics of four laccases from different basidiomycetes. Biochimie 2004, 86, 693–703. [Google Scholar] [CrossRef]
- Lee, H.; Jang, Y.; Choi, Y.S.; Kim, M.J.; Lee, J.; Lee, H.; Hong, J.H.; Min Lee, Y.; Kim, G.H.; Kim, J.J. Biotechnological procedures to select white rot fungi for the degradation of PAHs. J. Microbiol. Methods 2014, 97, 56–62. [Google Scholar] [CrossRef]
- Kumar, V.V.; Rapheal, V.S. Induction and purification by three-phase partitioning of aryl alcohol oxidase (AAO) from Pleurotus ostreatus. Appl. Biochem. Biotechnol. 2011, 163, 423–432. [Google Scholar] [CrossRef]
- Hammel, K.E. Oxidation of aromatic pollutants by lignin-degrading fungi and their extracellular peroxidases. In Metal Ions in Biological Systems; Sigel, H., Sigel, A., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1992; pp. 41–60. [Google Scholar]
- Batista-García, R.A.; Kumar, V.V.; Ariste, A.; Tovar-Herrera, O.E.; Savary, O.; Peidro-Guzmán, H.; González-Abradelo, D.; Jackson, S.A.; Dobson, A.D.W.; Sánchez-Carbente, M.d.R.; et al. Simple screening protocol for identification of potential mycoremediation tools for the elimination of polycyclic aromatic hydrocarbons and phenols from hyperalkalophile industrial effluents. J. Environ. Manag. 2017, 198, 1–11. [Google Scholar] [CrossRef]
- Lee, H.; Choi, Y.S.; Kim, M.J.; Huh, N.Y.; Kim, G.H.; Lim, Y.W.; Kang, S.M.; Cho, S.T.; Kim, J.J. Degrading ability of oligocyclic aromates by Phanerchaete sordida selected via screening of white rot fungi. Folia Microbiol. 2010, 55, 447–453. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica 2001, 4, 9. [Google Scholar]
- Hostettler, F.D.; Lorenson, T.D.; Bekins, B.A. Petroleum fingerprinting with organic markers. Environ. Forensics 2013, 14, 262–277. [Google Scholar] [CrossRef]
- Zafra, G.; Cortés-Espinosa, D.V. Biodegradation of polycyclic aromatic hydrocarbons by Trichoderma species: A mini review. Environ. Sci. Pollut. Res. 2015, 22, 19426–19433. [Google Scholar] [CrossRef] [PubMed]
- Silva, I.S.; Grossman, M.; Durrant, L.R. Degradation of polycyclic aromatic hydrocarbons (2–7 rings) under microaerobic and very-low-oxygen conditions by soil fungi. Int. Biodeterior. Biodegrad. 2009, 63, 224–229. [Google Scholar] [CrossRef]
- Hamzah, A.; Zarin, M.A.; Hamid, A.A.; Omar, O.; Senafi, S. Optimal physical and nutrient parameters for growth of Trichoderma virens UKMP-1M for heavy crude oil degradation. Sains Malaysiana 2012, 41, 71–79. [Google Scholar]
- Cobas, M.; Ferreira, L.; Tavares, T.; Sanroman, M.A.; Pazos, M. Development of permeable reactive biobarrier for the removal of PAHs by Trichoderma longibrachiatum. Chemosphere 2013, 91, 711–716. [Google Scholar] [CrossRef]
- Husaini, A. Bioremediation of crude oil by different fungal genera. Asian J. Plant Biol. 2014, 2, 11–18. [Google Scholar]
- Husaini, A.; Roslan, H.A.; Hii, K.S.Y.; Ang, C.H. Biodegradation of aliphatic hydrocarbon by indigenous fungi isolated from used motor oil contaminated sites. World J. Microbiol. Biotechnol. 2008, 24, 2789–2797. [Google Scholar] [CrossRef]
- Matsubara, M.; Lynch, J.M.; De Leij, F.A.A.M. A simple screening procedure for selecting fungi with potential for use in the bioremediation of contaminated land. Enzyme Microb. Technol. 2006, 39, 1365–1372. [Google Scholar] [CrossRef]
- Romero, M.C.; Salvioli, M.L.; Cazau, M.C.; Arambarri, A.M. Pyrene degradation by yeasts and filamentous fungi. Environ. Pollut. 2002, 117, 159–163. [Google Scholar] [CrossRef]
- Kadri, T.; Rouissi, T.; Kaur Brar, S.; Cledon, M.; Sarma, S.; Verma, M. Biodegradation of polycyclic aromatic hydrocarbons (PAHs) by fungal enzymes: A review. J. Environ. Sci. 2017, 51, 52–74. [Google Scholar] [CrossRef]
- Prenafeta-Boldú, F.X.; de Hoog, G.S.; Summerbell, R.C. Fungal communities in hydrocarbon degradation. In Microbial Communities Utilizing Hydrocarbons and Lipids: Members, Metagenomics and Ecophysiology. Handbook of Hydrocarbon and Lipid Microbiology; McGenity, T., Ed.; Springer: Cham, Switzerland, 2019; pp. 1–36. [Google Scholar]
- Jonathan, S.G.; Asemoloye, M.; Ahmad, R.; Olawuyi, O.J.; Adejoye, D. Response of a newly identified Trichoderma harzianum ky488466 to crude oil pollution and its expression of peroxidase genes. SSRN 2017, 1–10. [Google Scholar] [CrossRef]
- Durairaj, P.; Malla, S.; Nadarajan, S.P.; Lee, P.G.; Jung, E.; Park, H.H.; Kim, B.G.; Yun, H. Fungal cytochrome P450 monooxygenases of Fusarium oxysporum for the synthesis of ω-hydroxy fatty acids in engineered Saccharomyces cerevisiae. Microb. Cell Fact. 2015, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Asemoloye, M.D.; Jonathan, S.G.; Ahmad, R. Synergistic plant-microbes interactions in the rhizosphere: A potential headway for the remediation of hydrocarbon polluted soils. Int. J. Phytoremediat. 2019, 2, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Janusz, G.; Kucharzyk, K.H.; Pawlik, A.; Staszczak, M.; Paszczynski, A.J. Fungal laccase, manganese peroxidase and lignin peroxidase: Gene expression and regulation. Enzyme Microb. Technol. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Yang, J.; Li, W.; Bun Ng, T.; Deng, X.; Lin, J.; Ye, X. Laccases: Production, expression regulation, and applications in pharmaceutical biodegradation. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Casieri, L.; Anastasi, A.; Prigione, V.; Varese, G.C. Survey of ectomycorrhizal, litter-degrading, and wood-degrading basidiomycetes for dye decolorization and ligninolytic enzyme activity. Int. J. Gen. Mol. Microbiol. 2010, 98, 483–504. [Google Scholar] [CrossRef]
- Jeon, J.R.; Baldrian, P.; Murugesan, K.; Chang, Y.S. Laccase-catalysed oxidations of naturally occurring phenols: From in vivo biosynthetic pathways to green synthetic applications. Microb. Biotechnol. 2012, 5, 318–332. [Google Scholar] [CrossRef]
- Liu, J.; Liu, W.; Cai, Y.; Liao, X. Laccase production by Trametes hirsuta, characterization, and its capability of decoloring chlorophyll. Pol. J. Microbiol. 2014, 63, 323–333. [Google Scholar] [CrossRef]
- Si, J.; Peng, F.; Cui, B. Purification, biochemical characterization and dye decolorization capacity of an alkali-resistant and metal-tolerant laccase from Trametes pubescens. Bioresour. Technol. 2013, 128, 49–57. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Species |
|---|---|
| F1020 | T. asperellum |
| F12 | T. harzianum |
| F26 | T. harzianum |
| F58 | T. harzianum |
| Increment in the Coverage Percentage of the Dish Compared to the Control | |
|---|---|
| T. asperellum F1020 | + |
| T. harzianum F12 | ++ |
| T. harzianum F58 | ++ |
| T. harzianum F26 | +++ |
| Guaiacol | Gallic Acid | RBBR | |
|---|---|---|---|
| T. asperellum F1020 | +++ | +++ | 0 |
| T. harzianum F12 | +++ | + | + |
| T. harzianum F58 | + | 0 | + |
| T. harzianum F26 | ++ | 0 | +++ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daccò, C.; Nicola, L.; Temporiti, M.E.E.; Mannucci, B.; Corana, F.; Carpani, G.; Tosi, S. Trichoderma: Evaluation of Its Degrading Abilities for the Bioremediation of Hydrocarbon Complex Mixtures. Appl. Sci. 2020, 10, 3152. https://doi.org/10.3390/app10093152
Daccò C, Nicola L, Temporiti MEE, Mannucci B, Corana F, Carpani G, Tosi S. Trichoderma: Evaluation of Its Degrading Abilities for the Bioremediation of Hydrocarbon Complex Mixtures. Applied Sciences. 2020; 10(9):3152. https://doi.org/10.3390/app10093152
Chicago/Turabian StyleDaccò, Chiara, Lidia Nicola, Marta Elisabetta Eleonora Temporiti, Barbara Mannucci, Federica Corana, Giovanna Carpani, and Solveig Tosi. 2020. "Trichoderma: Evaluation of Its Degrading Abilities for the Bioremediation of Hydrocarbon Complex Mixtures" Applied Sciences 10, no. 9: 3152. https://doi.org/10.3390/app10093152
APA StyleDaccò, C., Nicola, L., Temporiti, M. E. E., Mannucci, B., Corana, F., Carpani, G., & Tosi, S. (2020). Trichoderma: Evaluation of Its Degrading Abilities for the Bioremediation of Hydrocarbon Complex Mixtures. Applied Sciences, 10(9), 3152. https://doi.org/10.3390/app10093152