Anti-Inflammatory Flavonolignans from Triticum aestivum Linn. Hull

 , , ,
, , ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. General Information
2.2. Plant Material
2.3. Extract and Isolation
2.4. Measurement of NO Production on LPS-Stimulated RAW 264.7 Cells
2.5. Cytotoxicity Assay
2.6. Statistical Analysis
3. Results
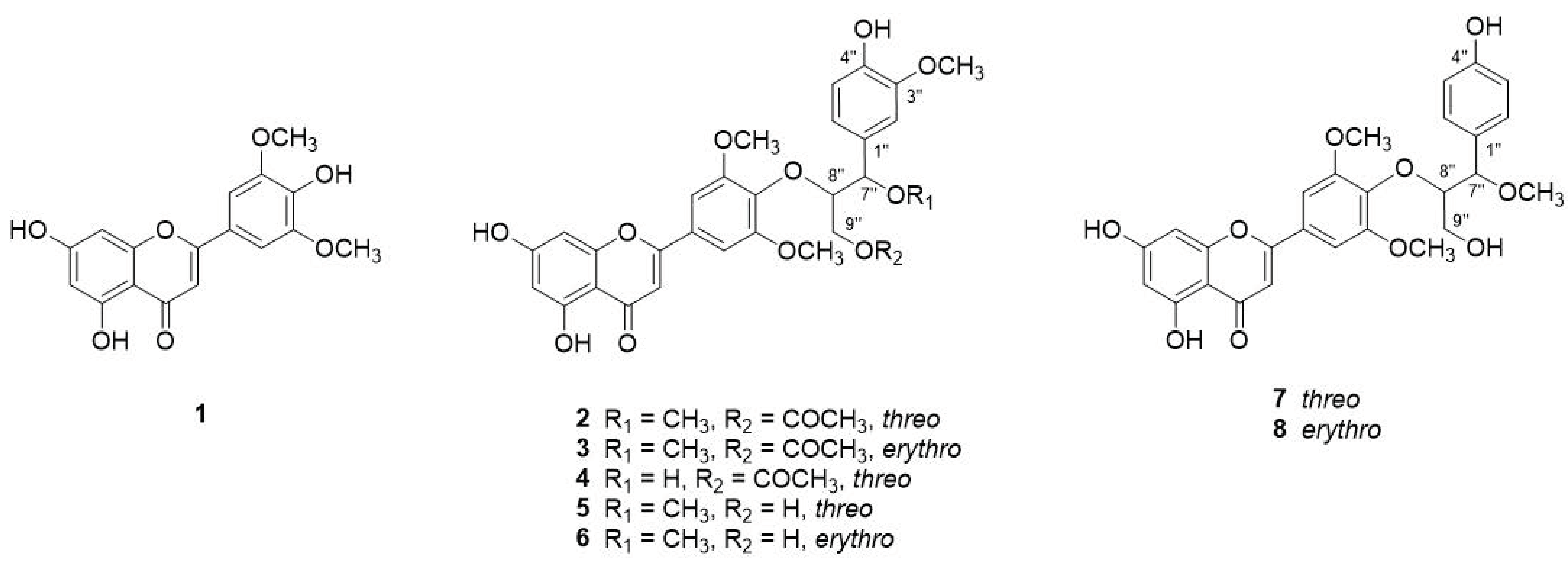
3.1. Structure Identification of the Isolated Compounds
3.2. Chemotaxonomic Significance
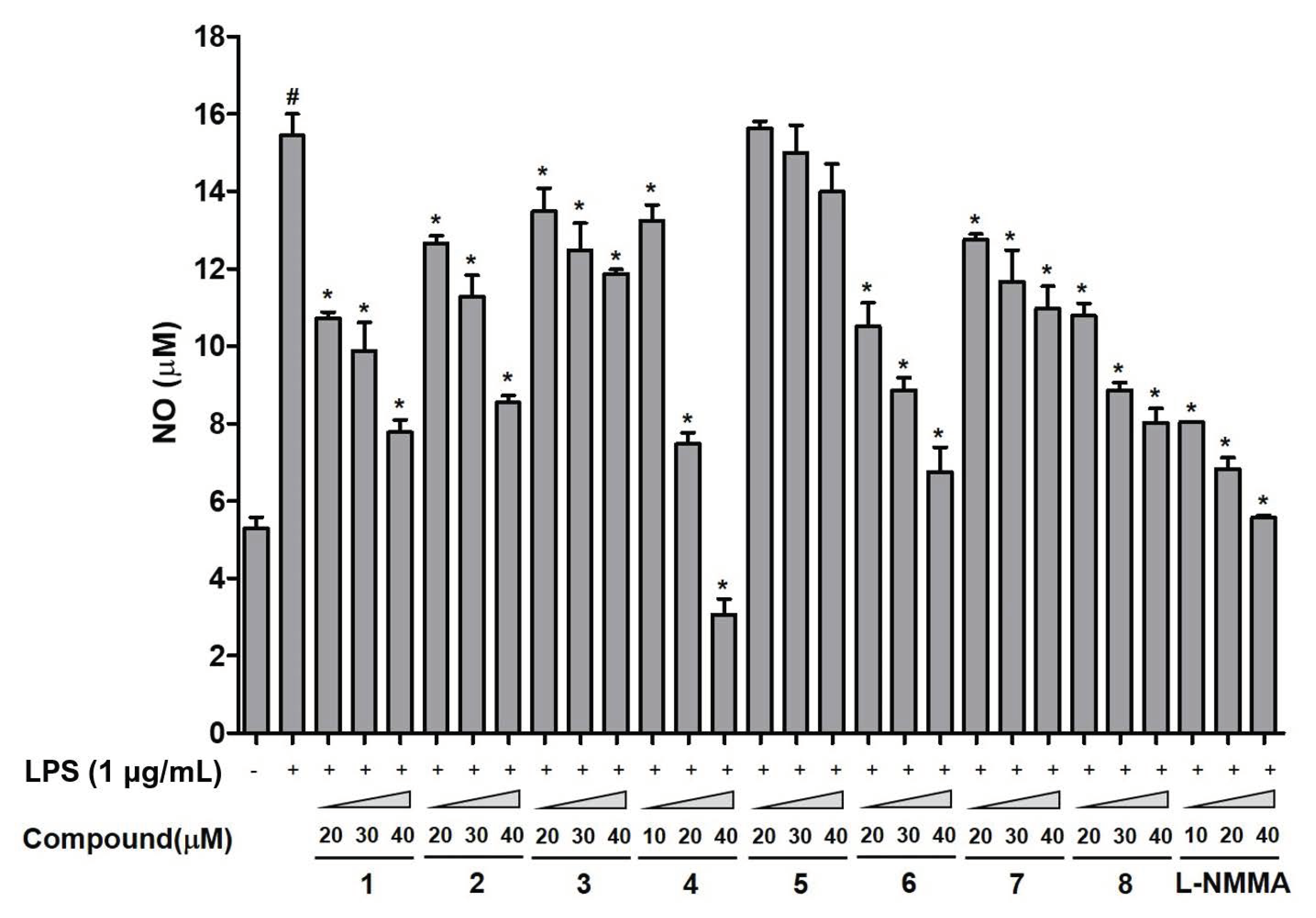
3.3. Evaluation of the Biological Effects of the Isolates from the Hulls of T. aestivum
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemicals and antioxidant activity of milled fractions of different wheat varieties. J. Agric. Food Chem. 2005, 53, 2297–2306. [Google Scholar] [CrossRef]
- Liu, R.H. Whole grain phytochemicals and health. J. Cereal Sci. 2007, 46, 207–219. [Google Scholar] [CrossRef]
- Hong, M.J.; Kim, D.Y.; Nam, B.M.; Ahn, J.-W.; Kwon, S.-J.; Seo, Y.W.; Kim, J.-B. Characterization of novel mutants of hexaploid wheat (Triticum aestivum L.) with various depths of purple grain color and antioxidant capacity. J. Sci. Food Agric. 2019, 99, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Sang, S. Phytochemicals in whole grain wheat and their health-promoting effects. Mol. Nutr. Food Res. 2017, 61, 1600852. [Google Scholar] [CrossRef] [PubMed]
- Deroover, L.; Tie, Y.; Verspreet, J.; Courtin, C.M.; Verbeke, K. Modifying wheat bran to improve its health benefits. Crit. Rev. Food Sci. Nutr. 2020, 60, 1104–1122. [Google Scholar] [CrossRef]
- Budhwar, S.; Chakraborty, M.; Sethi, K.; Chatterjee, A. Antidiabetic properties of rice and wheat bran—A review. J. Food Biochem. 2020, 44, e13424. [Google Scholar] [CrossRef]
- Arun, K.B.; Dhanya, R.; Chandran, J.; Abraham, B.; Satyan, S.; Nisha, P. A comparative study to elucidate the biological activities of crude extracts from rice bran and wheat bran in cell line models. J. Food Sci. Technol. 2020, 57, 3221–3231. [Google Scholar] [CrossRef]
- Lee, J.Y.; Ahn, E.-K.; Park, J.-H.; Oh, J.S. Wheat bran extract regulates mast cell-mediated allergic responses in vitro and in vivo. Molecules 2020, 25, 3997. [Google Scholar] [CrossRef]
- Bledzki, A.K.; Mamun, A.A.; Bonnia, N.N.; Ahmad, S. Basic properties of grain by-products and their viability in polypropylene composites. Ind. Crops Prod. 2012, 37, 427–434. [Google Scholar] [CrossRef]
- Kowalska, I.; Pecio, L.; Ciesla, L.; Oleszek, W.; Stochmal, A. Isolation, chemical characterization, and free radical scavenging activity of phenolics from Triticum aestivum L. aerial parts. J. Agric. Food Chem. 2014, 62, 11200–11208. [Google Scholar] [CrossRef]
- Zhu, Y.; Soroka, D.; Sang, S. Oxyphytosterols as active ingredients in wheat bran suppress human colon cancer cell growth: Identification, chemical synthesis, and biological evaluation. J. Agric. Food Chem. 2015, 63, 2264–2276. [Google Scholar] [CrossRef]
- Moheb, A.; Grondin, M.; Ibrahim, R.K.; Roy, R.; Sarhan, F. Winter wheat hull (husk) is a valuable source for tricin, a potential selective cytotoxic agent. Food Chem. 2013, 138, 931–937. [Google Scholar] [CrossRef]
- Hong, S.S.; Oh, J.S. Inhibitors of antigen-induced degranulation of RBL-2H3 cells isolated from wheat bran. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 69–74. [Google Scholar] [CrossRef]
- Luyen, B.T.T.; Thao, N.P.; Tai, B.H.; Lim, J.Y.; Ki, H.H.; Kim, D.K.; Lee, Y.M.; Kim, Y.H. Chemical constituents of Triticum aestivum and their effects on adipogenic differentiation of 3T3-L1 preadipocytes. Arch. Pharm. Res. 2015, 38, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Poudel, B.; Nepali, S.; Xin, M.; Ki, H.-H.; Kim, Y.-H.; Kim, D.-K.; Lee, Y.-M. Flavonoids from Triticum aestivum inhibit adipogenesis in 3T3-L1 cells by upregulating the insig pathway. Mol. Med. Rep. 2015, 12, 3139–3145. [Google Scholar] [CrossRef] [PubMed]
- Moncada, S.; Higgs, E.A. Molecular mechanisms and therapeutic strategies related to nitric oxide. FASEB J. 1995, 9, 1319–1330. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, H.; Bartsch, H. Chronic infections and inflammatory processes as cancer risk factors: Possible role of nitric oxide in carcinogenesis. Mutat. Res. 1994, 305, 253–264. [Google Scholar] [CrossRef]
- Krönche, K.D.; Fensel, K.; Kolb-Bachofen, V. Inducible nitric oxide synthase in human diseases. Clin. Exp. Immunol. 1998, 113, 147–156. [Google Scholar] [CrossRef]
- Nathan, C.; Xie, Q.W. Regulation of biosynthesis of nitric oxide. J. Biol. Chem. 1994, 269, 13725–13728. [Google Scholar]
- Hobbs, A.J.; Higgs, A.; Moncada, S. Inhibition of nitric oxide synthase as a potential therapeutic target. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 191–220. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef] [PubMed]
- Coleman, J.W. Nitric oxide in immunity and inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef]
- Zamora, R.; Vodovotz, Y.; Billiar, T.R. Inducible nitric oxide synthase and inflammatory diseases. Mol. Med. 2000, 6, 347–373. [Google Scholar] [CrossRef] [PubMed]
- Bai, N.; He, K.; Roller, M.; Lai, C.-S.; Bai, L.; Pan, M.-H. Flavonolignans and other constituents from Lepidium meyenii with activities in anti-inflammation and human cancer cell lines. J. Agric. Food Chem. 2015, 63, 2458–2463. [Google Scholar] [CrossRef]
- Nakajima, Y.; Yun, Y.S.; Kunugi, A. Six new flavonolignans from Sasa veitchii (Carr.) Rehder. Tetrahedron 2003, 59, 8011–8015. [Google Scholar] [CrossRef]
- Jeong, Y.H.; Lee, J.; Kwon, Y.J.; Seo, E.K. Reverse-phase HPLC method for identification of diastereomeric constituents from Sasa Boreal. Yakhak Hoeji 2006, 50, 21–25. [Google Scholar]
- Lee, S.-S.; Baek, Y.-S.; Eun, C.-S.; Yu, M.-H.; Baek, N.-I.; Chung, D.-K.; Bang, M.-H.; Yang, S.-A. Tricin derivatives as anti-inflammatory and anti-allergic constituents from the aerial part of Zizania latifolia. Biosci. Biotechnol. Biochem. 2015, 79, 700–706. [Google Scholar] [CrossRef]
- Liu, Q.; Wu, C.; Peng, A.; Gao, K.; Che, J.; Li, Y.; Fu, H. Flavonolignans from Elymus natans L. and phytotoxic activities. J. Agric. Food Chem. 2017, 65, 1320–1327. [Google Scholar] [CrossRef]
- Bouaziz, M.; Veitch, N.C.; Grayer, R.J.; Simmonds, M.S.J.; Damak, M. Flavonolignans from Hyparrhenia hirta. Phytochemistry 2002, 60, 515–520. [Google Scholar] [CrossRef]
- Chang, C.-L.; Wang, G.-J.; Zhang, L.-J.; Tsai, W.-J.; Chen, R.-Y.; Wu, Y.-C.; Kuo, Y.-H. Cardiovascular protective flavonolignans and flavonoids from Calamus quiquesetinervius. Phytochemistry 2010, 71, 271–279. [Google Scholar] [CrossRef]
- Wenzig, E.; Kunert, O.; Ferreira, D.; Schmid, M.; Schühly, W.; Bauer, R.; Hiermann, A. Flavonolignans from Avena sativa. J. Nat. Prod. 2005, 68, 289–292. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Sato, H.; Nakanishi, K. Configurational and conformational studies of sugars by the aromatic chirality method. J. Chem. Soc. Chem. Commun. 1970, 1691–1693. [Google Scholar] [CrossRef]
- Li, M.; Pu, Y.; Yoo, C.G.; Ragauskas, A.J. The occurrence of tricin and its derivatives in plants. Green Chem. 2016, 18, 1439. [Google Scholar] [CrossRef]
- Jeong, R.-H.; Lee, D.-Y.; Cho, J.-G.; Lee, S.-M.; Kang, H.-C.; Seo, W.-D.; Kang, H.-W.; Kim, J.-Y.; Baek, N.-I. A new flavonolignan from the aerial parts of Oryza sativa L. inhibits nitric oxide production in RAW 264.7 macrophage cells. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 865–870. [Google Scholar] [CrossRef]
- Mohanlal, S.; Maney, S.K.; Santhoshkumar, T.R.; Jayalekshmy, A. Tricin 4’-O-(erythro-β-guaiacylglyceryl) ether and tricin 4’-O-(threo-β-guaiacylglyceryl) ether isolated from Njavara (Oryza sativa L. var. Njavara), induce apoptosis in multiple tumor cells by mitochondrial pathway. J. Nat. Med. 2013, 67, 528–533. [Google Scholar] [CrossRef]
- Mohanlal, S.; Parvathy, R.; Shalini, V.; Helen, A.; Jayalekshmy, A. Isolation, characterization and quantification of tricin and flavonolignans in the medicinal rice Njavara (Oryza sativa L.), as compared to staple varieties. Plant. Foods Hum. Nutr. 2011, 66, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Chen, W.; Wang, W.; Zhang, H.; Liu, X.; Luo, J. Comprehensive profiling and natural variation of flavonoids in rice. J. Integr. Plant Biol. 2014, 56, 876–886. [Google Scholar] [CrossRef]
- Jung, Y.-J.; Park, J.-H.; Cho, J.-G.; Seo, K.-H.; Lee, D.-S.; Kim, Y.-C.; Kang, H.-C.; Song, M.-C.; Baek, N.-I. Lignan and flavonoids from the stems of Zea mays and their anti-inflammatory and neuroprotective activities. Arch. Pharm. Res. 2015, 38, 178–185. [Google Scholar] [CrossRef]
- Huang, H.; Liu, Y.; Meng, Q.; Wei, S.; Cui, H.; Zhang, C. Flavonolignans and other phenolic compounds from Sorghum halepense (L.) Pers. Biochem. Syst. Ecol. 2010, 38, 656–658. [Google Scholar] [CrossRef]
- Colombo, R.; Yariwake, J.H.; Queiroz, E.F.; Ndjoko, K.; Hostettmann, K. On-line identification of sugarcane (Saccharum officinarum L.) methoxyflavones by liquid chromatography–UV detection using post-column derivatization and liquid chromatography–mass spectrometry. J. Chromatogr. A 2005, 1082, 51–59. [Google Scholar] [CrossRef]
- Colombo, R.; Yariwake, J.H.; Queiroz, E.F.; Ndjoko, K.; Hostettmann, K. On-line identification of further flavone C- and O-glycosides from sugarcane (Saccharum officinarum L., Gramineae) by HPLC-UV-MS. Phytochem. Anal. 2006, 17, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Bottcher, A.; Cesarino, I.; dos Santos, A.B.; Vicentini, R.; Mayer, J.L.S.; Vanholme, R.; Morreel, K.; Goeminne, G.; Moura, J.C.M.S.; Nobile, P.M. Lignification in sugarcane: Biochemical characterization, gene discovery, and expression analysis in two genotypes contrasting for lignin content. Plant Physiol. 2013, 163, 1539–1557. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.-C.; Lu, Z.-X.; Xu, H.-X.; Zheng, X.-S.; Yang, Y.-J. Chemical constituents from the aerial parts of Vetiveria zizanioides, Chem. Nat. Compd. 2012, 48, 128–129. [Google Scholar] [CrossRef]
- Chang, C.-L.; Zhang, L.-J.; Chen, R.-Y.; Wu, C.-C.; Huang, H.-C.; Roy, M.C.; Huang, J.-P.; Wu, Y.-C.; Kuo, Y.-H. Quiquelignan A–H, eight new lignoids from the rattan palm Calamus quiquesetinervius and their antiradical, anti-inflammatory and antiplatelet aggregation activities. Bioorga. Med. Chem. 2010, 18, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Qiu, D.; Guo, J.; Yu, H.; Yan, J.; Yang, S.; Li, X.; Zhang, Y.; Sun, J.; Cong, J.; He, S.; et al. Antioxidant phenolic compounds isolated from wild Pyrus ussuriensis Maxim. fruit peels and leaves. Food Chem. 2018, 15, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Shao, Y.; Wang, Q.; Mei, L.; Tao, Y. Purification of flavonolignan diastereoisomers from Arenaria kansuensis by two-dimensional liquid chromatography combined with solid-phase extraction. J. Chromatogr. Sci. 2019, 57, 418–425. [Google Scholar] [CrossRef]
- Choi, J.; Yoon, K.D.; Kim, J. Chemical constituents from Taraxacum officinale and their α-glucosidase inhibitory activities. Bioorg. Med. Chem. Lett. 2018, 28, 476–481. [Google Scholar] [CrossRef]
- Kang, B.-M.; An, B.-K.; Jung, W.-S.; Jung, H.-K.; Cho, J.-H.; Cho, H.-W.; Jang, S.J.; Yun, Y.B.; Kuk, Y.I. Anti-inflammatory effect of tricin isolated from Alopecurus aequalis Sobol on the LPS-induced inflammatory response in RAW 264.7 cells. Int. J. Mol. Med. 2016, 38, 1614–1620. [Google Scholar] [CrossRef]
- Gautam, R.; Jachak, S.M. Recent developments in anti-inflammatory natural products. Med. Res. Rev. 2009, 29, 767–820. [Google Scholar] [CrossRef]
- Azab, A.; Nassar, A.; Azab, A.N. Anti-inflammatory activity of natural products. Molecules 2016, 21, 1321. [Google Scholar] [CrossRef]
- Lau, T.A.; Bray, W.M.; Lokey, R.S. Macrophage cytological profiling and anti-Inflammatory drug discovery. Assay Drug Dev. Technol. 2019, 17, 14–16. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Compounds | IC50 (μM) | Cell Viability (%) 1 |
|---|---|---|
| 1 | 46.41 | >100 |
| 2 | 58.95 | 99.7 ± 2.1 |
| 3 | >100 | 99.3 ± 2.3 |
| 4 | 24.14 | 85.7 ± 4.0 |
| 5 | >100 | 98.8 ± 3.2 |
| 6 | 37.52 | >100 |
| 7 | >100 | 99.8 ± 10.8 |
| 8 | 45.67 | >100 |
| L-NMMA 2 | 51.01 | >100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, A.-R.; Kil, Y.-S.; Hong, M.J.; Park, J.; Park, H.H.; Jin, C.H.; Nam, J.-W.; Kim, J.-B. Anti-Inflammatory Flavonolignans from Triticum aestivum Linn. Hull. Appl. Sci. 2020, 10, 8656. https://doi.org/10.3390/app10238656
Han A-R, Kil Y-S, Hong MJ, Park J, Park HH, Jin CH, Nam J-W, Kim J-B. Anti-Inflammatory Flavonolignans from Triticum aestivum Linn. Hull. Applied Sciences. 2020; 10(23):8656. https://doi.org/10.3390/app10238656
Chicago/Turabian StyleHan, Ah-Reum, Yun-Seo Kil, Min Jeong Hong, Jisu Park, Hyeon Hwa Park, Chang Hyun Jin, Joo-Won Nam, and Jin-Baek Kim. 2020. "Anti-Inflammatory Flavonolignans from Triticum aestivum Linn. Hull" Applied Sciences 10, no. 23: 8656. https://doi.org/10.3390/app10238656
APA StyleHan, A.-R., Kil, Y.-S., Hong, M. J., Park, J., Park, H. H., Jin, C. H., Nam, J.-W., & Kim, J.-B. (2020). Anti-Inflammatory Flavonolignans from Triticum aestivum Linn. Hull. Applied Sciences, 10(23), 8656. https://doi.org/10.3390/app10238656