Abstract
Sialic acids in the structural matrix of biofilms developing in engineered water systems constitute a potential target in the battle against biofouling. This report focuses specifically on the presence of sialic acids as part of the extracellular polymeric substances (EPS) of biofilms forming in cooling towers and the potential effect of nutrient starvation on sialic acid presence and abundance. Two cooling water compositions were compared in parallel pilot-scale cooling towers, one poor in nutrients and one enriched in nutrients. Fresh deposits from the two cooling towers were collected after a five-week operation period. EPS extractions and analyses by Fourier transform infrared spectroscopy (FTIR) and high-resolution mass spectrometry (MS), along with 16S rRNA gene amplicon sequencing were performed. The results of MS analyses showed the presence of pseudaminic/legionaminic acids (Pse/Leg) and 2-keto-3-deoxy-d-glycero-d-galacto-nononic acid (KDN) in both biofilm EPS samples. FTIR measurements showed the characteristic vibration of sialic acid-like compounds ν(C=O)OH in the nutrient poor sample exclusively. Our findings, combined with other recent studies, suggest that bacterial sialic acids are common compounds in environmental biofilms. Additionally, the conservation of sialic acid production pathways under nutrient starvation highlights their importance as constituents of the EPS. Further in-depth studies are necessary to understand the role of sialic acids in the structural cohesion and protection of environmental biofilm layer.
Keywords:
biofouling; industry; EPS; sialic acids; mass spectrometry; environmental biofilm; cleaning 1. Introduction
Biofilm formation is highly detrimental to the daily operation and lifetime of engineered water processes, such as heat exchangers and membrane filtration systems. Commonly applied chemical cleanings of fouled units do not manage to recover the initial efficiency of the systems. This is often due to a lack of understanding of the extracellular polymeric substances (EPS) formed by the microorganisms [1], eventually resulting in the use of non-adapted chemicals (e.g., surfactants or acid and base cleaning products).
Biofilm formation is facilitated by the production of EPS, consisting of carbohydrates, proteins, nucleic acids, and other biopolymers [1]. Carbohydrates produced by the microorganisms exhibit great diversity. Within those carbohydrates, sialic acids, a subset of the nonulosonic acid family comprising more than 50 structurally distinct acidic carbohydrates, have been proposed to play an important role in biofilm formation related to human bacterial infections [2]. For example, these nine-carbon acidic sugars can act as growth and adhesion factors for some periodontal organisms, both for colonization of host surfaces and interactions with other oral dwelling bacteria within a biofilm [3]. Since it was widely believed that the ability of bacteria to biosynthesize sialic acids was predominantly found in pathogenic and commensal species [4], little study has been performed on environmental biofilms.
Sialic acids have very recently been reported in water treatment systems as part of the EPS of seawater-adapted granular sludge [5]. In addition, a newly established mass spectrometric approach by Kleikamp et al. [6] revealed their wide-spread occurrence in non-pathogenic species. These recent discoveries raise interesting questions regarding the occurrence of sialic acids in the biofouling layer of engineered water systems. Investigation of sialic acids in samples from water processes, such as cooling towers, would contribute to a better understanding of the biofilm composition and encourage further in-depth studies on their functions as a component of the extracellular matrix.
Furthermore, sialic acids are present on the surface of all vertebrate cells where they terminate glycan chains and are exposed to many interactions with the surrounding environment [7]. In granular activated sludge, they were found both on bacterial surface-related structures and in the EPS of granular activated sludge, i.e., they are located within the space between the cells and their environment [5]. In this respect, it is reasonable to assume that changes in the surrounding environment may also have an impact on sialic acid production. Exploring the potential impact of environmental stresses (e.g., nutrient starvation) on their diversity and abundance in environmental biofilms would provide significant insights on their protective role and relevance in the matrix.
For this study, biofilm samples were collected at the end of a 5-week experiment from pilot-scale cooling tower systems operated in parallel. The full experiment has been previously described by Pinel et al. [8], investigating the efficiency of phosphate limitation as a biofouling control method for cooling towers. In the present report, sialic acid identification and relative abundance were assessed from the biofilms obtained from two cooling towers supplied with seawater reverse osmosis permeate—the first without addition of nutrients (nutrient poor), primarily limited by phosphorus, and the second with addition of nutrients (nutrient enriched). The objectives are to (i) evaluate the presence of sialic acids in EPS of environmental biofilms and estimate the potential for sialic acid production by the biofilm bacterial communities, (ii) investigate the impact of nutrient poor condition on the presence and abundance of sialic acids, and (iii) explore the implications of the findings related to engineered water processes prone to biofouling.
2. Materials and Methods
2.1. Pilot Set-Up
The cooling tower pilot facility is located at the King Abdullah University of Science and Technology (KAUST) in Saudi Arabia. The process and operating parameters have been previously described [8,9]. Two parallel cooling towers were considered in this study. Seawater reverse osmosis (SWRO) permeate produced by the nearby desalination plant [10] was used as feed water during the 5-week experiment. Cycles of concentration were maintained between 5 and 7, with cooling water conductivities varying between 6 and 8 mS/cm, and pH between 8.0 and 8.3. The temperature of the water in the cooling tower basins varied between 27 and 30 °C, depending on external climate factors.
Biofilms were developed under the following conditions: (i) poor in nutrients, i.e., SWRO permeate without additional nutrients as feed water, and (ii) enriched in nutrients, i.e., SWRO permeate with addition of 0.5:0.1:0.05 (C:N:P) mg/L and trace metals as feed water. Concentrations were chosen as described in Pinel et al. [8]. The C, N, and P sources used in the enriched nutrient condition were as follows: sodium acetate trihydrate (CH3COONa·3H2O), sodium nitrate (NaNO3), and monosodium phosphate monohydrate (NaH2PO4·H2O), all purchased from Sigma Aldrich.
2.2. Samples Collection
The collected deposits were the same as described in Pinel et al. [8]. Deposits were sampled from the basin surface of the cooling towers after drainage of the two basins at the end of 5 consecutive weeks of stable operation. Pieces measuring 4 cm × 4 cm were collected in 50 mL sterile Greiner tubes. Locations of the samples were arbitrary selected but repeated identically in the cooling towers. Two of the pieces from each investigated cooling tower—nutrient enriched or nutrient poor—were used for the analyses described below.
2.3. Microscopic Pictures
Deposit samples from the cooling tower basins were frozen at −80 °C upon collection and lyophilized at −50 °C and 0.05 mbar (Alpha 1–4 LDplus, Martin Christ, Osterode am Harz, Germany). Microscopic pictures of the freeze-dried raw deposits were taken with a Zeiss AxioPlan 2 imaging microscope (Carl Zeiss, Oberkochen, Germany). Samples were observed with normal light and all images were obtained with a 400× total magnification.
2.4. EPS Extraction
Extraction of EPS was performed on the freeze-dried raw deposits. EPS was extracted at 80 °C in alkaline conditions, following a method previously described [8], and lyophilized. The freeze-dried EPS samples were kept in a dry environment before further analyses.
2.5. Fourier Transform Infrared Spectroscopy (FTIR)
The Fourier transform infra-red (FTIR) spectra of the freeze-dried raw deposits and extracted EPS samples were performed on a FTIR Spectrophotometer (Perkin-Elmer, Shelton, CT, USA) at room temperature, with a wavenumber range from 500 cm−1 to 4000 cm−1. Resolution of 4 cm−1 and accumulation of 8 scans were applied on each sample.
2.6. High-Resolution Mass Spectrometry (MS)
Freeze-dried extracted EPS samples were analyzed for sialic acids using a recently established untargeted high-resolution mass spectrometric approach, described by Kleikamp et al. [6]. Sialic acid peaks from 2-keto-3-deoxy-d-glycero-d-galacto-nononic acid (KDN) and pseudaminic/legionaminic acids (Pse/Leg) identified by mass and characteristic fragment ions were integrated using Thermo Xcalibur Qualbrowser. Observed and theoretical masses, C-9 marker fragments and water loss peaks are provided as Supplementary Information (Table S1). Analyses were performed in duplicates. A statistical analysis was conducted on the sialic acid peak areas at 95% confidence level based on Z-scores. Variations in abundances with p-values higher than 0.05 were considered statistically insignificant.
2.7. DNA Extraction and 16S rRNA Gene Amplicon Sequencing
The raw deposit samples from the basin surface were frozen at −20 °C upon collection and until DNA extraction. The genomic DNA of 0.5 g of sample was extracted using the DNeasy Power Water kit (Qiagen, Germany) according to the manufacturer’s instructions. DNA concentration was quantified using a Qubit dsDNA high sensitivity (HS) or Qubit broad range (BR) assay kit (Life Technologies, Carlsbad CA, USA). The extracted DNA was stored at −80 °C until further analysis. The DNA extracts were sent to DNASense ApS (Aalborg, Denmark) for amplicon sequencing of the 16S rRNA gene targeting the variable region V4 (position 515–806). Samples were paired-end sequenced (2 × 300 bp) on an Illumina MiSeq instrument. The reads were trimmed for quality, clustered and OTU abundances were generated based on 97% similarities using USEARCH. Taxonomy was assigned using the Ribosomal Database Project (RDP) classifier [11] as implemented in the parallel_assign_taxonomy_rdp.py script in QIIME [12], with a confidence of 0.8 and the SILVA database. OTUs were generated based on a subsampling parameter of 10,000 sequences per sample. Extraction and analyses were performed in duplicates. Alpha and beta diversity analyses are available as Supplementary Information (Table S3 and Figure S1).
2.8. Basic Local Alignment Search Tool (BLAST)
The Basic Local Alignment Search Tool (BLAST) was accessed on the NCBI website. Reference protein sequences of CMP-pseudaminic acid (Pse), CMP-legionaminic acid (Leg), and CMP-2-keto-3-deoxy-d-glycero-d-galacto-nononic acid (KDN) synthetases [13,14] were blasted against the available NCBI databases sequences of the main bacterial groups, families or genera, present in the biofilms. The compared sequences were considered having a significant match when the E-value was inferior to 10−15.
3. Results and Discussion
3.1. Sialic Acids Are Present in Environmental Biofilms
Biofilms formed at the bottom of the cooling tower basins—under (i) nutrient poor and (ii) nutrient enriched conditions. Figure 1 shows pictures of the biofilms before collection (A and B) and microscopic images of the collected samples (C and D). Both biofilm samples contained a complex ecosystem within the matrix, including both algae and bacteria. Visual observations revealed a less extensive biofilm formed under nutrient poor conditions with lower algae growth than in the nutrient enriched biofilm.

Figure 1.
Biofilm deposit in the basins of the cooling tower fed with reverse osmosis (RO) permeate poor in nutrients (A) and the cooling tower fed with RO permeate enriched in nutrients (B) and associated microscopic pictures (C,D) respectively.
The 16S rRNA gene amplicon sequencing analyses revealed large fractions of Proteobacteria: 32 and 19%, Cyanobacteria: 25 and 57%, and Planctomycetes: 21 and 7% in the nutrient poor and nutrient enriched biofilms respectively. The relative abundances of the main families are shown in Figure 2. Microscillaceae, Leptolyngbyaceae and an undefined Nostocales family were the most represented groups in both samples.
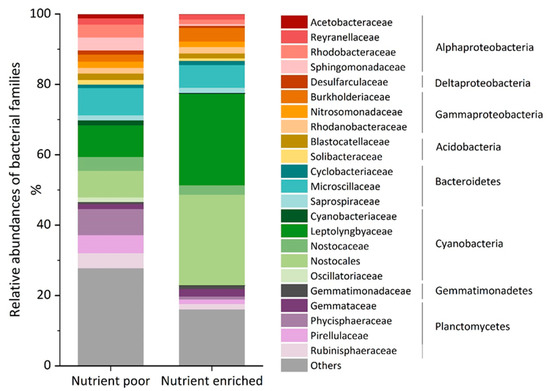
Figure 2.
Relative abundance of the main bacterial families in the biofilm samples grown under nutrient poor and nutrient enriched conditions with corresponding phyla and class of Proteobacteria. “Others” contains bacterial families of less than 1% abundance in both samples.
Due to the presence of a large fraction of inorganic particles (e.g., sand) accumulated in the biofilm, EPS was extracted to eliminate the inorganic background and most intracellular compounds. The presence of sialic acids and their relative abundance in the extracted biofilm EPS samples are displayed in Figure 3. Sialic acids with the compositions consistent with Pse/Leg, which are indistinguishable by mass spectrometry only, and KDN, or a related desamino form of a molecule consistent with the neuraminic acid composition, were present in both biofilm EPS samples. KDN was reported to be part of glycoconjugates, including glycolipids, glycoproteins, and capsular polysaccharides, at the terminate position of the glycan chain. Pse/Leg have been mainly found as constituents of heteropolysaccharides, as internal carbohydrate [15]. Their charged group allows both sialic acids to engage in cross-linking, potentially protecting the biofilm matrix.
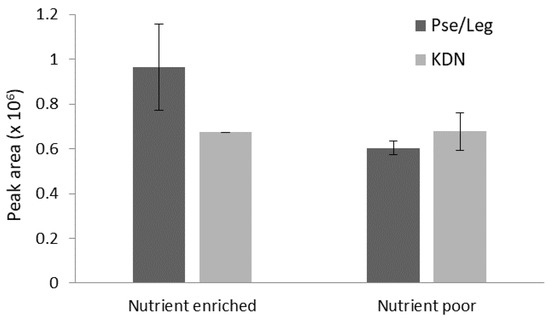
Figure 3.
Observed mass spectrometric peak areas of sialic acids in nutrient enriched and nutrient poor biofilm extracellular polymeric substances (EPS). The error bars represent the minimum and maximum values. Abundances of both sialic acids were not significantly different between the samples (p-values > 0.05).
Sialyltransferases are key enzymes in the biosynthesis of sialic acid-containing oligosaccharides and glycoconjugates [16]. They catalyze the transfer reaction of a sialic acid residue to the glycoconjugates. Presence of these compounds in the extracellular matrix indicates that some bacterial members of the biofilms are able to produce the transferases. Reference protein sequences of the transferases were compared to the most abundant bacterial groups in the biofilms in order to approximately evaluate the potential of sialic acid production by the bacterial community. Since Campylobacter jejuni has characterized pathways for biosynthesis of sialic acids (e.g., Leg and Pse), sialyltransferases in these pathways were used as reference protein sequences [13]. The reference sequence for KDN was obtained from Wang et al. [14]. A BLAST analysis was then performed to compare the reference sequences of the enzymes CMP-Pse, CMP-Leg, and CMP-KDN synthetases with the NCBI database sequences of the identified bacterial groups (Figure 2). Significant matches (E-value < 10−15) are shown in Table 1 for the main families and corresponding genera identified in the biofilm samples. A large part of the genera present in both biofilm samples includes species that possess the genes responsible for the sialic acid residue transfer to the glycoconjugates, suggesting that sialic acids can be produced by these microorganisms.

Table 1.
Results of The Basic Local Alignment Search Tool (BLAST) analysis comparing the enzymes CMP-pseudaminic acid (CMP-Pse), CMP-legionaminic acid (CMP-Leg), and CMP-2-keto-3-deoxy-d-glycero-d-galacto-nononic acid (CMP-KDN) synthetases to the available NCBI database sequences of the bacterial groups in the biofilms (identified by 16S rRNA gene amplicon sequencing). The main families (>1%) and identified genera in these families, all present in both biofilm samples, were blasted. Significant matches (E-value < 10−15) are indicated by a green background, no match or insignificant matches (E-value > 10−15) by a red background.
In short, the mass spectrometric measurements showed that diverse sialic acids were produced in the two environmental biofilms collected from the cooling towers. Results of the BLAST analysis, combined to observations from earlier studies, indicate that a wide range of microorganisms have the ability to produce these compounds. This confirms that sialic acid-containing glycoconjugates are more widely found in environmental biofilms than previously thought [6,7,16,17,18].
3.2. Nutrient Availability Did Not Significantly Influence the Production of Sialic Acids in the Environmental Biofilms
Due to the exposure of sialic acid molecules to the surrounding environment, both as terminal position of the glycan chain on vertebrate cell surfaces [7] and as constituent of the EPS of granular sludge [5], it was assumed that changes in the direct environment would impact sialic acid production. The influence of nutrient availability—nutrient poor and nutrient enriched conditions in this study—was, however, not reflected by the MS results of the biofilm EPS extracts. Two different types of sialic acids, Pse/Leg and KDN (methylated), were detected in both the nutrient poor and nutrient enriched biofilm samples (Figure 3). Their abundances were not significantly different between the samples (p-values > 0.05 based on Z-scores). The fact that sialic acid production is maintained under nutrient starvation suggests that these carbohydrates do not only contribute to biofilm formation but might also have value under stress conditions to maintain protection and structure of the biofilm. Firstly, the acidic functional group and the ability of forming ionic bond with multivalent cations, such as Ca2+ [19], suggest the involvement of the molecules in complex network and contribution to the structural integrity of the overall biofilm. Secondly, sialic acids are not only produced and used by microorganisms to colonize host surfaces, but are also produced and released in the EPS to protect the biofilm and microbiome from environmental stresses [5,17,20,21]. A recent study on granular sludge has shown that sialic acids, due to their terminal position and large size with nine carbons, were able to protect the granules from enzymatic degradation [5]. Other systems have shown similar behavior, such as the CMP-sialic acid transporter which is upregulated in diatoms under silica-limited conditions. This transporter transfers CMP-sialic acid from the cytosol into Golgi vesicles where glycosyltransferases are activated and produce sialylated glycolipids that can act as defensive molecules. Therefore, this upregulation indicates that diatom cells are coping with starvation by accumulating defensive molecules [20]. Another example are cancer cells, which react to nutrient-deprived conditions by using sialic acids to maintain cell surface glycosylation, through complementary mechanisms [21].
The FTIR analysis shows a sharp peak around 1738 cm−1 and a shoulder peak at 1730 cm−1 in the nutrient poor EPS spectrum, which are absent from the nutrient enriched EPS spectrum (Figure 4). The shoulder peak at 1730 cm−1 has been previously associated to the alpha-keto aldonic acid structure in sialic acids [5]. The presence of this peak in the FTIR analysis of the nutrient poor EPS might indicate additional production of other ulosonic acid variants (e.g., 7 or 8 carbon sugars) at larger quantities potentially induced by nutrient starvation, or increased exposure of the observed nine-carbon sugars. Correlation between FTIR and MS analyses needs to be further investigated.
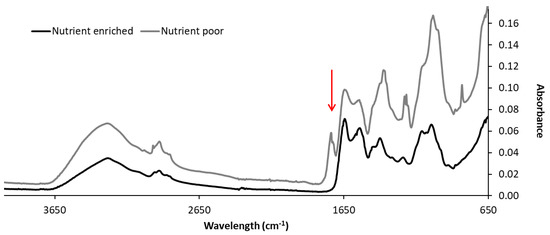
Figure 4.
FTIR of the extracted EPS of the biofilms. The arrow shows the peak at 1730 cm−2 associated with the ν(C=O)OH of sialic acids.
To summarize, changes in nutrient availability did not affect the types and abundances of sialic acids produced by the bacterial community. The conservation of energetically costly production pathways of sialic acids under nutrient starvation highlights their importance as a constituent of EPS and raises questions on their function in environmental biofilms.
3.3. Relevance to Industrial Processes and Future Considerations
Removal of biofilms in industrial processes and from medical devices, specially grown over a long period of time, is very challenging with conventional chemical cleaning methods (e.g., surfactant and chlorine dosages or acid and base cleanings). Thanks to increasing awareness of the role of EPS on biofilm integrity and efforts in understanding its composition, alternative biofilm removal methods are being developed [22,23,24]. The use of enzymes able to cleave the sugar chains and proteins of the extracellular matrix have shown promising results in the medical field for solubilizing the biofilm and preventing the spread of infections [24,25]. However, enzymatic treatments face limitations when used on environmental biofilms due to a lack of substrate specific enzymes and heterogeneous mixture of extracellular molecules [26].
A study on the efficiency of acylase I and proteinase K on removal of biofilms from reverse osmosis membranes revealed that the enzymes were ineffective on some fractions of the EPS and should be used with additional EPS degrading agents to improve cleaning [27]. Their analyses showed that the treatments removed humic-like substances but did not affect protein-like molecules. This is likely caused by the presence of functional groups protecting the macromolecules from degradation. In this context, sialic acids might be protecting the matrix from such enzymatic agents, as it is suggested in recent research [5,17] and supported by our preliminary results. Cleavage of the molecule would weaken the matrix and provide exposure of the glycoproteins to enzymatic hydrolysis. Additionally, the combination with emerging physical biofilm disruptive strategies, such as magnetic fields or ultrasounds [28,29], could significantly improve the efficiency of the cleaning. The partial breakups and detachments of the biofilm would eventually allow a better penetration of biofilm degrading agents.
Hence, the potential combination of sialidases with established enzymatic agents, and biofilm disrupting technologies, needs to be further investigated as sialic acids might constitute a key compound in biofilm stability.
Supplementary Materials
The following are available online at https://www.mdpi.com/2076-3417/10/21/7694/s1, Figure S1: Venn diagram of the nutrient poor and nutrient enriched cooling tower biofilm based on the 16S rRNA gene amplicon sequencing data. 64% of the OTUs are shared between the two samples. 97% of the sequences are included in the shared OTUs; Table S1: Observed and theoretical masses (averaged from duplicate analyses) for Pse/Leg and KDN, including their characteristic C-9 marker fragments and water-loss peak masses; Table S2. Processing data of the raw sequences of the 16S rRNA gene amplicon sequencing indicating the raw reads, reads after low quality read removal (trimmed), after merging of forward and reverse reads (merged) and reads mapped to OTUs (mapped), Table S3: Alpha diversity analyses on the 16S rRNA gene amplicon sequencing data.
Author Contributions
Conceptualization, Y.L., M.C.M.v.L. and I.S.M.P.; Methodology, I.S.M.P. and Y.L.; Software, H.B.C.K., M.P. and I.S.M.P.; Validation, I.S.M.P. and H.B.C.K.; Formal Analysis, I.S.M.P. and H.B.C.K.; Investigation, I.S.M.P. and H.B.C.K.; Writing—Original Draft Preparation, I.S.M.P. and Y.L.; Writing—Review & Editing, M.P., M.C.M.v.L. and J.S.V.; Visualization, I.S.M.P.; Supervision, Y.L. and M.C.M.v.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Acknowledgments
The authors would like to thank Evides Industriewater B.V. for their financial support. Thank you also to Lan Hee Kim and Nadia Farhat for their contributions with the process operation and sample collections.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Seviour, T.; Derlon, N.; Dueholm, M.S.; Flemming, H.-C.; Girbal-Neuhauser, E.; Horn, H.; Kjelleberg, S.; van Loosdrecht, M.C.M.; Lotti, T.; Malpei, M.F.; et al. Extracellular polymeric substances of biofilms: Suffering from an identity crisis. Water Res. 2019, 151, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Trappetti, C.; Kadioglu, A.; Carter, M.; Hayre, J.; Iannelli, F.; Pozzi, G.; Andrew, P.W.; Oggioni, M.R. Sialic acid: A preventable signal for pneumococcal biofilm formation, colonization, and invasion of the host. J. Infect. Dis. 2009, 199, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Stafford, G.; Roy, S.; Honma, K.; Sharma, A. Sialic acid, periodontal pathogens and tannerella forsythia: Stick around and enjoy the feast! Mol. Oral Microbiol. 2012, 27, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Angata, T.; Varki, A. Chemical diversity in the sialic acids and related alpha-keto acids: An evolutionary perspective. Chem. Rev. 2002, 102, 439–469. [Google Scholar] [CrossRef]
- De Graaff, D.R.; Felz, S.; Neu, T.R.; Pronk, M.; van Loosdrecht, M.C.M.; Lin, Y. Sialic acids in the extracellular polymeric substances of seawater-adapted aerobic granular sludge. Water Res. 2019, 155, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Kleikamp, H.B.C.; Lin, Y.M.; McMillan, D.G.G.; Geelhoed, J.S.; Naus-Wiezer, S.N.H.; van Baarlen, P.; Saha, C.; Louwen, R.; Sorokin, D.Y.; van Loosdrecht, M.C.M.; et al. Tackling the chemical diversity of microbial nonulosonic acids—A universal large-scale survey approach. Chem. Sci. 2020, 11, 3074–3080. [Google Scholar] [CrossRef]
- Lewis, A.L.; Desa, N.; Hansen, E.E.; Knirel, Y.A.; Gordon, J.I.; Gagneux, P.; Nizet, V.; Varki, A. Innovations in host and microbial sialic acid biosynthesis revealed by phylogenomic prediction of nonulosonic acid structure. Proc. Natl. Acad. Sci. USA 2009, 106, 13552. [Google Scholar] [CrossRef] [PubMed]
- Pinel, I.S.M.; Kim, L.H.; Proença Borges, V.R.; Farhat, N.M.; Witkamp, G.J.; van Loosdrecht, M.C.M.; Vrouwenvelder, J.S. Effect of phosphate availability on biofilm formation in cooling towers. Biofouling 2020, 1–16. [Google Scholar]
- Al-Bloushi, M.; Saththasivam, J.; Jeong, S.; Amy, G.L.; Leiknes, T. Effect of organic on chemical oxidation for biofouling control in pilot-scale seawater cooling towers. J. Water Process Eng. 2017, 20, 1–7. [Google Scholar] [CrossRef]
- Belila, A.; El-Chakhtoura, J.; Otaibi, N.; Muyzer, G.; Gonzalez-Gil, G.; Saikaly, P.E.; van Loosdrecht, M.C.M.; Vrouwenvelder, J.S. Bacterial community structure and variation in a full-scale seawater desalination plant for drinking water production. Water Res. 2016, 94, 62–72. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbial. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [PubMed]
- Schoenhofen, I.C.; Vinogradov, E.; Whitfield, D.M.; Brisson, J.-R.; Logan, S.M. The cmp-legionaminic acid pathway in campylobacter: Biosynthesis involving novel gdp-linked precursors. Glycobiology 2009, 19, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Lu, Z.; Allen, K.N.; Mariano, P.S.; Dunaway-Mariano, D. Human symbiont bacteroides thetaiotaomicron synthesizes 2-keto-3-deoxy-d-glycero-d- galacto-nononic acid (kdn). Chem. Biol. 2008, 15, 893–897. [Google Scholar] [CrossRef][Green Version]
- Knirel, Y.A.; Shashkov, A.S.; Tsvetkov, Y.E.; Jansson, P.E.; Zãhringer, U. 5,7-diamino-3,5,7,9-tetradeoxynon-2-ulosonic acids in bacterial glycopolymers: Chemistry and biochemistry. ADV Carbohyd. Chem. Biochem. 2003, 58, 371–417. [Google Scholar]
- Harduin-Lepers, A.; Mollicone, R.; Delannoy, P.; Oriol, R. The animal sialyltransferases and sialyltransferase-related genes: A phylogenetic approach. Glycobiology 2005, 15, 805–817. [Google Scholar] [CrossRef] [PubMed]
- Boleij, M.; Kleikamp, H.; Pabst, M.; Neu, T.R.; van Loosdrecht, M.C.M.; Lin, Y. Decorating the anammox house: Sialic acids and sulfated glycosaminoglycans in the extracellular polymeric substances of anammox granular sludge. Environ. Sci. Technol. 2020, 54, 5218–5226. [Google Scholar] [CrossRef]
- Wagstaff, B.A.; Rejzek, M.; Field, R.A. Identification of a kdn biosynthesis pathway in the haptophyte prymnesium parvum suggests widespread sialic acid biosynthesis among microalgae. J. Biol. Chem. 2018, 293, 16277–16290. [Google Scholar] [CrossRef]
- Ganss, B.; Hoffmann, W. Calcium binding to sialic acids and its effect on the conformation of ependymins. J. Biochem. 1993, 217, 275–280. [Google Scholar] [CrossRef]
- Di Dato, V.; Di Costanzo, F.; Barbarinaldi, R.; Perna, A.; Ianora, A.; Romano, G. Unveiling the presence of biosynthetic pathways for bioactive compounds in the thalassiosira rotula transcriptome. Sci. Rep. 2019, 9, 9893. [Google Scholar] [CrossRef]
- Badr, H.A.; AlSadek, D.M.; Mathew, M.P.; Li, C.-Z.; Djansugurova, L.B.; Yarema, K.J.; Ahmed, H. Nutrient-deprived cancer cells preferentially use sialic acid to maintain cell surface glycosylation. Biomaterials 2015, 70, 23–36. [Google Scholar] [CrossRef] [PubMed]
- Kaur, A.; Rishi, V.; Soni, S.K.; Rishi, P. A novel multi-enzyme preparation produced from aspergillus niger using biodegradable waste: A possible option to combat heterogeneous biofilms. AMB Express 2020, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Jia, R.; Li, Y.; Gu, T. Advances in the treatment of problematic industrial biofilms. World J. Microb. Biotechnol. 2017, 33, 97. [Google Scholar] [CrossRef] [PubMed]
- Stiefel, P.; Mauerhofer, S.; Schneider, J.; Maniura-Weber, K.; Rosenberg, U.; Ren, Q. Enzymes enhance biofilm removal efficiency of cleaners. Antimicrob. Agents Chem. 2016, 60, 3647–3652. [Google Scholar] [CrossRef]
- Saggu, S.K.; Jha, G.; Mishra, P.C. Enzymatic degradation of biofilm by metalloprotease from microbacterium sp. SKS10. Front. Bioeng. Biotechnol. 2019, 7, 192. [Google Scholar] [CrossRef]
- Nahar, S.; Mizan, M.F.R.; Ha, A.J.-W.; Ha, S.-D. Advances and future prospects of enzyme-based biofilm prevention approaches in the food industry. Compr. Rev. Food. Sci. Food Saf. 2018, 17, 1484–1502. [Google Scholar] [CrossRef]
- Kim, L.H.; Kim, S.-J.; Kim, C.-M.; Shin, M.S.; Kook, S.; Kim, I.S. Effects of enzymatic treatment on the reduction of extracellular polymeric substances (eps) from biofouled membranes. Desalin. Water Treat. 2013, 51, 6355–6361. [Google Scholar] [CrossRef]
- Li, J.; Nickel, R.; Wu, J.; Lin, F.; van Lierop, J.; Liu, S. A new tool to attack biofilms: Driving magnetic iron-oxide nanoparticles to disrupt the matrix. Nanoscale 2019, 11, 6905–6915. [Google Scholar] [CrossRef]
- Hotrum, N.E.; de Jong, P.; Akkerman, J.C.; Fox, M.B. Pilot scale ultrasound enabled plate heat exchanger–its design and potential to prevent biofouling. J. Food Eng. 2015, 153, 81–88. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).