Early Intervention Using Fecal Microbiota Transplantation Combined with Probiotics Influence the Growth Performance, Diarrhea, and Intestinal Barrier Function of Piglets

 ,
,
Abstract
Featured Application
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Fecal Microbiota Suspension of Donor Pigs
2.2. Animals and Experimental Design
2.3. Transplantation of Fecal Microbiota Experiment and Animal Management
2.4. Determinations of Growth Performance and Diarrhea Rate
2.5. Sample Collections
2.6. Determination of Intestinal Barrier Biomarkers
2.7. Determinations of Fecal Short-Chain Fatty Acids and Fecal Succinate
2.8. DNA Extraction and Real-Time Quantitative Polymerase Chain Reaction (PCR)
2.9. Statistical Analysis
3. Results
3.1. Growth Performance and Diarrhea
3.2. Biomarkers of Intestinal Barrier Function
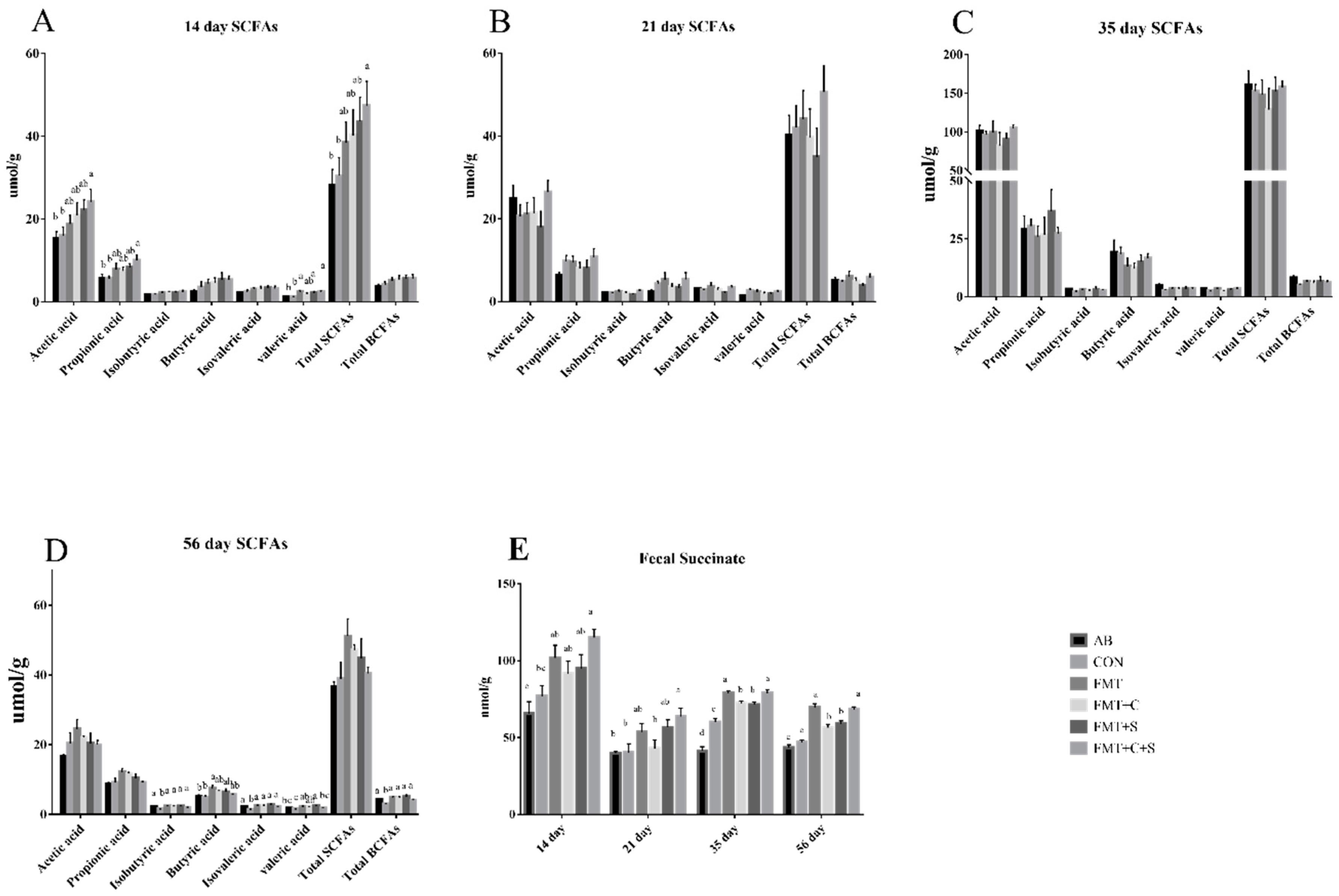
3.3. Fecal Short-Chain Fatty Acids and Fecal Succinate
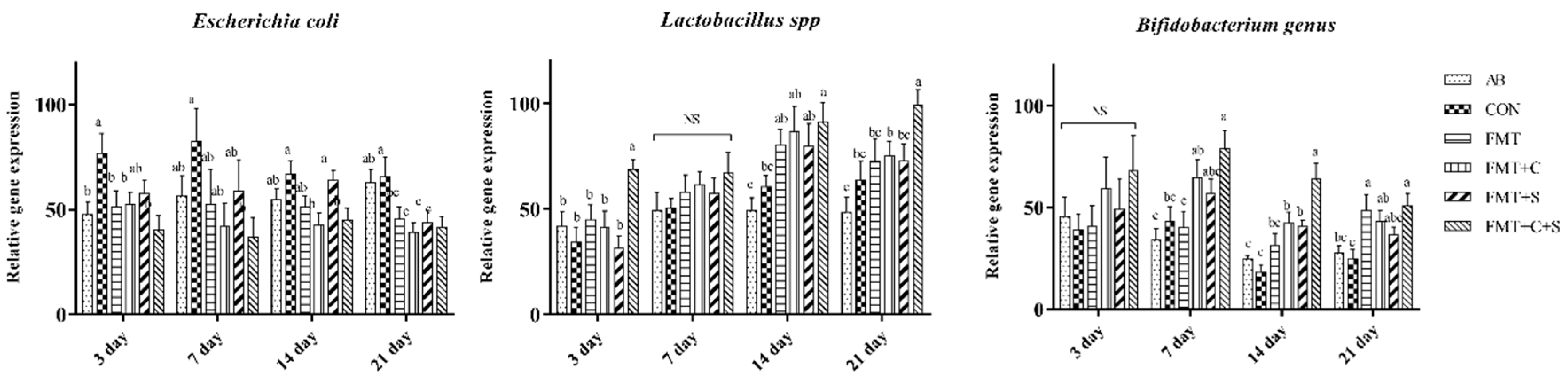
3.4. Selected Fecal Bacterial Populations in Suckling Period
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| FMT | Fecal Microbiota Transplantation |
| C. butyricum | Clostridium butyricum |
| S. boulardii | Saccharomyces boulardii |
| BW | body weight |
| ADG | average daily weight gain |
| ADFI | average daily feed intake |
| F:G | the ratio of ADFI and ADG |
| SCFAs | short-chain fatty acids |
| DAO | diamine oxidase |
| D-LA | D-lactate |
References
- Hu, J.; Ma, L.; Nie, Y.; Chen, J.; Zheng, W.; Wang, X.; Xie, C.; Zheng, Z.; Wang, Z.; Yang, T.; et al. A Microbiota-Derived Bacteriocin Targets the Host to Confer Diarrhea Resistance in Early-Weaned Piglets. Cell Host Microbe 2018, 24, 817–832. [Google Scholar] [CrossRef]
- Hu, L.; Geng, S.; Li, Y.; Cheng, S.; Fu, X.; Yue, X.; Han, X. Exogenous Fecal Microbiota Transplantation from Local Adult Pigs to Crossbred Newborn Piglets. Front. Microbiol. 2017, 8, 2663. [Google Scholar] [CrossRef] [PubMed]
- Kollmann, T.R.; Levy, O.; Montgomery, R.R.; Goriely, S. Innate Immune Function by Toll-like Receptors: Distinct Responses in Newborns and the Elderly. Immunity 2012, 37, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Kamada, N.; Seo, S.-U.; Chen, G.Y.; Núez, G. Role of the gut microbiota in immunity and inflammatory disease. Nat. Rev. Immunol. 2013, 13, 321–335. [Google Scholar] [CrossRef] [PubMed]
- Tanoue, T.; Atarashi, K.; Honda, K. Development and maintenance of intestinal regulatory T cells. Nat. Rev. Immunol. 2016, 16, 295. [Google Scholar] [CrossRef] [PubMed]
- Grover, M.; Kashyap, P.C. Germ-free mice as a model to study effect of gut microbiota on host physiology. Neurogastroenterol. Motil. 2014, 26, 745–748. [Google Scholar] [CrossRef]
- Cheng, C.S.; Wei, H.K.; Wang, P.; Yu, H.C.; Zhang, X.M.; Jiang, S.W.; Peng, J. Early intervention with faecal microbiota transplantation: An effective means to improve growth performance and the intestinal development of suckling piglets. Anim. Int. J. Anim. Biosci. 2018, 13, 533–541. [Google Scholar] [CrossRef]
- Kim, Y.-G.; Sakamoto, K.; Seo, S.-U.; Pickard, J.M.; Gillilland, M.G.; Pudlo, N.A.; Hoostal, M.; Li, X.; Wang, T.D.; Feehley, T. Neonatal acquisition of Clostridia species protects against colonization by bacterial pathogens. Science 2017, 356, 315. [Google Scholar] [CrossRef]
- Nguyen, Q.N.; Himes, J.E.; Martinez, D.R.; Permar, S.R. The Impact of the Gut Microbiota on Humoral Immunity to Pathogens and Vaccination in Early Infancy. PLoS Pathog. 2016, 12, e1005997. [Google Scholar] [CrossRef]
- Geng, S.; Cheng, S.; Li, Y.; Wen, Z.; Ma, X.; Jiang, X.; Wang, Y.; Han, X. Faecal Microbiota Transplantation Reduces Susceptibility to Epithelial Injury and Modulates Tryptophan Metabolism of the Microbial Community in a Piglet Model. J. Crohn’s Colitis 2018, 12, 1359–1374. [Google Scholar] [CrossRef]
- Fukuda, S.; Toh, H.; Hase, K.; Oshima, K.; Nakanishi, Y.; Yoshimura, K.; Tobe, T.; Clarke, J.M.; Topping, D.L.; Suzuki, T. Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 2012, 469, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Li, S.S.; Zhu, A.; Benes, V.; Costea, P.I.; Hercog, R.; Hildebrand, F.; Huerta-Cepas, J.; Nieuwdorp, M.; Salojarvi, J.; Voigt, A.Y.; et al. Durable coexistence of donor and recipient strains after fecal microbiota transplantation. Science 2016, 352, 586–589. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.Y.; Cisalpino, D.; Varadarajan, S.; Hellman, J.; Warren, H.S.; Cascalho, M.; Inohara, N.; Núñez, G. Gut Microbiota-Induced Immunoglobulin G Controls Systemic Infection by Symbiotic Bacteria and Pathogens. Immunity 2016, 44, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Li, G.; Zhang, X.; Wang, Y.; Wang, Y.; Wang, B.; Li, Y.; Li, W. Effect of Saccharomyces boulardii and Bacillus subtilis B10 on gut microbiota modulation in broilers. Anim. Nutr. 2018, 4, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.C.; Cara, D.C.; Fretez, S.H.; Cunha, F.Q.; Vieira, E.C.; Nicoli, J.R.; Vieira, L.Q. Saccharomyces boulardii stimulates sIgA production and the phagocytic system of gnotobiotic mice. J. Appl. Microbiol. 2010, 89, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, B.; Winckler, C.; Failing, K.; Breves, G. Studies on the Time Course of the Effects of the Probiotic YeastSaccharomyces boulardiion Electrolyte Transport in Pig Jejunum. Dig. Dis. Sci. 2004, 49, 1311–1317. [Google Scholar] [CrossRef]
- Li, M.; Zhu, L.; Xie, A.; Yuan, J. Oral Administration of Saccharomyces boulardiiAmeliorates Carbon Tetrachloride-Induced Liver Fibrosis in Rats via Reducing Intestinal Permeability and Modulating Gut Microbial Composition. Inflammation 2015, 38, 170–179. [Google Scholar] [CrossRef] [PubMed]
- Swidsinski, A.; Loening-Baucke, V.; Schulz, S.; Manowsky, J.; Verstraelen, H.; Swidsinski, S. Functional anatomy of the colonic bioreactor: Impact of antibiotics and Saccharomyces boulardii on bacterial composition in human fecal cylinders. Syst. Appl. Microbiol. 2016, 39, 67–75. [Google Scholar] [CrossRef]
- Jing, L.; Huang, H.; Lu, M.; Yong, Y.; Liu, S.; Ding, Y.; Bai, L.; Jie, J.; Zheng, P.; Gastroenterology, D.O. Effects of clostridium butyricum on intestinal barrier function in food allergic mice. Chongqing Med. 2017, 46, 3028–3032. [Google Scholar]
- Kanai, T.; Mikami, Y.; Hayashi, A. A breakthrough in probiotics: Clostridium butyricum regulates gut homeostasis and anti-inflammatory response in inflammatory bowel disease. J. Gastroenterol. 2015, 50, 928–939. [Google Scholar] [CrossRef]
- Li, H.H.; Li, Y.P.; Zhu, Q.; Qiao, J.Y.; Wang, W.J. Dietary supplementation with Clostridium butyricum helps to improve the intestinal barrier function of weaned piglets challenged with enterotoxigenic Escherichia coli K88. J. Appl. Microbiol. 2018, 125, 964–975. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Chen, X.; Liu, P.; Zhao, J.; Sun, J.; Guan, W.; Johnston, L.J.; Levesque, C.L.; Fan, P.; He, T.; et al. Dietary Clostridium butyricum Induces a Phased Shift in Fecal Microbiota Structure and Increases the Acetic Acid-Producing Bacteria in a Weaned Piglet Model. J. Agric. Food Chem. 2018, 66, 5157–5166. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Hua, X.; Qian, Y.; Ding, D.; Che, C.; Li, C.; Wei, J.; Bucheli, P.; Zhao, L. Inter-species transplantation of gut microbiota from human to pigs. ISME J. 2007, 1, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Hermann-Bank, M.L.; Skovgaard, K.; Stockmarr, A.; Strube, M.L.; Larsen, N.; Kongsted, H.; Ingerslev, H.C.; Molbak, L.; Boye, M. Characterization of the bacterial gut microbiota of piglets suffering from new neonatal porcine diarrhoea. BMC Vet. Res. 2015, 11, 139. [Google Scholar] [CrossRef]
- Koh, A.; Vadder, F.D.; Kovatcheva-Datchary, P.; Ckhed, F.B. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Macfarlane, S. Bacteria, Colonic Fermentation, and Gastrointestinal Health. J. Aoac Int. 2012, 95, 50–60. [Google Scholar] [CrossRef]
- Furusawa, Y.; Obata, Y.; Fukuda, S.; Endo, T.A.; Nakato, G.; Takahashi, D.; Nakanishi, Y.; Uetake, C.; Kato, K.; Kato, T. Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 2013, 504, 446. [Google Scholar] [CrossRef]
- Lin, M.Y.; De Zoete, M.R.; Van Putten, J.P.M.; Strijbis, K. Redirection of Epithelial Immune Responses by Short-Chain Fatty Acids through Inhibition of Histone Deacetylases. Front. Immunol. 2015, 6, 1500. [Google Scholar] [CrossRef]
- Schneider, C.; O’Leary, C.E.; von Moltke, J.; Liang, H.E.; Ang, Q.Y.; Turnbaugh, P.J.; Radhakrishnan, S.; Pellizzon, M.; Ma, A.; Locksley, R.M. A Metabolite-Triggered Tuft Cell-ILC2 Circuit Drives Small Intestinal Remodeling. Cell 2018, 174, 271–284. [Google Scholar] [CrossRef]
- Diao, H.; Jiao, A.R.; Yu, B.; He, J.; Yu, J.; Zheng, P.; Huang, Z.Q.; Luo, Y.H.; Luo, J.Q.; Mao, X.B. Stimulation of intestinal growth with distal ileal infusion of short-chain fatty acid: A reevaluation in a pig model. RSC Adv. 2017, 7, 30792–30806. [Google Scholar] [CrossRef]
- Fukudome, I.; Kobayashi, M.; Dabanaka, K.; Maeda, H.; Okamoto, K.; Okabayashi, T.; Baba, R.; Kumagai, N.; Oba, K.; Fujita, M. Diamine oxidase as a marker of intestinal mucosal injury and the effect of soluble dietary fiber on gastrointestinal tract toxicity after intravenous 5-fluorouracil treatment in rats. Med. Mol. Morphol. 2014, 47, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Rokkas, T.; Vaja, S.; Murphy, G.M.; Dowling, R.H. Postheparin plasma diamine oxidase in health and intestinal disease. Gastroenterology 1990, 98, 1493–1501. [Google Scholar] [CrossRef]
- Xun, W.; Shi, L.; Zhou, H.; Hou, G.; Cao, T.; Zhao, C. Effects of curcumin on growth performance, jejunal mucosal membrane integrity, morphology and immune status in weaned piglets challenged with enterotoxigenic Escherichia coli. Int. Immunopharmacol. 2015, 27, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Ho, S.B. Intestinal Goblet Cells and Mucins in Health and Disease: Recent Insights and Progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef]
- Chen, J.; Tellez, G.; Richards, J.D.; Escobar, J. Identification of Potential Biomarkers for Gut Barrier Failure in Broiler Chickens. Front. Vet. Sci. 2015, 2, 14. [Google Scholar] [CrossRef]
- Hartman, A.L.; Lough, D.M.; Barupal, D.K.; Fiehn, O.; Fishbein, T.; Zasloff, M.; Eisen, J.A. Human gut microbiome adopts an alternative state following small bowel transplantation. Proc. Natl. Acad. Sci. USA 2009, 106, 17187–17192. [Google Scholar] [CrossRef]
- Kushkevych, I.; Kotrsova, V.; Dordevic, D.; Bunkova, L.; Vitezova, M.; Amedei, A. Hydrogen Sulfide Effects on the Survival of Lactobacilli with Emphasis on the Development of Inflammatory Bowel Diseases. Biomolecules 2019, 9, 752. [Google Scholar] [CrossRef]
- Kushkevych, I.; Dordevic, D.; Kollar, P.; Vitezova, M.; Drago, L. Hydrogen Sulfide as a Toxic Product in the Small-Large Intestine Axis and its Role in IBD Development. J. Clin. Med. 2019, 8, 1054. [Google Scholar] [CrossRef]
- Jiao, J.; Wu, J.; Zhou, C.; Tang, S.; Wang, M.; Tan, Z. Composition of Ileal Bacterial Community in Grazing Goats Varies across Non-rumination, Transition and Rumination Stages of Life. Front. Microbiol. 2016, 7, 1364. [Google Scholar] [CrossRef]
- López, P.; Gueimonde, M.; Margolles, A.; Suárez, A. Distinct Bifidobacterium strains drive different immune responses in vitro. Int. J. Food Microbiol. 2010, 138, 157–165. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti–PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [PubMed]
- Menard, O.; Butel, M.-J.; Gaboriau-Routhiau, V.; Waligora-Dupriet, A.-J. Gnotobiotic Mouse Immune Response Induced by Bifidobacterium sp. Strains Isolated from Infants. Appl. Environ. Microbiol. 2008, 74, 660–666. [Google Scholar] [CrossRef] [PubMed]
- Ashiruoredope, D.; Hopkins, S. Antimicrobial resistance: Moving from professional engagement to public action. J. Antimicrob. Chemother. 2015, 70, 2927–2930. [Google Scholar] [CrossRef] [PubMed]
- Russell, S.L.; Gold, M.J.; Willing, B.P.; Thorson, L.; McNagny, K.M.; Finlay, B.B. Perinatal antibiotic treatment affects murine microbiota, immune responses and allergic asthma. Gut Microbes 2013, 4, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Vangay, P.; Ward, T.; Gerber, J.S.; Knights, D. Antibiotics, Pediatric Dysbiosis, and Disease. Cell Host Microbe 2015, 17, 553–564. [Google Scholar] [CrossRef] [PubMed]
- Vijay-Kumar, M.; Aitken, J.D.; Carvalho, F.A.; Cullender, T.C.; Mwangi, S.; Srinivasan, S.; Sitaraman, S.V.; Knight, R.; Ley, R.E.; Gewirtz, A.T. Metabolic Syndrome and Altered Gut Microbiota in Mice Lacking Toll-Like Receptor 5. Science 2010, 328, 228–231. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Target Group | Sequence of Primers (5′ to 3′) | Size (bp) | Annealing Temperature (°C) | Reference |
|---|---|---|---|---|
| Total bacteria | Forward: ACTCCTACGGGAGGCAGCAG | 175 | 60 | Cheng et al., 2018 |
| Reverse: ATTACCGCGGCTGCTGG | ||||
| Lactobacillus spp. | Forward: CACCGCTACACATGGAG | 341 | 58 | Cheng et al., 2018 |
| Reverse: TGGAAGATTCCCTACTGCT | ||||
| Escherichia coli | Forward: CATGCCGCGTGTATGAAGAA | 96 | 60 | Huijsdens et al., 2002 |
| Reverse: TTTGCTCATTGACGTTACCCG | ||||
| Reverse: AATTCCGCCTACCTCTGCACT | ||||
| Bifidobacterium genus | Forward: TCGCGTC(C/T)GGTGTGAAAG | 243 | 58 | Rinttila et al., 2004 |
| Reverse: CCACATCCAGC(A/G)TCCAC |
| Phases | Item | Treatments 1 | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|
| Control Groups | Treatment Groups | ||||||||
| AB | CON | FMT | FMT+C | FMT+S | FMT+C+S | ||||
| Suckling period (Days 1–27) | Piglets | 132 | 84 | 157 | 149 | 112 | 106 | ||
| Sows | 13 | 10 | 15 | 15 | 12 | 12 | |||
| BW 2, kg | |||||||||
| Day 1 | 1.55 | 1.57 | 1.55 | 1.53 | 1.55 | 1.54 | 0.01 | 0.9681 | |
| Day 7 | 2.52 | 2.46 | 2.57 | 2.48 | 2.6 | 2.57 | 0.02 | 0.2404 | |
| Day 14 | 4.11 a,b | 4.01 b | 4.31 a | 4.17 a,b | 4.25 a | 4.26 a | 0.03 | 0.0762 | |
| Day 21 | 5.59 b,c | 5.51 c | 5.87 a,b | 5.76 b,c | 5.83 a,b | 6.11 a | 0.03 | 0.0017 | |
| Day 27 | 6.88 b,c | 6.85 c | 7.26 a,b | 6.96 b,c | 7.14 a,b,c | 7.51 a | 0.04 | 0.0028 | |
| ADG 3, g/d | |||||||||
| Days 1–7 | 161.15 | 151.79 | 164 | 156.9 | 167.71 | 162.63 | 1.74 | 0.3105 | |
| Days 8–14 | 227.37 b,c | 220.66 c | 247.81 a | 241.51 a,b | 235.31 a,b,c | 242.4 a,b | 2.17 | 0.0254 | |
| Days 15–21 | 211.24 b | 214.13 b | 223.64 b | 226.26 b | 226.12 b | 255.46 a | 2.56 | 0.0007 | |
| Days 22–27 | 214.80 b,c | 223.29 a,b,c | 230.57 a,b | 200.49 c | 219.17 b,c | 243.40 a | 2.62 | 0.0030 | |
| Days 1–27 | 204.85 b | 203.62 b | 217.99 a,b | 208.42 b | 213.51 b | 227.74 a | 1.39 | 0.0032 | |
| Diarrhea rate, % | |||||||||
| Days 1–7 | 10.41 | 13.97 | 7.89 | 7.34 | 11.21 | 5.93 | 1.11 | 0.3757 | |
| Days 8–14 | 5.14 a | 4.27 a,b | 2.81 b,c | 1.98 c | 3.76 a,b,c | 1.96 c | 0.33 | 0.0182 | |
| Days 15–21 | 8.95 a,b | 11.66 a | 7.53 b | 6.27 b | 7.45 b | 5.14 b | 0.58 | 0.0144 | |
| Days 22–27 | 4.93 | 4.93 | 4.93 | 4.93 | 4.93 | 4.93 | 0.35 | 0.6937 | |
| Days 1–27 | 7.47 a,b | 9.33 a | 5.92 b,c | 5.16 b,c | 7.08 a,b,c | 4.49 c | 0.38 | 0.0047 | |
| Diarrhea index | |||||||||
| Days 1–7 | 0.52 | 0.6 | 0.47 | 0.47 | 0.58 | 0.46 | 0.09 | 0.9979 | |
| Days 8–14 | 0.11 a | 0.1 a,b | 0.07 a,b | 0.06 a,b | 0.09 a,b | 0.05 b | 0.01 | 0.0830 | |
| Days 15–21 | 0.17 b | 0.27 a | 0.18 b | 0.15 b | 0.17 b | 0.12 b | 0.01 | 0.0202 | |
| Days 22–27 | 0.12 | 0.16 | 0.13 | 0.13 | 0.15 | 0.14 | 0.01 | 0.8997 | |
| Days 1–27 | 0.23 | 0.29 | 0.22 | 0.21 | 0.25 | 0.19 | 0.03 | 0.9388 | |
| Post-weaning period (Days 28–63) | Piglets | 81 | 71 | 83 | 83 | 84 | 78 | ||
| Pans | 4 | 4 | 4 | 4 | 4 | 4 | |||
| BW, kg | |||||||||
| Day 27 | 6.79 c | 7.25 b | 7.78 a | 7.17 b | 7.37 b | 7.83 a | 0.06 | <0.0001 | |
| Day 35 | 7.47 d | 7.89 c | 8.30 b | 7.79 c | 7.94 c | 8.49 a | 0.06 | <0.0001 | |
| Day 42 | 10.27 c | 10.75 b,c | 11.13 b | 10.96 b | 10.65 b,c | 11.79 c | 0.08 | <0.0001 | |
| Day 49 | 13.87 d | 14.45 c | 15.26 b | 14.67 c | 14.46 c | 16.12 a | 0.10 | <0.0001 | |
| Day 56 | 18.60 d | 18.92 c,d | 19.62 b | 19.34 b,c | 18.95 c,d | 21.01 a | 0.12 | <0.0001 | |
| Day 63 | 23.66 c | 23.92 c | 25.07 b | 24.36 b,c | 23.79 c | 26.54 a | 0.15 | <0.0001 | |
| ADG, g/d | |||||||||
| Days 27–35 | 96.33 | 90.54 | 73.79 | 89.02 | 80.64 | 94.49 | 3.53 | 0.3333 | |
| Days 36–42 | 400.77 b | 405.71 b | 405.71 b | 456.10 a | 393.00 b | 462.54 a | 4.80 | <0.0001 | |
| Days 43–49 | 503.09 d | 529.34 c,d | 587.14 a,b | 527.21 c,d | 552.67 b,c | 617.25 a | 5.86 | <0.0001 | |
| Days 50–56 | 682.17 a,b | 634.53 b,c | 622.79 c | 667.30 a,b,c | 642.52 b,c | 698.55 a | 6.54 | 0.0254 | |
| Days 57–63 | 723.05 a,b | 726.24 a,b | 777.62 a | 717.23 a,b | 687.31 b | 789.36 a | 8.73 | 0.0211 | |
| Days 27–63 | 481.27 b | 476.74 b | 494.37 b | 490.71 b | 471.06 b | 530.74 a | 3.47 | <0.0001 | |
| ADFI 4, g/d | |||||||||
| Days 27–35 | 148.91 | 153.47 | 146.9 | 147.9 | 134.95 | 143.56 | 2.36 | 0.3301 | |
| Days 36–42 | 466.58 | 472.92 | 475.98 | 486.33 | 488.87 | 520.17 | 6.7 | 0.2404 | |
| Days 43–49 | 718.82 | 759.39 | 776.59 | 766.47 | 756.98 | 847.84 | 13.22 | 0.1060 | |
| Days 50–56 | 966.53 | 937.12 | 956.86 | 967.15 | 929.34 | 1031.81 | 14.42 | 0.4080 | |
| Days 57–63 | 1155.95 b,c | 1176.29 b,c | 1237.52 a,b | 1150.79 b,c | 1108.41 c | 1290.33 a | 17.98 | 0.0235 | |
| Days 27–63 | 693.72 | 701.96 | 720.38 | 707.02 | 687.47 | 769.76 | 9.52 | 0.1284 | |
| Diarrhea rate, % | |||||||||
| Days 27–35 | 6.35 b | 9.78 a,b | 12.16 a | 7.25 b | 9.69 a,b | 5.89 b | 0.64 | 0.0209 | |
| Days 36–42 | 3.48 a | 6.60 a | 6.12 a,b | 4.59 b,c | 4.30 b,c | 2.59 c | 0.33 | 0.0007 | |
| Days 43–49 | 4.34 a,b | 5.87 a | 3.48 a,b | 3.04 b | 4.48 a,b | 2.04 b | 0.36 | 0.0374 | |
| Days 50–56 | 2.21 b,c | 5.56 a | 3.53 b | 1.99 b,c | 2.41 b,c | 1.12 c | 0.3 | <0.0001 | |
| Days 57–63 | 2.08 b | 5.56 a | 4.42 a | 1.03 b | 2.41 b | 1.05 b | 0.37 | <0.0001 | |
| Days 27–63 | 3.75 c,d | 6.67 a | 5.96 a,b | 3.69 c,d | 4.67 b,c | 2.60 d | 0.23 | <0.0001 | |
| Diarrhea index | |||||||||
| Days 27–35 | 0.17 | 0.24 | 0.29 | 0.22 | 0.27 | 0.18 | 0.02 | 0.1802 | |
| Days 36–42 | 0.10 b,c | 0.17 a | 0.15 a,b | 0.12 b,c | 0.13 a,b,c | 0.08 c | 0.01 | 0.0039 | |
| Days 43–49 | 0.10 a,b | 0.13 a | 0.09 a,b | 0.08 a,b | 0.12 a | 0.05 b | 0.01 | 0.0367 | |
| Days 50–56 | 0.06 b,c | 0.12 a | 0.08 b | 0.04 c | 0.06 b,c | 0.04 c | 0.01 | 0.0002 | |
| Days 57–63 | 0.05 b | 0.13 a | 0.11 a | 0.03 b | 0.06 b | 0.03 b | 0.01 | <0.0001 | |
| Days 27–63 | 0.10 c,d | 0.16 a | 0.14 a,b | 0.10 b,c,d | 0.13 a,b,c | 0.08 d | 0.01 | 0.0003 | |
| Item | Treatment | SEM | p-Value | |||||
|---|---|---|---|---|---|---|---|---|
| Control Groups | Treatment Groups | |||||||
| AB | CON | FMT | FMT+C | FMT+S | FMT+C+S | |||
| Day 14 | ||||||||
| Plasma DAO, ng/mL | 207.2 a | 190.38 a,b | 193.89 a,b | 173.05 b,c | 189.81 a,b | 161.41 c | 3.34 | 0.0007 |
| Plasma D-LA, umol/mL | 180.08 a,b | 149.35 b,c | 208.71 a | 147.14 b,c | 204.02 a | 122.80 c | 8.04 | 0.0079 |
| Fecal MUC2, ng/g | 268.07b | 322.24 b | 443.13 a | 455.90 a | 412.03 a | 479.72 a | 15.18 | <0.0001 |
| Day 21 | ||||||||
| Plasma DAO, ng/mL | 175.87 a,b | 194.63 a | 162.95 b | 165.39 b | 124.43 c | 118.29 c | 6.29 | <0.0001 |
| Plasma D-LA, umol/mL | 389.47 a,b | 438.17 a | 343.40 b | 268.90 c | 222.59 c | 229.70 c | 18.19 | <0.0001 |
| Fecal MUC2, ng/g | 237.70c | 301.31 b | 375.16 a | 425.41 a | 425.37 a | 416.82 a | 16.24 | <0.0001 |
| Day 35 | ||||||||
| Plasma DAO, ng/mL | 166.59 a | 177.94 a | 102.02 d | 152.17 b | 130.41 c | 103.81 d | 3.85 | <0.0001 |
| Plasma D-LA, umol/mL | 335.19 a | 360.20 a | 184.47 d | 302.31 b | 251.76 c | 190.56 d | 9.20 | <0.0001 |
| Fecal MUC2, ng/g | 197.46 d | 343.40 c | 472.95 a | 414.30 b | 398.62 b | 489.34 a | 12.94 | <0.0001 |
| Day 56 | ||||||||
| Plasma DAO, ng/mL | 179.73 a | 153.02 b | 146.03 b | 117.26 c | 94.41 d | 92.58 d | 4.09 | <0.0001 |
| Plasma D-LA, umol/mL | 338.88 a | 326.82 a | 272.74 b | 242.06 c | 166.48 d | 178.75 d | 8.66 | <0.0001 |
| Fecal MUC2, ng/g | 221.01 d | 263.35 c | 364.45 a | 318.08 b | 310.81 b | 371.43 a | 8.47 | <0.0001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, Q.; Wu, X.; Pan, Y.; Wang, L.; Guo, Y.; Cui, C.; Hu, L.; Zhu, L.; Peng, J.; Wei, H. Early Intervention Using Fecal Microbiota Transplantation Combined with Probiotics Influence the Growth Performance, Diarrhea, and Intestinal Barrier Function of Piglets. Appl. Sci. 2020, 10, 568. https://doi.org/10.3390/app10020568
Xiang Q, Wu X, Pan Y, Wang L, Guo Y, Cui C, Hu L, Zhu L, Peng J, Wei H. Early Intervention Using Fecal Microbiota Transplantation Combined with Probiotics Influence the Growth Performance, Diarrhea, and Intestinal Barrier Function of Piglets. Applied Sciences. 2020; 10(2):568. https://doi.org/10.3390/app10020568
Chicago/Turabian StyleXiang, Quanhang, Xiaoyu Wu, Ye Pan, Liu Wang, Yuwei Guo, Chenbin Cui, Lingling Hu, Lingling Zhu, Jian Peng, and Hongkui Wei. 2020. "Early Intervention Using Fecal Microbiota Transplantation Combined with Probiotics Influence the Growth Performance, Diarrhea, and Intestinal Barrier Function of Piglets" Applied Sciences 10, no. 2: 568. https://doi.org/10.3390/app10020568
APA StyleXiang, Q., Wu, X., Pan, Y., Wang, L., Guo, Y., Cui, C., Hu, L., Zhu, L., Peng, J., & Wei, H. (2020). Early Intervention Using Fecal Microbiota Transplantation Combined with Probiotics Influence the Growth Performance, Diarrhea, and Intestinal Barrier Function of Piglets. Applied Sciences, 10(2), 568. https://doi.org/10.3390/app10020568