Latitudinal Variation in Circadian Rhythmicity in Nasonia vitripennis

 , , ,
, , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Lines
2.2. Locomotor Activity Recording
2.3. Data Analysis and Statistics
3. Results
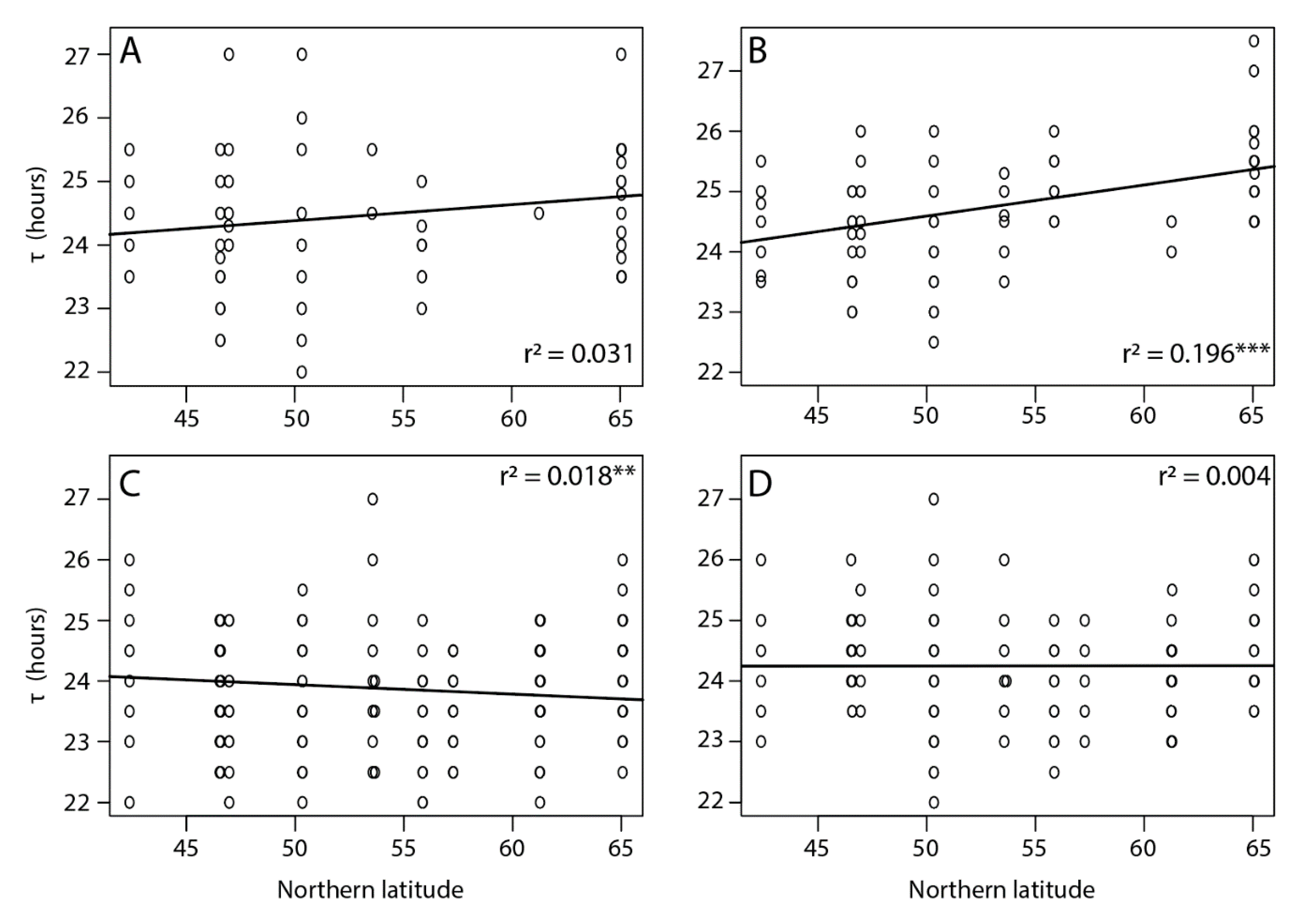
3.1. Rhythmicity and Free Running Periods (τ)
3.2. Activity Timing
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pittendrigh, C.S.; Kyner, W.T.; Takamura, T. The Amplitude of circadian oscillations: Temperature dependence, latitudinal clines, and the photoperiodic time measurement. J. Biol. Rhythm. 1991, 6, 299–313. [Google Scholar] [CrossRef]
- Bradshaw, W.E.; Holzapfel, C.M. Evolution of animal photoperiodism. Annu. Rev. Ecol. Evol. Syst. 2007, 38, 1–25. [Google Scholar] [CrossRef]
- Bunning, E. Circadian rhythms and the mime measurement in photoperiodism. Cold Spring Harb. Symp. Quant. Biol. 1960, 25, 249–256. [Google Scholar] [CrossRef]
- Saunders, D.S. Controversial aspects of photoperiodism in insects and mites. J. Insect Physiol. 2010, 56, 1491–1502. [Google Scholar] [CrossRef] [PubMed]
- Koštál, V. Insect photoperiodic calendar and circadian clock: Independence, cooperation, or unity? J. Insect Physiol. 2011, 57, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Mukai, A.; Goto, S.G. The clock gene period is essential for the photoperiodic response in the jewel wasp Nasonia vitripennis (Hymenoptera: Pteromalidae). Appl. Entomol. Zool. 2016, 51, 185–194. [Google Scholar] [CrossRef]
- Urbanová, V.; Bazalová, O.; Vaněčková, H.; Dolezel, D. Photoperiod regulates growth of male accessory glands through juvenile hormone signaling in the linden bug, Pyrrhocoris apterus. Insect Biochem. Mol. Biol. 2016, 70, 184–190. [Google Scholar] [CrossRef]
- Dalla Benetta, E.; Beukeboom, L.W.; van de Zande, L. Adaptive differences in circadian clock gene expression patterns and photoperiodic diapause induction in Nasonia vitripennis. Am. Nat. 2019, 193, 881–896. [Google Scholar] [CrossRef]
- Hut, R.A.; Paolucci, S.; Dor, R.; Kyriacou, C.P.; Daan, S. Latitudinal clines: An evolutionary view on biological rhythms. Proc. R. Soc. B 2013, 280, 20130433. [Google Scholar] [CrossRef]
- Hut, R.A.; Beersma, D.G.M. Evolution of time-keeping mechanisms: Early emergence and adaptation to photoperiod. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2141–2154. [Google Scholar] [CrossRef]
- Paolucci, S.; van de Zande, L.; Beukeboom, L.W. Adaptive latitudinal cline of photoperiodic diapause induction in the parasitoid Nasonia vitripennis in Europe. J. Evol. Biol. 2013, 26, 705–718. [Google Scholar] [CrossRef] [PubMed]
- Saunders, D.S. Insect photoperiodism: measuring the night. J. Insect Physiol. 2013, 59, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Saunders, D.S. Photoperiodism and time measurement in the parasitic wasp, Nasonia vitripennis. J. Insect Physiol. 1968, 14, 433–450. [Google Scholar] [CrossRef]
- Paolucci, S.; Salis, L.; Vermeulen, C.J.; Beukeboom, L.W.; van de Zande, L. QTL analysis of the photoperiodic response and clinal distribution of period alleles in Nasonia vitripennis. Mol. Ecol. 2016, 25, 4805–4817. [Google Scholar] [CrossRef]
- Schmid, B.; Helfrich-Förster, C.; Yoshii, T. A new ImageJ plug-in “ActogramJ” for chronobiological analyses. J. Biol. Rhythm. 2011, 26, 464–467. [Google Scholar] [CrossRef]
- Sokolove, P.G.; Bushell, W.N. The chi square periodogram: Its utility for analysis of circadian rhythms. J. Theor. Biol. 1978, 72, 131–160. [Google Scholar] [CrossRef]
- Schlichting, M.; Helfrich-Förster, C. Photic entrainment in Drosophila assessed by locomotor activity recordings. Methods Enzymol. 2015, 552, 105–123. [Google Scholar]
- Floessner, T.S.E.; Boekelman, F.E.; Druiven, S.J.M.; de Jong, M.; Rigter, P.M.F.; Beersma, D.G.M.; Hut, R.A. Lifespan is unaffected by size and direction of daily phase shifts in Nasonia, a hymenopteran insect with strong circadian light resetting. J. Insect Physiol. 2019, 117, 103896. [Google Scholar] [CrossRef]
- Bertossa, R.C.; van Dijk, J.; Diao, W.; Saunders, D.; Beukeboom, L.W.; Beersma, D.G.M. Circadian rhythms differ between sexes and closely related species of Nasonia wasps. PLoS ONE 2013, 8, e60167. [Google Scholar] [CrossRef]
- Sharma, V.; Lone, S.; Goel, A.; Chandrashekaran, M.K. Circadian consequences of social organization in the ant species Camponotus compressus. Naturwissenschaften 2004, 91, 386–390. [Google Scholar] [CrossRef]
- Kauranen, H.; Menegazzi, P.; Costa, R.; Helfrich-Förster, C.; Kankainen, A.; Hoikkala, A. Flies in the north: Locomotor behavior and clock neuron organization of Drosophila montana. J. Biol. Rhythm. 2012, 27, 377–387. [Google Scholar] [CrossRef]
- Prabhakaran, P.M.; Sheeba, V. Sympatric Drosophilid species melanogaster and ananassae differ in temporal patterns of activity. J. Biol. Rhythm. 2012, 27, 365–376. [Google Scholar] [CrossRef] [PubMed]
- Prabhakaran, P.M.; Sheeba, V. Insights into differential activity patterns of drosophilids under semi-natural conditions. J. Exp. Biol. 2013, 216, 4691–4702. [Google Scholar] [CrossRef] [PubMed]
- Michael, T.P. Enhanced fitness conferred by naturally occurring variation in the circadian clock. Science 2003, 302, 1049–1053. [Google Scholar] [CrossRef]
- Shinkawa, Y.; Takeda, S.; Tomioka, K.; Matsumoto, A.; Oda, T.; Chiba, Y. Variability in circadian activity patterns within the Culex pipiens complex (Diptera: Culicidae). J. Med. Entomol. 1994, 31, 49–56. [Google Scholar] [CrossRef]
- Pivarciova, L.; Vaneckova, H.; Provaznik, J.; Wu, B.C.-H.; Pivarci, M.; Peckova, O.; Bazalova, O.; Cada, S.; Kment, P.; Kotwica-Rolinska, J.; et al. Unexpected geographic variability of the free running period in the linden bug Pyrrhocoris apterus. J. Biol. Rhythm. 2016, 31, 568–576. [Google Scholar] [CrossRef]
- Helfrich-Förster, C.; Bertolini, E.; Menegazzi, P. Flies as models for circadian clock adaptation to environmental challenges. Eur. J. Neurosci. 2018. [Google Scholar] [CrossRef]
- Majercak, J.; Chen, W.F.; Edery, I. Splicing of the period gene 3′-terminal intron is regulated by light, circadian clock factors, and phospholipase C. Mol. Cell Biol. 2004, 24, 3359–3372. [Google Scholar] [CrossRef]
- Low, K.H.; Lim, C.; Ko, H.W.; Edery, I. Natural variation in the splice site strength of a clock gene and species-specific thermal adaptation. Neuron 2008, 60, 1054–1067. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paolucci, S.; Dalla Benetta, E.; Salis, L.; Doležel, D.; van de Zande, L.; Beukeboom, L.W. Latitudinal Variation in Circadian Rhythmicity in Nasonia vitripennis. Behav. Sci. 2019, 9, 115. https://doi.org/10.3390/bs9110115
Paolucci S, Dalla Benetta E, Salis L, Doležel D, van de Zande L, Beukeboom LW. Latitudinal Variation in Circadian Rhythmicity in Nasonia vitripennis. Behavioral Sciences. 2019; 9(11):115. https://doi.org/10.3390/bs9110115
Chicago/Turabian StylePaolucci, Silvia, Elena Dalla Benetta, Lucia Salis, David Doležel, Louis van de Zande, and Leo W. Beukeboom. 2019. "Latitudinal Variation in Circadian Rhythmicity in Nasonia vitripennis" Behavioral Sciences 9, no. 11: 115. https://doi.org/10.3390/bs9110115
APA StylePaolucci, S., Dalla Benetta, E., Salis, L., Doležel, D., van de Zande, L., & Beukeboom, L. W. (2019). Latitudinal Variation in Circadian Rhythmicity in Nasonia vitripennis. Behavioral Sciences, 9(11), 115. https://doi.org/10.3390/bs9110115