Atypical Processing of Novel Distracters in a Visual Oddball Task in Autism Spectrum Disorder


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods and Materials
2.1. Participants
2.2. ERP Data Acquisition, and Signal Processing
2.3. Three Stimuli Visual Oddball Test with Novel Distracters
2.4. Behavioral Measures
2.5. Event-Related Potentials (ERP)
2.5.1. Stimulus-Locked ERPs
2.5.2. Response-Locked Dependent Variables (ERN/Pe)
2.6. Statistical Data Analysis
3. Results
3.1. Behavioral Responses
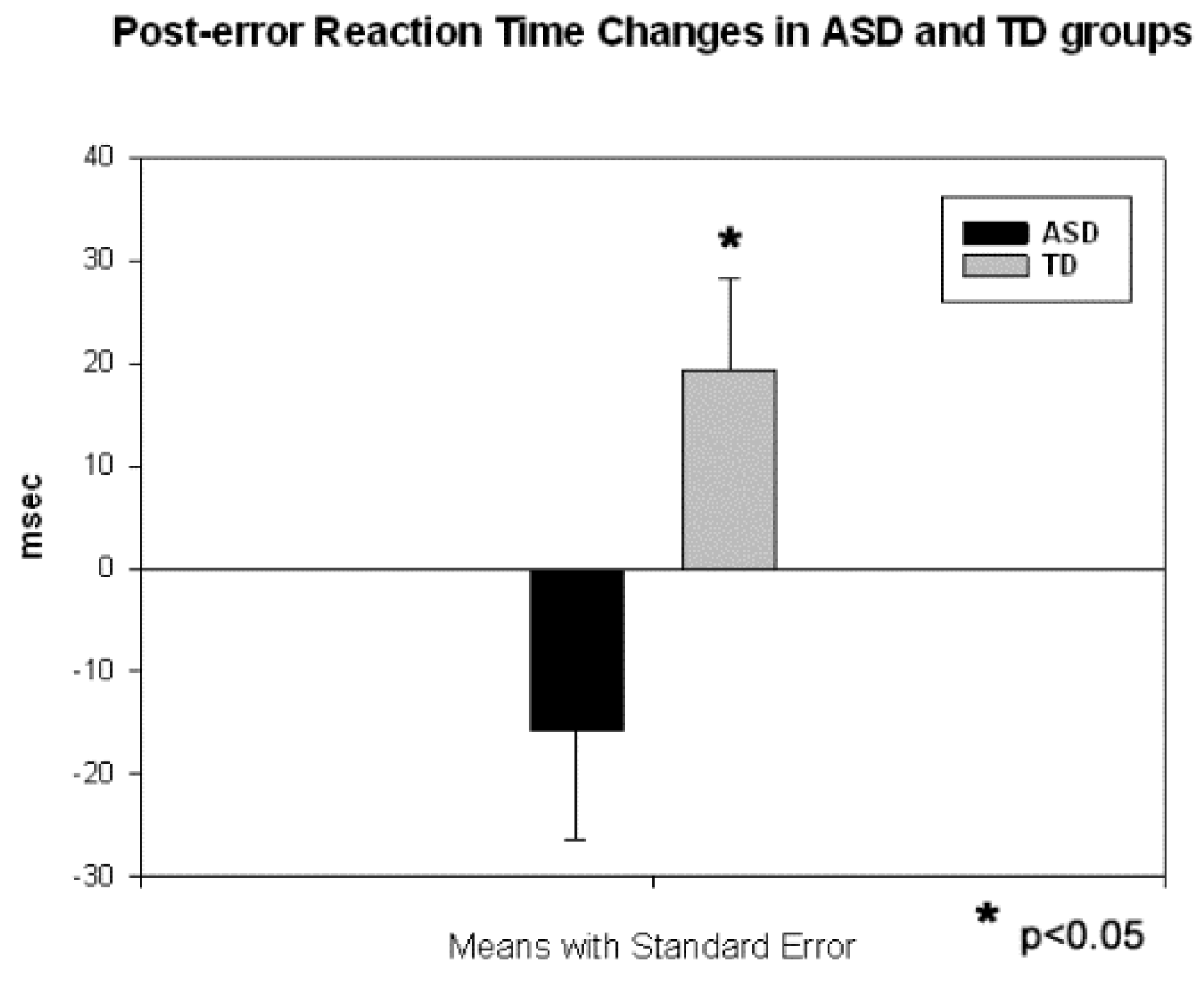
3.2. Motor Response-Locked Fronto-Central ERPs
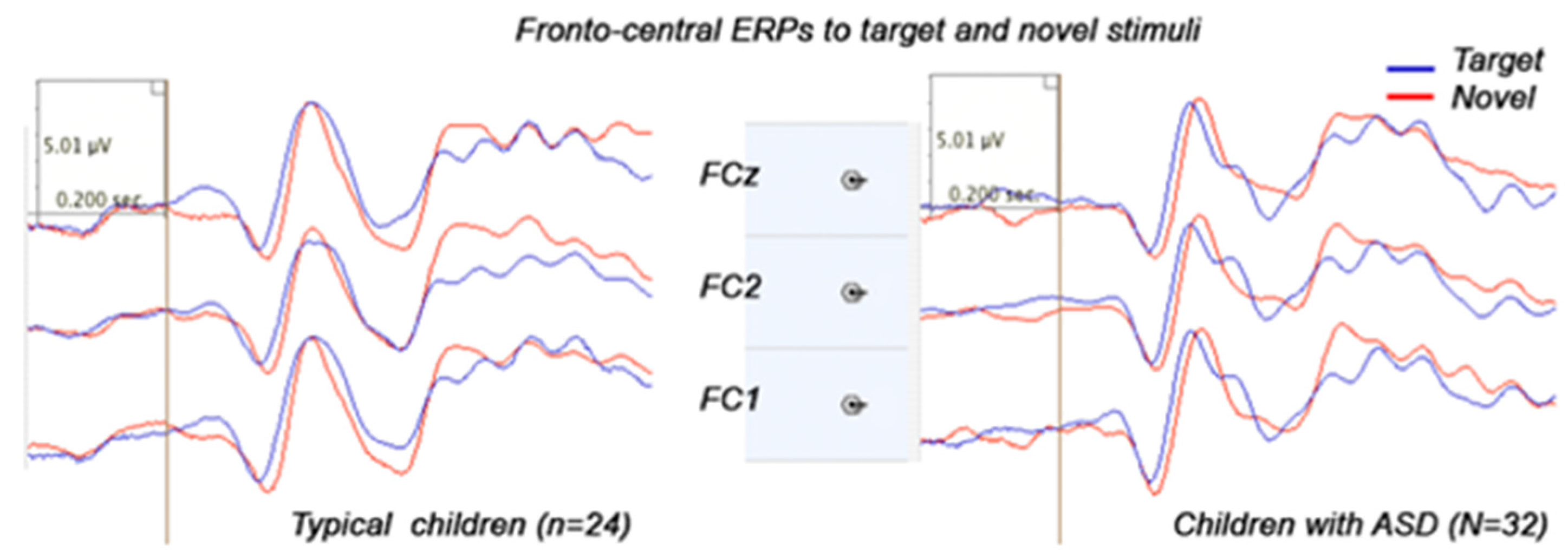
3.3. Stimulus-Locked Event-Related Potentials
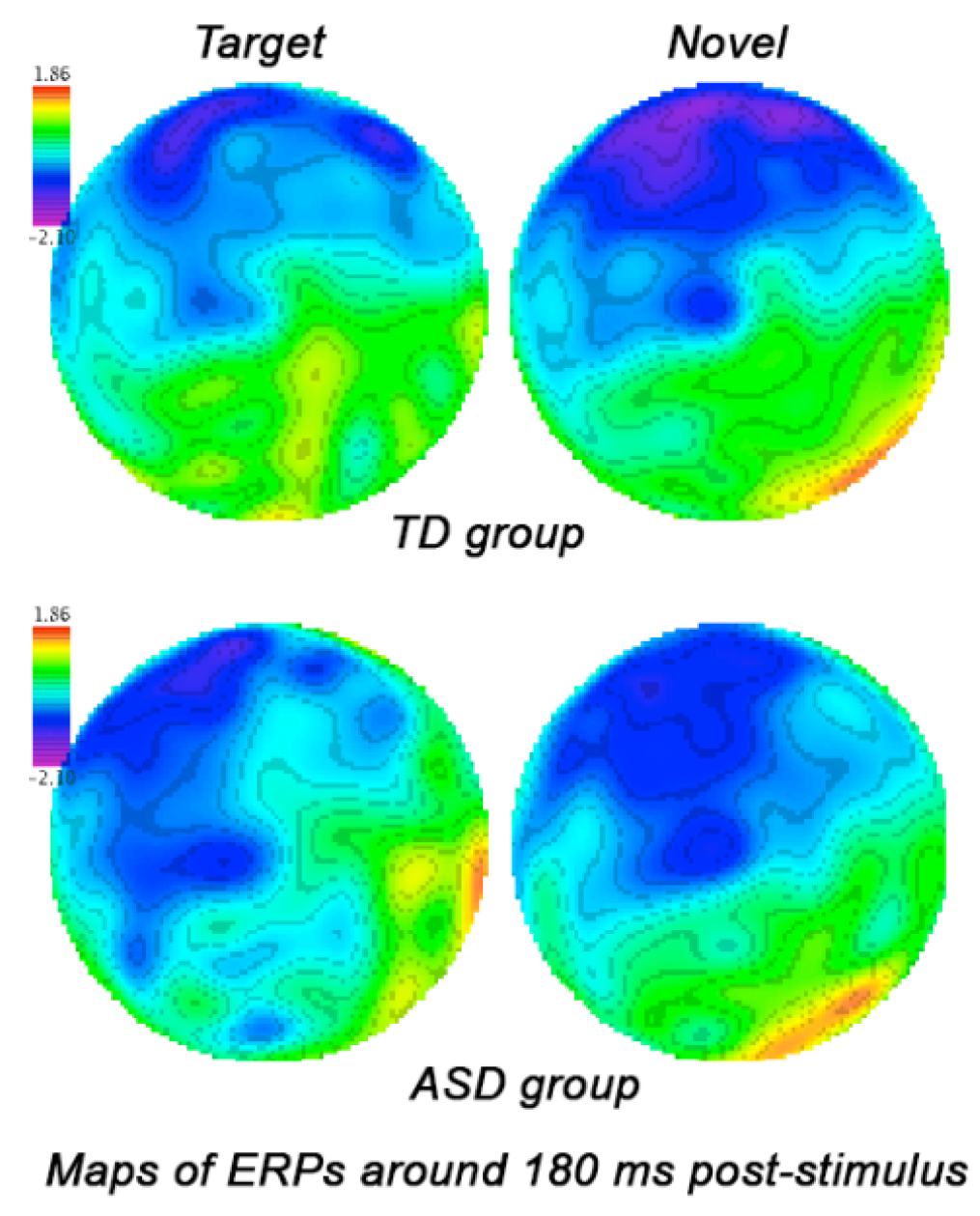
3.3.1. Frontal ERPs
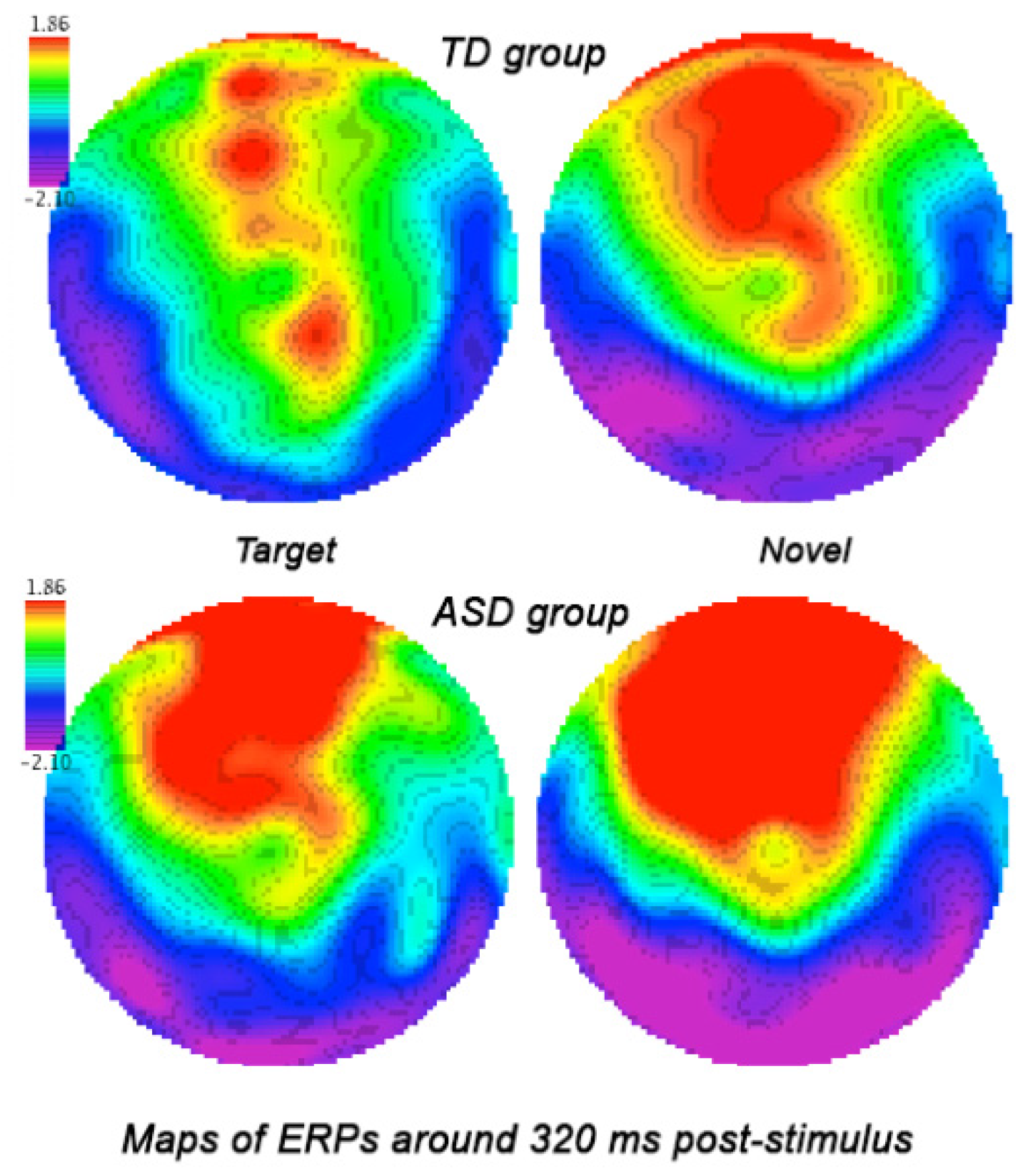
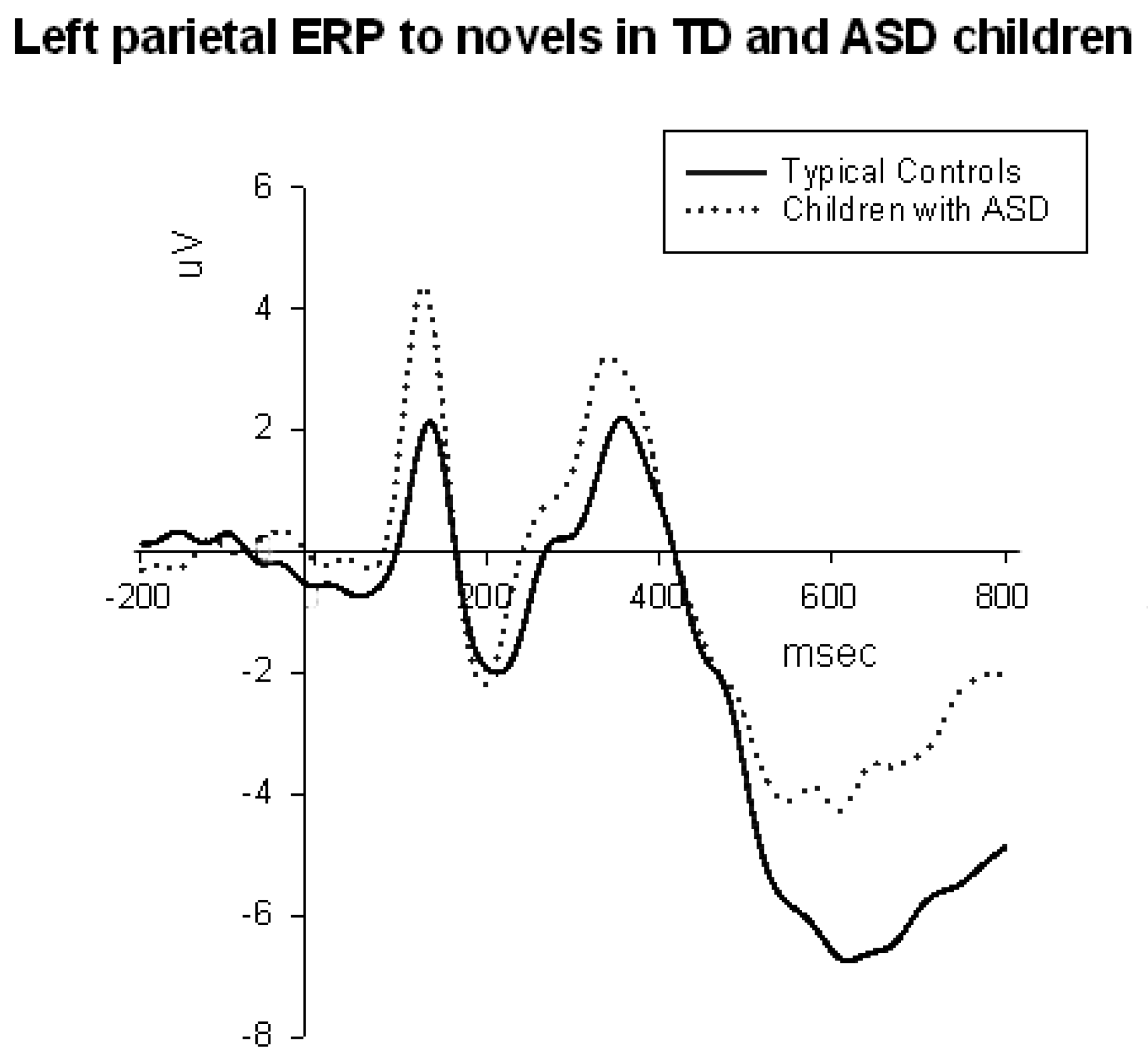
3.3.2. Parietal and Parieto-Occipital ERPs
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Diagnostic and Statistical Manual of Mental Disorders (DSM-V), 5th ed.; American Psychiatric Publishing, Inc.: Washington, DC, USA, 2013; ISBN 089042554X.
- Gomes, E.; Pedroso, F.S.; Wagner, M.B. Auditory hypersensitivity in the autistic spectrum disorder. Pro Fono 2008, 20, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Baruth, J.M.; Casanova, M.F.; Sears, L.; Sokhadze, E. Early-stage visual processing abnormalities in autism spectrum disorder (ASD). Transl. Neurosci. 2010, 1, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.F.; Baruth, J.M.; El-Baz, A.; Tasman, A.; Sears, L.; Sokhadze, E. Repetitive transcranial magnetic stimulation (rTMS) modulates event-related potential (ERP) indices of attention in autism. Transl. Neurosci. 2012, 3, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Sokhadze, E.; Baruth, J.; Tasman, A.; Sears, L.; Mathai, G.; El-Baz, A.; Casanova, M.F. Event-related potential study of novelty processing abnormalities in autism. Appl. Psychophysiol. Biofeedback 2009, 34, 37–51. [Google Scholar] [CrossRef] [PubMed]
- Sokhadze, E.; Baruth, J.; Tasman, A.; Mansoor, M.; Ramaswamy, R.; Sears, L.; Mathai, G.; El-Baz, A.; Casanova, M.F. Low-frequency repetitive transcranial magnetic stimulation (rTMS) affects event-related potential measures of novelty processing in autism. Appl. Psychophysiol. Biofeedback 2010, 35, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Sokhadze, E.; Baruth, J.; El-Baz, A.; Horrell, T.; Sokhadze, G.; Carroll, T.; Tasman, A.; Sears, L.; Casanova, M. Impaired error monitoring and correction function in autism. J. Neurother. 2010, 14, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Sokhadze, E.M.; Baruth, J.M.; Sears, L.; Sokhadze, G.E.; El-Baz, A.S.; Williams, E.; Klapheke, R.; Casanova, M.F. Event related potentials study of attention regulation during illusory figure categorization task in ADHD, autism spectrum disorders, and typical children. J. Neurother. 2012, 16, 12–31. [Google Scholar] [CrossRef] [PubMed]
- Sokhadze, E.M.; Baruth, J.; Tasman, A.; Casanova, M.F. Event-related potential studies of cognitive processing abnormalities in autism. In Imaging the Brain in Autism; Casanova, M.F., El-Baz, A.S., Suri, J.S., Eds.; Springer: New York, NY, USA, 2013; pp. 61–86. ISBN 978-1-4614-6843-1. [Google Scholar]
- Casanova, M.F.; Sokhadze, E.; Opris, I.; Wang, Y.; Li, X. Autism spectrum disorders: Linking neuropathological findings to treatment with transcranial magnetic stimulation. Acta Pediatr. 2015, 104, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Sokhadze, E.M.; Casanova, M.F.; Casanova, E.; Kelly, D.P.; Khachidze, I.; Wang, Y.; Li, X. Applications of ERPs in autism research and as functional outcomes of neuromodulation treatment. In Event-Related Potential (ERP): Methods, Outcomes and Research Insights; Harris, S.R., Ed.; NOVA Science Publishers: Hauppauge, NY, USA, 2017; pp. 27–88. ISBN 1536108057. [Google Scholar]
- Duncan, C.C.; Barry, R.J.; Connolly, J.F.; Fischer, C.; Michie, P.T.; Näätänen, R.; Polich, J.; Reinvang, I.; van Petten, C. Event-related potentials in clinical research: Guidelines for eliciting, recording, and quantifying mismatch negativity, P300, and N400. Clin. Neurophysiol. 2009, 120, 1883–1908. [Google Scholar] [CrossRef] [PubMed]
- Picton, T.W. The P300 wave of the human event-related potential. J. Clin. Neurophysiol. 1992, 9, 456–479. [Google Scholar] [CrossRef] [PubMed]
- Polich, J. Updating P300: An integrative theory of P3a and P3b. Clin. Neurophysiol. 2007, 118, 2128–2148. [Google Scholar] [CrossRef] [PubMed]
- Pritchard, W. Psychophysiology of P300. Psychol. Bull. 1981, 89, 506–540. [Google Scholar] [CrossRef] [PubMed]
- Coles, M.G.H.; Rugg, M.D. Event-related brain potentials: An introduction. In Electrophysiology of Mind. Event-Related Brain Potentials and Cognition; Rugg, M.D., Coles, M.G.H., Eds.; Oxford University Press: Oxford, UK, 1995; pp. 1–26. ISBN 0198524161. [Google Scholar]
- Herrmann, C.S.; Knight, R.T. Mechanisms of human attention: Event related potentials and oscillations. Neurosci. Biobehav. Rev. 2001, 25, 465–476. [Google Scholar] [CrossRef]
- Patel, S.H.; Azzam, P.N. Characterization of N200 and P300: Selected studies of the event-related potential. Int. J. Med. Sci. 2005, 2, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, J.E. Event-related potentials and controlled processes. In Event-Related Brain Potentials: Basic Issues and Applications; Rohrbaugh, J.W., Parasuraman, R., Johnson, R., Jr., Eds.; Oxford University Press: New York, NY, USA, 1990; pp. 145–157. ISBN 0195048911. [Google Scholar]
- Katayama, J.; Polich, J. Stimulus context determines P3a and P3b. Psychophysiol. 1998, 35, 23–33. [Google Scholar] [CrossRef]
- Polich, J. Theoretical overview of P3a and P3b. In Detection of Change: Event-related Potential and fMRI Findings; Polich, J., Ed.; Kluwer Academic Press: Boston, MA, USA, 2003; pp. 83–98. ISBN 978-1-4613-5008-8. [Google Scholar]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-IV-TR), 4th ed.; Text Revised; American Psychiatric Publishing, Inc.: Washington, DC, USA, 2000; ISBN 0890420629. [Google Scholar]
- Le Couteur, A.; Lord, C.; Rutter, M. The Autism Diagnostic Interview—Revised (ADI-R); Western Psychological Services: Los Angeles, CA, USA, 2003. [Google Scholar]
- Wechsler, D. Wechsler Intelligence Scale for Children—Fourth Edition (WISC-IV); Harcourt Assessment, Inc.: San Antonio, TX, USA, 2003. [Google Scholar]
- Roid, G.H. Stanford-Binet Intelligence Scales Technical Manual, 5th ed.; Riverside Publishing: Itasca, IL, USA, 2003. [Google Scholar]
- Wechsler, D. Wechsler Abbreviated Scale for Intelligence; Psychological Corporation: San Antonio, TX, USA, 1999. [Google Scholar]
- First, M.B.; Spitzer, R.L.; Gibbon, M.; Williams, J.B.W. Structured Clinical Interview for DSM-IV-TR Axis I Disorders—Non-Patient Edition (SCID-NP); New York State Psychiatric Institute: New York, NY, USA, 2002. [Google Scholar]
- Net Station Acquisition Technical Manual; Electrical Geodesics, Inc.: Eugene, OR, USA, 2003.
- Fletcher, E.M.; Kussmaul, C.L.; Mangun, G.R. Estimation of interpolation errors in scalp topographic mapping. Electroctoencephalogr. Clin. Neurophysiol. 1996, 98, 422–434. [Google Scholar] [CrossRef]
- Luu, P.; Tucker, D.M.; Englander, R.; Lockfeld, A.; Lutsep, H.; Oken, B. Localizing acute stroke-related EEC changes: assessing the effects of spatial undersampling. J. Clin. Neurophysiol. 2001, 18, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Perrin, E.; Pernier, J.; Bertrand, O.; Giard, M.; Echallier, J.F. Mapping of scalp potentials by surface spline interpolation. Electroencephalogr. Clin. Neurophysiol. 1987, 66, 75–81. [Google Scholar] [CrossRef]
- Srinivasan, R.; Tucker, D.M.; Murias, M. Estimating the spatial Nyquist of the human EEG. Behav. Res. Methods 1998, 30, 8–19. [Google Scholar] [CrossRef]
- Franken, H.A.; van Strien, J.W.; Franzek, E.J.; van de Wetering, B.J. Error-processing deficits in patients with cocaine dependence. Biol. Psychol. 2007, 75, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Opris, I.; Lebedev, M.A.; Nelson, R.J. Neostriatal neuronal activity correlates better with movement kinematics under certain reward. Front. Neurosci. 2016, 10, 336. [Google Scholar] [CrossRef] [PubMed]
- Levine, T.R.; Hullett, C.R. Eta squared, partial eta squared, and misreporting of effect size in communication research. Hum. Commun. Res. 2002, 28, 612–625. [Google Scholar] [CrossRef]
- Cui, T.; Wang, P.P.; Liu, S.; Zhang, X. P300 amplitude and latency in autism spectrum disorder: A meta-analysis. Eur. Child Adolesc. Psychiatry 2017, 26, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Kemner, C.; Verbaten, M.N.; Cuperus, J.M.; Camfferman, G.; Van Engeland, H. Visual and somatosensory event-related brain potentials in autistic children and three different control groups. Electroencephalogr. Clin. Neurophysiol. 1994, 92, 225–237. [Google Scholar] [CrossRef]
- Kemner, C.; Verbaten, M.N.; Cuperus, J.M.; Camfferman, G.; Van Engeland, H. Auditory event- related potentials in autistic children and three different control groups. Biol. Psychiatry 1995, 38, 150–165. [Google Scholar] [CrossRef]
- Kemner, C.; van der Gaag, R.J.; Verbaten, M.; van Engeland, H. ERP differences among subtypes of pervasive developmental disorders. Biol. Psychiatry 1999, 46, 781–789. [Google Scholar] [CrossRef]
- Donchin, E.; Coles, M.G. Is the p300 component a manifestation of context updating? Behav. Brain Sci. 1988, 11, 357–374. [Google Scholar] [CrossRef]
- Townsend, J.; Westerfield, M.; Leaver, E.; Makeig, S.; Jung, T.; Pierce, K.; Courchesne, E. Event-related brain response abnormalities in autism: Evidence for impaired cerebello-frontal spatial attention networks. Brain Res. Cogn. Brain Res. 2001, 11, 127–145. [Google Scholar] [CrossRef]
- Ciesielski, K.T.; Courchesne, E.; Elmasian, R. Effects of focused attention tasks on event-related potentials in autistic and normal individuals. Electroencephalogr. Clin. Neurophysiol. 1990, 75, 207–220. [Google Scholar] [CrossRef]
- Ferri, R.; Elia, M.; Agarwal, N.; Lanuzza, B.; Musumeci, S.A.; Pennisi, G. The mismatch negativity and the P3a components of the auditory event-related potentials in autistic low-functioning subjects. Clin. Neurophysiol. 2003, 114, 1671–1680. [Google Scholar] [CrossRef]
- Bomba, M.D.; Pang, E.W. Cortical auditory evoked potentials in autism: A review. Int. J. Psychophysiol. 2004, 53, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Verbaten, M.N.; Roelofs, J.W.; van Engeland, H.; Kenemans, J.K.; Slangen, J.L. Abnormal visual event-related potentials of autistic children. J. Autism Dev. Disord. 1991, 21, 449–470. [Google Scholar] [CrossRef] [PubMed]
- Courchesne, E.; Lincoln, A.J.; Yeung-Courchesne, R.; Elmasian, R.; Grillon, C. Pathophysiologic findings in nonretarded autism and receptive developmental disorder. J. Autism Dev. Disord. 1989, 19, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, A.J.; Courchesne, E.; Harms, L.; Allen, M. Contextual probability evaluation in autistic, receptive developmental disorder and control children: Event-related potential evidence. J. Autism Dev. Disord. 1993, 23, 37–58. [Google Scholar] [CrossRef] [PubMed]
- Belmonte, M.K.; Yurgelun-Todd, D.A. Functional anatomy of impaired selective attention and compensatory processing in autism. Brain Res. Cogn. Brain Res. 2003, 17, 651–664. [Google Scholar] [CrossRef]
- Belmonte, M.K.; Yurgelun-Todd, D.A. Anatomic dissociation of selective and suppressive processes in visual attention. Neuroimage 2003, 19, 180–189. [Google Scholar] [CrossRef]
- Hopf, J.M.; Vogel, E.; Woodman, G.; Heinze, H.J.; Luck, S.J. Localizing visual discrimination processes in time and space. J. Neurophysiol. 2002, 88, 2088–2095. [Google Scholar] [CrossRef] [PubMed]
- Hillyard, S.A.; Anllo-Vento, L. Event-related brain potentials in the study of visual selective attention. Proc. Natl. Acad. Sci. USA 1998, 95, 781–787. [Google Scholar] [CrossRef] [PubMed]
- Luck, S.J.; Heinze, H.; Mangun, G.R.; Hillyard, S.A. Visual event-related potentials index focused attention within bilateral stimulus arrays. II. Functional dissociation of P1 and N1 components. Electroencephalogr. Clin. Neurophysiol. 1990, 75, 528–542. [Google Scholar] [CrossRef]
- Boutros, N.N.; Korzyukov, O.; Jansen, B.; Feingold, A.; Bell, M. Sensory gating deficits during the mid-latency phase of information processing in medicated schizophrenia patients. Psychiatry Res. 2004, 126, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.F.; Buxhoeveden, D.; Gomez, J. Disruption in the inhibitory architecture of the cell minicolumn: Implications for autism. Neuroscientist 2003, 9, 496–507. [Google Scholar] [CrossRef] [PubMed]
- Rubenstein, J.L.; Merzenich, M.M. Model of autism: Increased ratio of excitation/inhibition in key neural systems. Genes Brain Behav. 2003, 2, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Overbeek, T.J.M.; Nieuwenhuis, S.; Ridderinkhof, K.R. Dissociable components of error processing. J. Psychophysiol. 2005, 19, 319–329. [Google Scholar] [CrossRef]
- Yeung, N.; Botvinick, M.M.; Cohen, J.D. The neural basis of error detection: Conflict monitoring and the error-related negativity. Psychol. Rev. 2004, 111, 931–959. [Google Scholar] [CrossRef] [PubMed]
- Yeung, N.; Cohen, J.D. The impact of cognitive deficits on conflict monitoring. Predictable dissociations between the error-related negativity and N2. Psychol. Sci. 2006, 17, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Van Veen, V.; Carter, C.S. The timing of action-monitoring process in the anterior cingulate cortex. J. Cogn. Neurosci. 2002, 14, 593–602. [Google Scholar] [CrossRef] [PubMed]
- Burle, B.; Vidal, F.; Tandonnet, C.; Hasbroucq, T. Physiological evidence for response inhibition in choice reaction time tasks. Brain Cogn. 2004, 56, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Ridderinkhof, K.R.; Ullsperger, M.; Crone, E.A.; Nieuwenhuis, S. The role of the medial frontal cortex in cognitive control. Science 2004, 306, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Casanova, M.F. Minicolumnar pathology in autism. In Recent Developments in Autism Research; Casanova, M.F., Ed.; Nova Biomedical Books: New York, NY, USA, 2005; pp. 133–143. ISBN 1-59454-497-2. [Google Scholar]
- Casanova, M.F. Neuropathological and genetic findings in autism: The significance of a putative minicolumnopathy. Neuroscientist 2006, 12, 435–441. [Google Scholar] [CrossRef] [PubMed]
- Just, M.A.; Cherkassky, V.; Keller, T.A.; Minshew, N.J. Cortical activation and synchronization during sequence comprehension in high-functioning autism: Evidence of underconnectivity. Brain 2004, 127, 1811–1821. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.L.; Casanova, M.F. Autism and dyslexia: A spectrum of cognitive styles as defined by minicolumnar morphometry. Med. Hypotheses 2009, 74, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Bogte, H.; Flamma, B.; van der Meere, J.; van Engeland, H. Post-error adaptation in adults with high functioning autism. Neuropsychologia 2007, 45, 1707–1714. [Google Scholar] [CrossRef] [PubMed]
- Bauman, M.L.; Kemper, T.L. Structural brain anatomy in autism: What is the evidence? In The Neurobiology of Autism, 2nd ed.; Bauman, M.L., Kemper, T.L., Eds.; John Hopkins University Press: Baltimore, MD, USA, 2005; pp. 121–135. ISBN 0801880475. [Google Scholar]
- Minshew, N.J.; Sweeney, J.A.; Bauman, M.L.; Webb, S.J. Neurological aspects of autism. In Handbook of Autism and Pervasive Developmental Disorders, Diagnosis, Development, Neurobiology, and Behavior, 3rd ed.; Volkmar, F.R., Paul, R., Klin, A., Cohen, D., Eds.; Wiley: New York, NY, USA, 2005; Volume 1, pp. 473–514. ISBN 0471716960. [Google Scholar]
- Mundy, P. Annotation: The neural basis of social impairments in autism: The role of the dorsal medial-frontal cortex and anterior cingulate system. J. Child Psychol. Psychiatry 2003, 44, 793–809. [Google Scholar] [CrossRef] [PubMed]
- Henderson, H.; Schwartz, C.; Mundy, P.; Burnette, C.; Sutton, S.; Zahka, N.; Pradella, A. Response monitoring, the error-related negativity, and differences in social behavior in autism. Brain Cogn. 2006, 61, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Devinsky, O.; Luciano, D. The contributions of cingulate cortex to human behavior. I. In Neurobiology of Cingulate Cortex and Limbic Thalamus: A Comprehensive Handbook; Gabriel, M., Vogt, B.A., Eds.; Birkhauser: Boston, MA, USA, 1993; pp. 527–556. ISBN 978-1-4899-6706-0. [Google Scholar]
- Tamnes, C.K.; Walhovd, K.B.; Torstveit, M.; Sells, V.T.; Fjell, A.M. Performance monitoring in children and adolescents: A review of developmental changes in the error-related negativity and brain maturation. Dev. Cogn. Neurosci. 2013, 6, 1–13. [Google Scholar] [CrossRef] [PubMed]






© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sokhadze, E.M.; Lamina, E.V.; Casanova, E.L.; Kelly, D.P.; Opris, I.; Khachidze, I.; Casanova, M.F. Atypical Processing of Novel Distracters in a Visual Oddball Task in Autism Spectrum Disorder. Behav. Sci. 2017, 7, 79. https://doi.org/10.3390/bs7040079
Sokhadze EM, Lamina EV, Casanova EL, Kelly DP, Opris I, Khachidze I, Casanova MF. Atypical Processing of Novel Distracters in a Visual Oddball Task in Autism Spectrum Disorder. Behavioral Sciences. 2017; 7(4):79. https://doi.org/10.3390/bs7040079
Chicago/Turabian StyleSokhadze, Estate M., Eva V. Lamina, Emily L. Casanova, Desmond P. Kelly, Ioan Opris, Irma Khachidze, and Manuel F. Casanova. 2017. "Atypical Processing of Novel Distracters in a Visual Oddball Task in Autism Spectrum Disorder" Behavioral Sciences 7, no. 4: 79. https://doi.org/10.3390/bs7040079
APA StyleSokhadze, E. M., Lamina, E. V., Casanova, E. L., Kelly, D. P., Opris, I., Khachidze, I., & Casanova, M. F. (2017). Atypical Processing of Novel Distracters in a Visual Oddball Task in Autism Spectrum Disorder. Behavioral Sciences, 7(4), 79. https://doi.org/10.3390/bs7040079