
Learned Use of Picture Cues by Bumblebees (Bombus impatiens) in a Delayed Matching Task

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
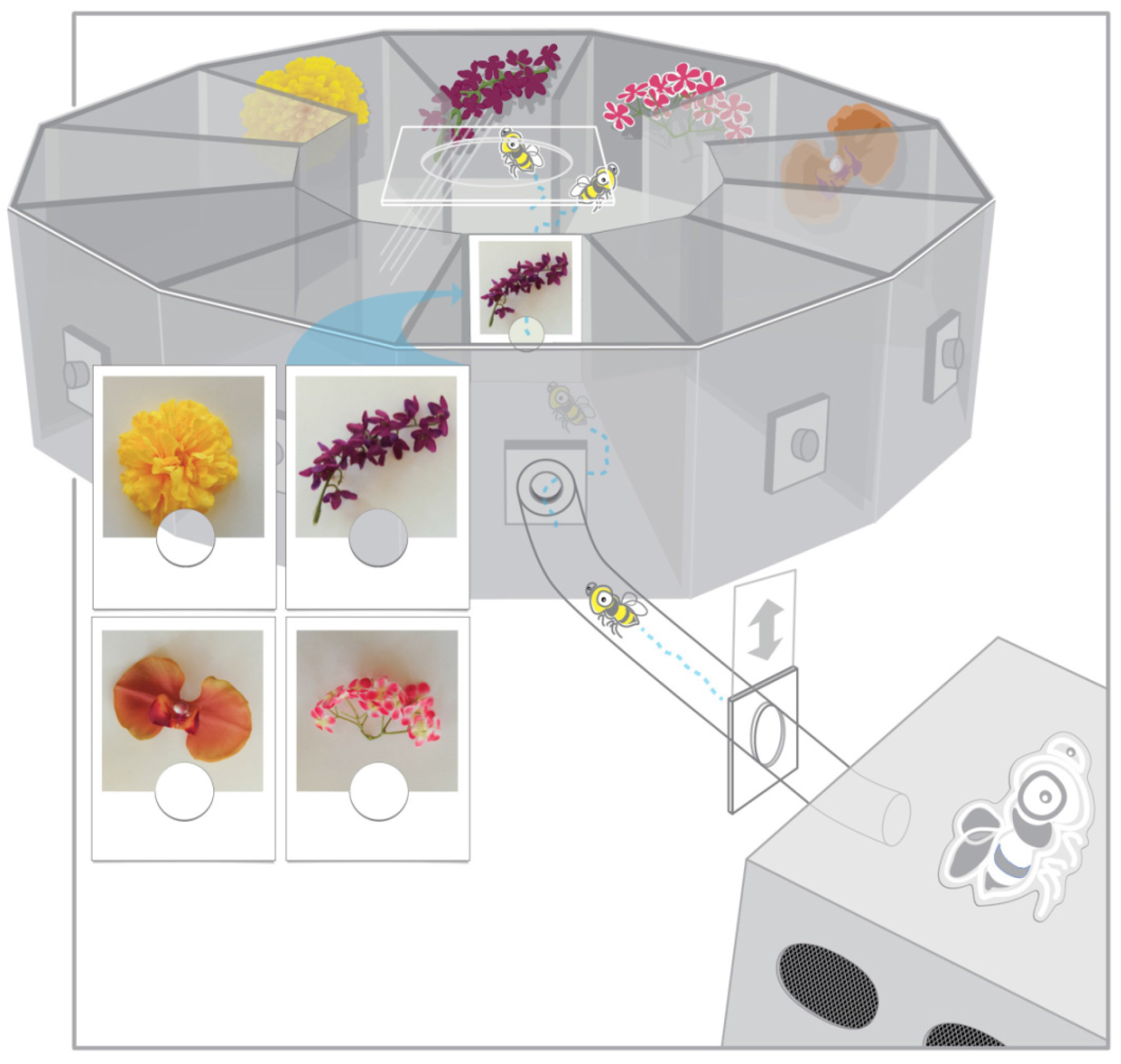
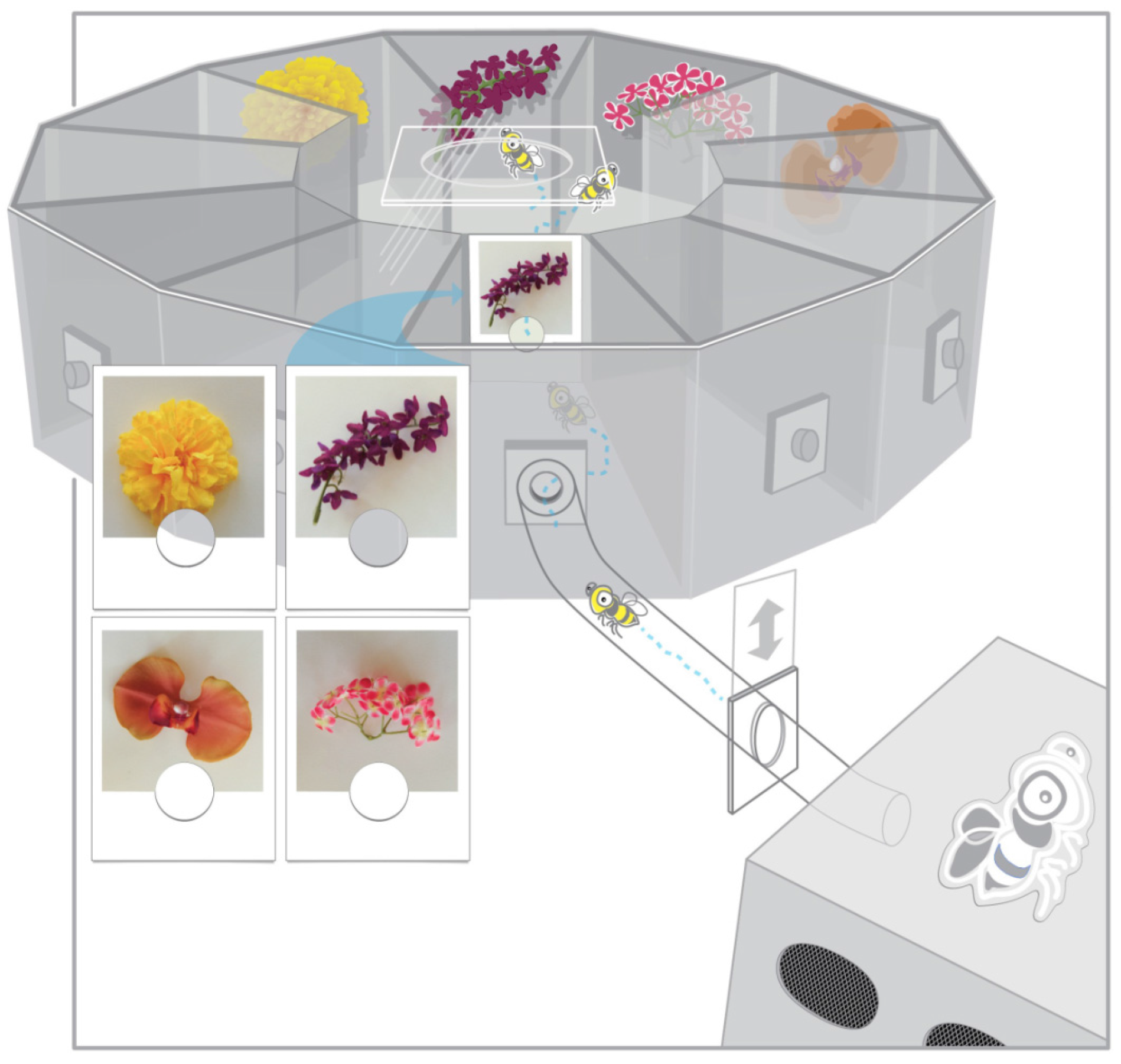
2. Materials and Methods
2.1. Subjects
2.2. Materials
2.3. Procedure
2.3.1. Design
2.3.2. Training
2.3.3. Testing
2.4. Statistical Analysis
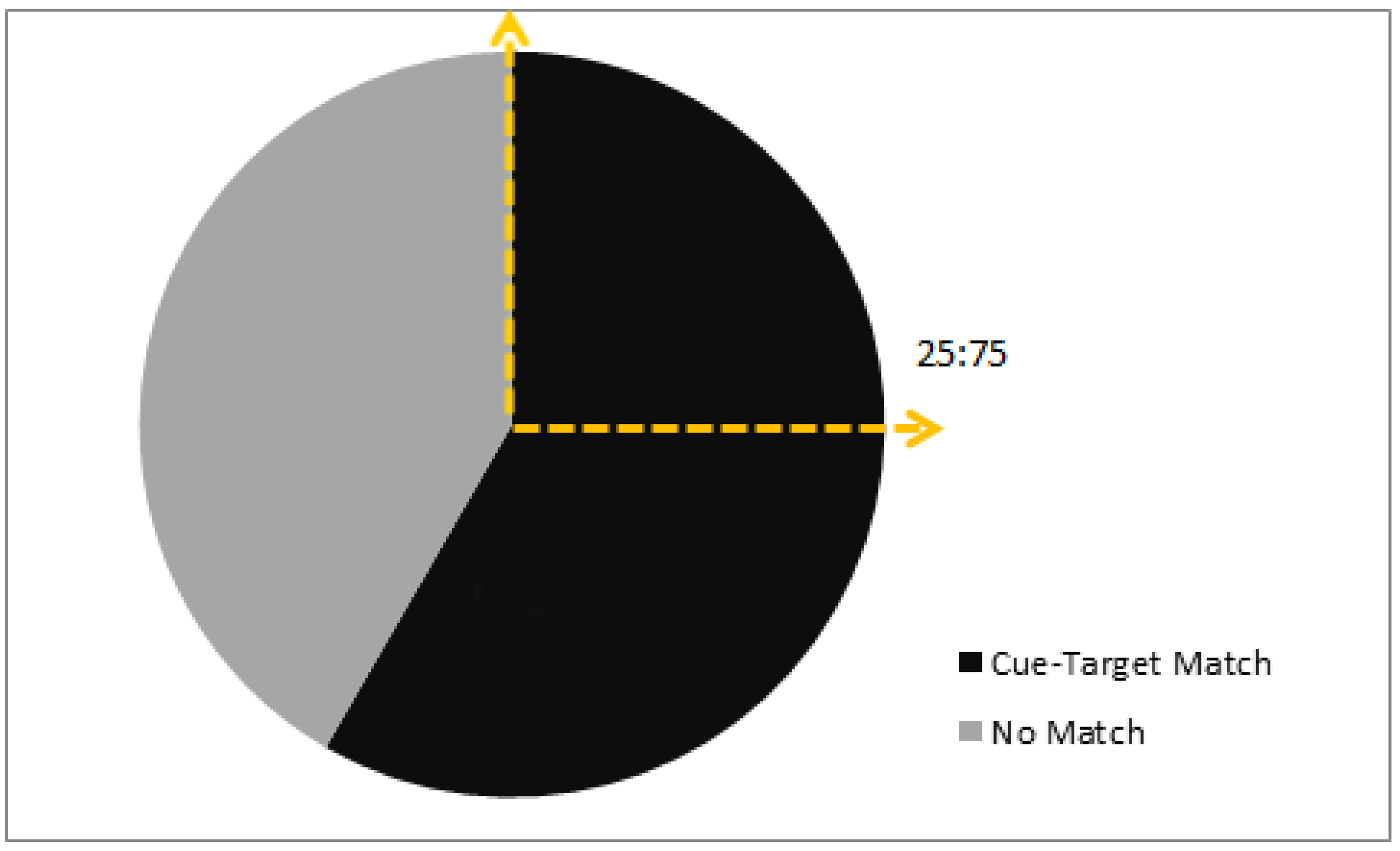
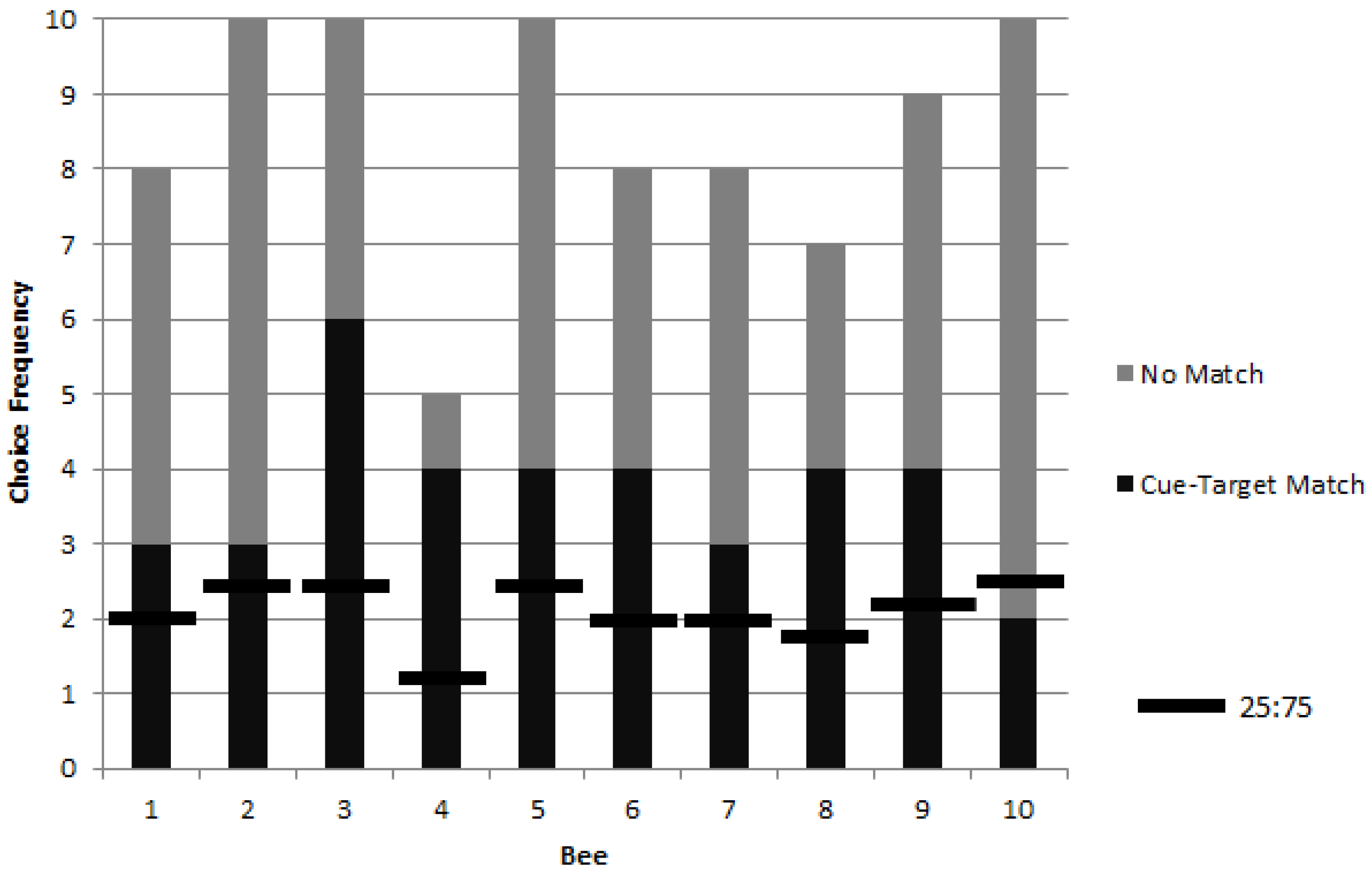
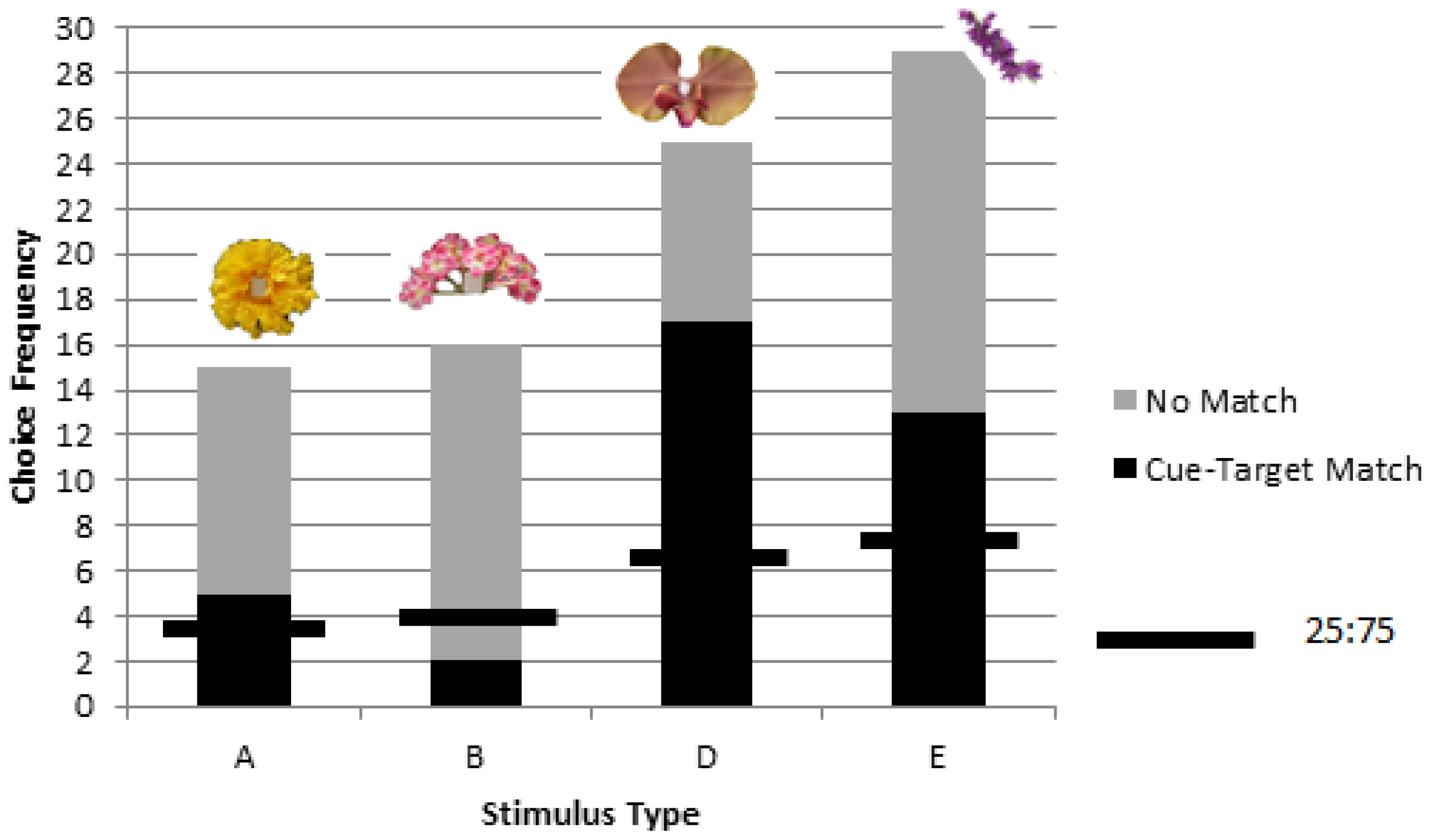
3. Results
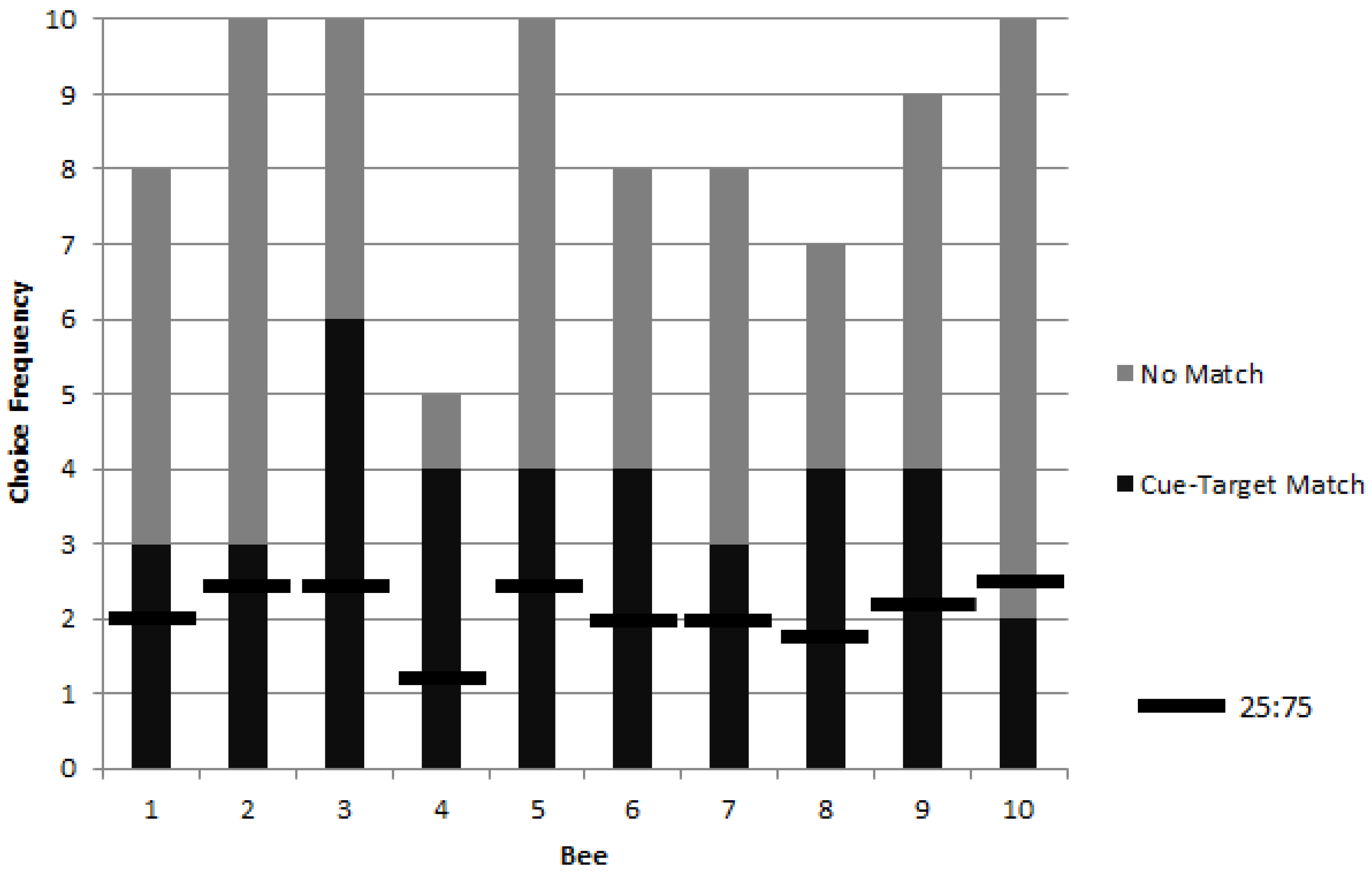
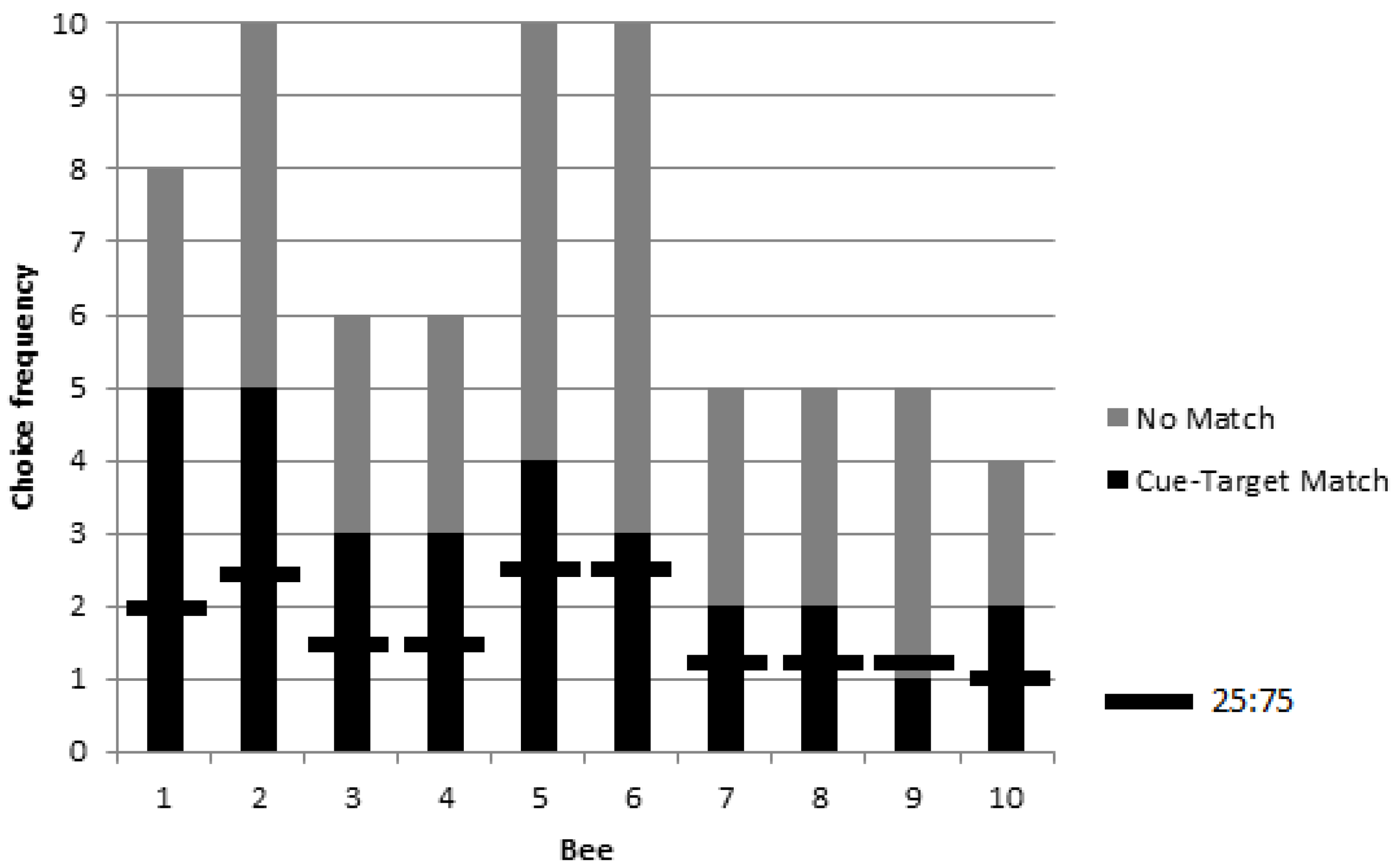
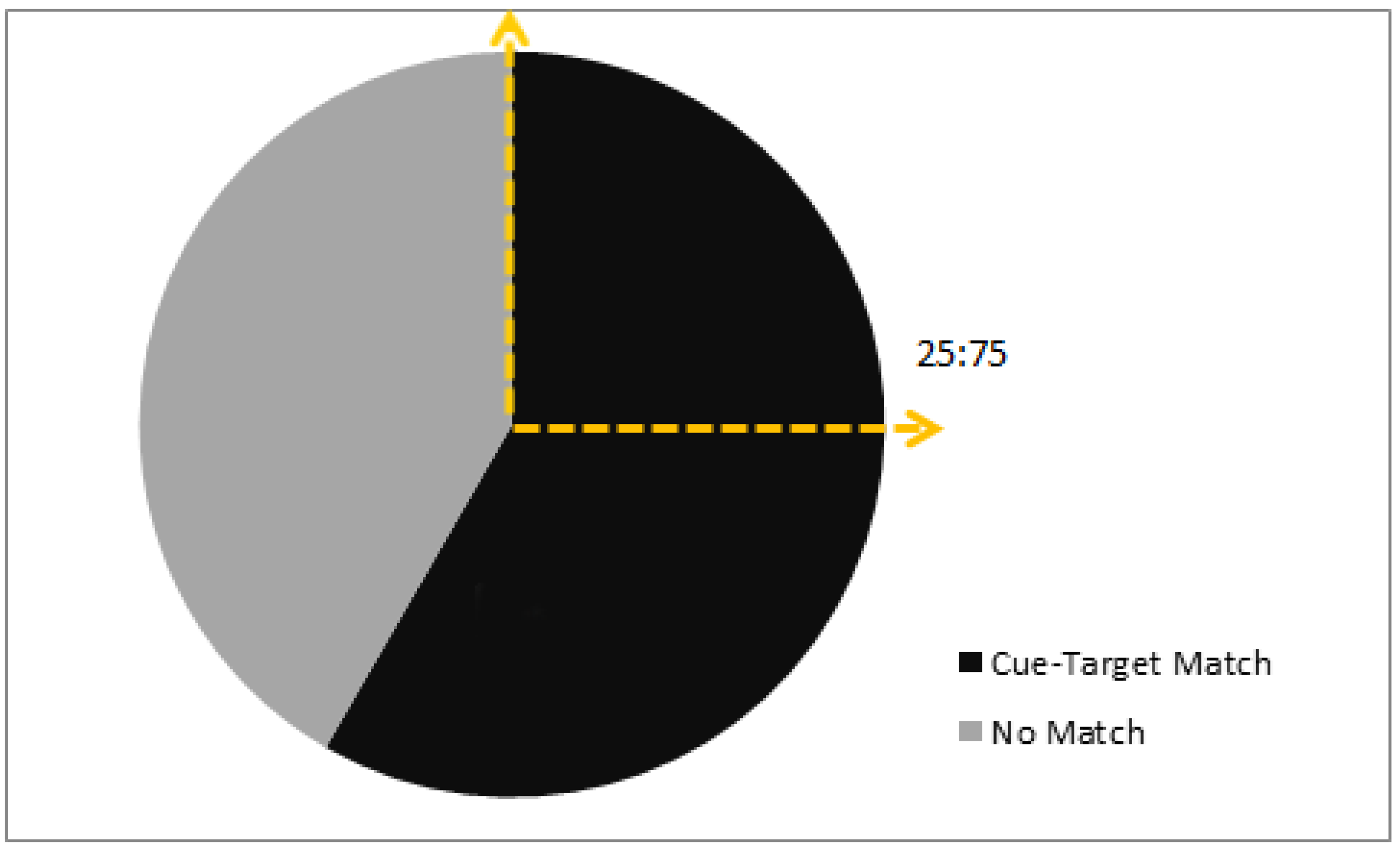
3.1. Colony 1: Rewarded Tests of Delayed Matching
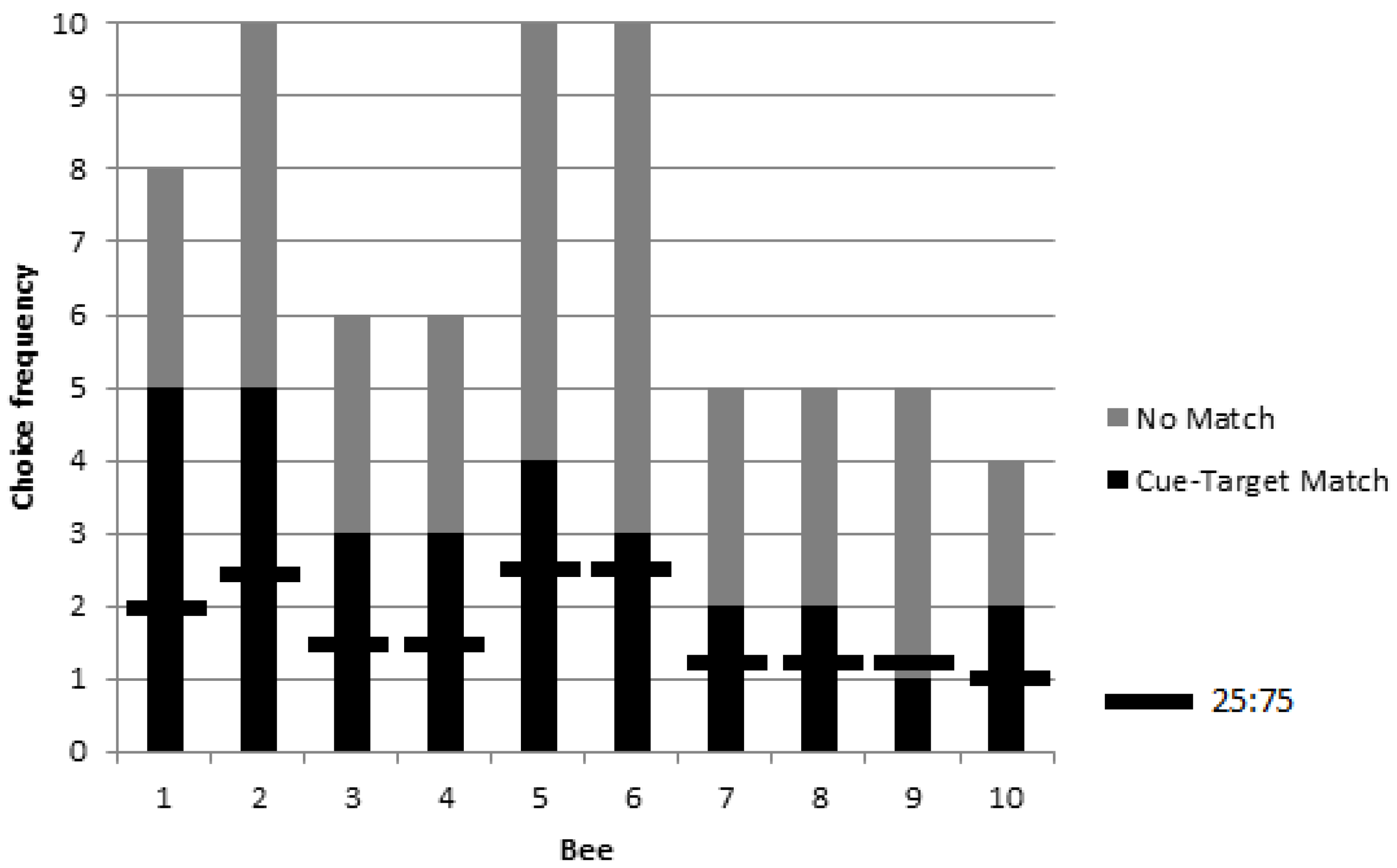
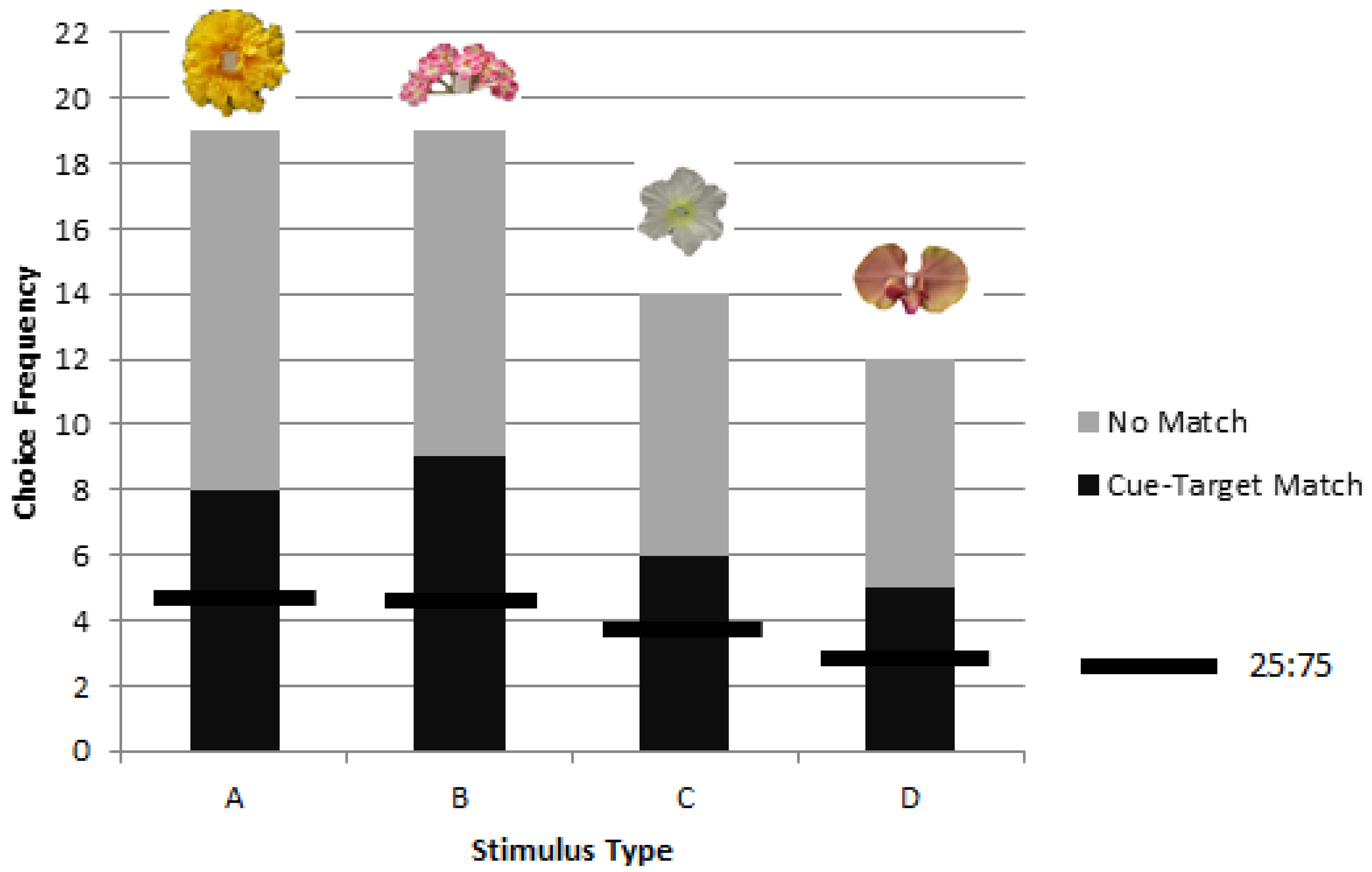
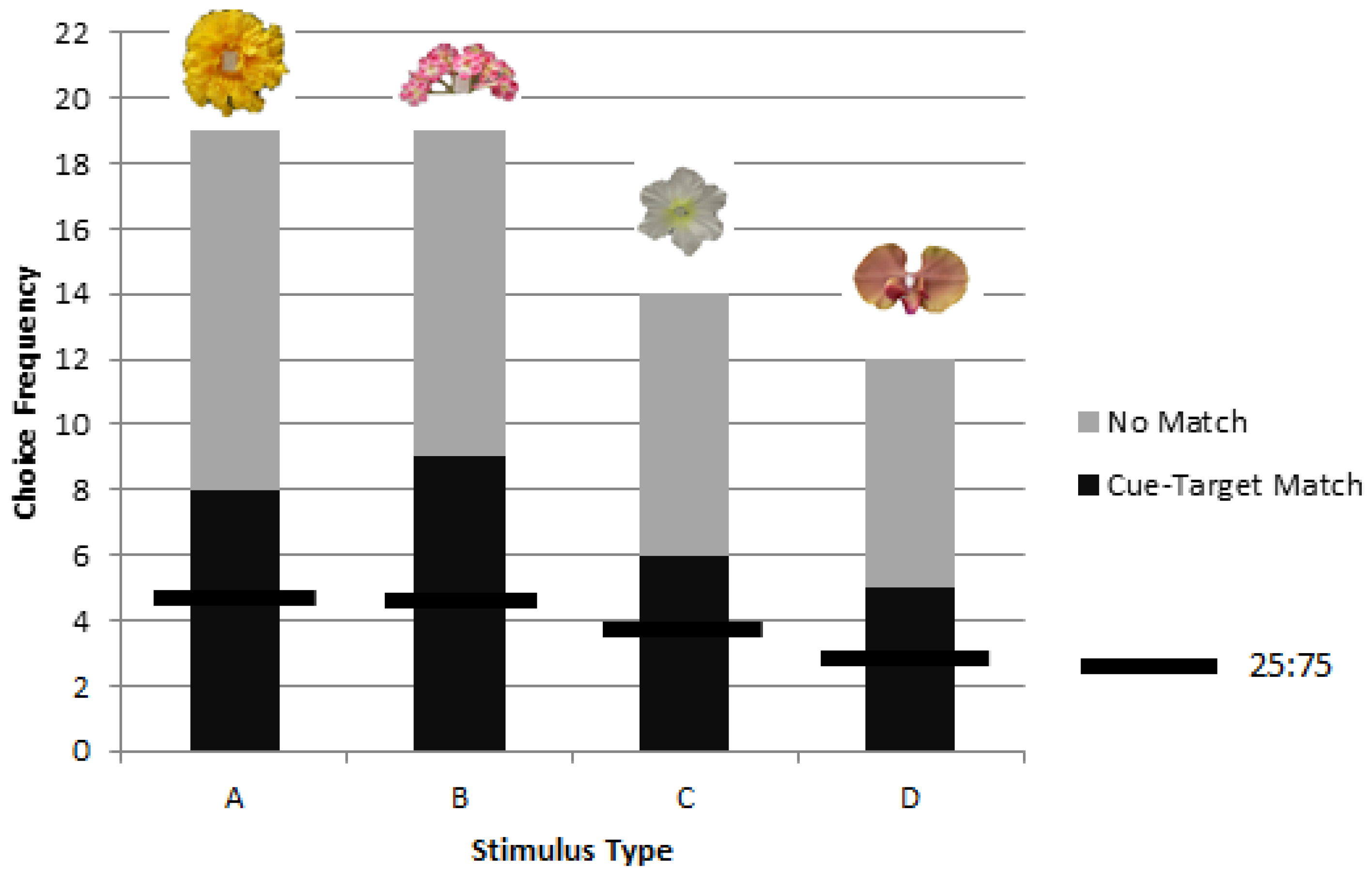
3.2. Colonies 2 and 3: Rewarded, Unrewarded, and Novel Tests of Delayed Matching
4. Discussion
(i) Experience
(ii) Reward Inconsistency
(iii) Location
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dukas, R. Constraints on information processing and their effects on behavior. In Cognitive Ecology: The Evolutionary Ecology of Information Processing and Decision Making; Dukas, R., Ed.; The University of Chicago Press: Chicago, IL, USA; London, UK, 1998; pp. 89–119. [Google Scholar]
- Dukas, R. Learning: Mechanisms, ecology, and evolution. In Cognitive Ecology II; Dukas, R., Ratcliffe, J.M., Eds.; The University of Chicago Press: Chicago, IL, USA; London, UK, 2009; pp. 7–26. [Google Scholar]
- Kendal, R.L.; Coolen, I.; Laland, K.N. Adaptive trade-offs in the use of social and personal information. In Cognitive Ecology II; Dukas, R., Ratcliffe, J.M., Eds.; The University of Chicago Press: Chicago, IL, USA; London, UK, 2009; pp. 249–271. [Google Scholar]
- Chittka, L.; Geiger, K. Can honey bees count landmarks? Anim. Behav. 1995, 49, 159–164. [Google Scholar] [CrossRef]
- Shettleworth, S.J. Cognition, Evolution and Behavior, 2nd ed.; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- Brown, M.F.; Sayde, J.M. Same/different discrimination by bumblebee colonies. Anim. Cognit. 2013, 16, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Dale, K.; Harland, D.P.; Manning-Jones, A.; Collett, T.S. Weak and strong priming cues in bumblebee contextual learning. J. Exp. Biol. 2005, 208, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Cooke, M.H.Y.; Couvillon, P.A.; Bitterman, M.E. Delayed symbolic matching in honeybees (Apis mellifera). J. Comp. Psychol. 2007, 121, 106–108. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W.; Lehrer, M.; Srinivasan, M.V. Honeybee memory: Navigation by associative grouping and recall of visual stimuli. Neurobiol. Learn. Mem. 1999, 72, 180–201. [Google Scholar] [CrossRef] [PubMed]
- Giurfa, M.; Zhang, S.; Jenett, A.; Menzel, R.; Srinivasan, M.V. The concepts of ‘sameness’ and ‘difference’ in an insect. Nature 2001, 410, 930–933. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Bock, F.; Si, A.; Tautz, J.; Srinivasan, M.V. Visual working memory in decision making by honeybees. Proc. Natl. Acad. Sci. USA 2005, 102, 5250–5255. [Google Scholar] [CrossRef] [PubMed]
- Sherry, D.F.; Strang, C.G. Contrasting styles in cognition and behaviour in bumblebees and honeybees. Behav. Process. 2015, 117, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Dyer, A.G.; Spaethe, J.; Prack, S. Comparative psychophysics of bumblebee and honeybee colour discrimination and object detection. J. Comp. Physiol. A 2008, 194, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Avarguès-Weber, A.; Deisig, N.; Giurfa, M. Visual cognition in social insects. Ann. Rev. Entomol. 2011, 56, 423–443. [Google Scholar] [CrossRef] [PubMed]
- Fauria, K.; Dale, K.; Colborn, M.; Collett, T.S. Learning speed and contextual isolation in bumblebees. J. Exp. Biol. 2002, 205, 1009–1018. [Google Scholar] [PubMed]
- Van Hest, A.; Steckler, T. Effects of procedural parameters on response accuracy: Lessons from delayed (non-) matching procedures in animals. Cogn. Brain Res. 1996, 3, 193–203. [Google Scholar] [CrossRef]
- Thompson, E.L.; Plowright, C.M.S. How images may or may not represent flowers: Picture-object correspondence in bumblebees (Bombus impatiens)? Anim. Cognit. 2014, 17, 1031–1043. [Google Scholar] [CrossRef] [PubMed]
- Chittka, L.; Jensen, K. Animal cognition: Concepts from apes to bees. Curr. Biol. 2011, 21, R116–R119. [Google Scholar] [CrossRef] [PubMed]
- Plowright, C.M.S.; Ferguson, K.A.; Jellen, S.L.; Xu, V.; Service, E.W.; Dookie, A.L. Bumblebees occupy: When foragers do and do not use the presence of others to first find food. Insectes Sociaux 2013, 60, 517–524. [Google Scholar] [CrossRef]
- Srinivasan, M.V.; Lehrer, M. Temporal acuity of honeybee vision: Behavioural studies using moving stimuli. J. Comp. Physiol. A 1984, 155, 297–312. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry, 4th ed.; W.H. Freeman: New York, NY, USA, 2012. [Google Scholar]
- Keshen, C.; Cohen, J. Rats (Rattus norvegicus) flexibly retrieve objects’ non-spatial and spatial information from their visuospatial working memory: Effects of integrated and separate processing of these features in a missing-object recognition task. Anim. Cognit. 2016, 19, 91–107. [Google Scholar] [CrossRef] [PubMed]
- Lind, J.; Enquist, M.; Ghirlanda, S. Animal memory: A review of delayed matching-to-sample data. Behav. Process. 2014, 117, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Kulahci, I.G.; Dornhaus, A.; Papaj, D.R. Multimodal signals enhance decision making in foraging bumble-bees. Proc. R. Soc. Biol. Sci. 2008, 275, 797–802. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.S.; Dornhaus, A.; Papaj, D.R. Forget-me-not: Complex floral displays, inter-signal interactions, and pollinator cognition. Curr. Zool. 2011, 57, 215–224. [Google Scholar] [CrossRef]
- Raguso, R.A. Flowers as sensory billboards: Progress towards an integrated understanding of floral advertisement. Curr. Opin. Plant Biol. 2004, 7, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Dornhaus, A.; Chittka, L. Evolutionary origins of bee dances. Nature 1999, 401, 38. [Google Scholar] [CrossRef]
- Raine, N.E.; Chittka, L. No trade-off between learning speed and associative flexibility in bumblebees: A reversal learning test with multiple colonies. PLoS ONE 2012, 7, e45096. [Google Scholar] [CrossRef] [PubMed]
- Jin, N.; Landgraf, T.; Klein, S.; Menzel, R. Walking bumblebees memorize panorama and local cues in a laboratory test of navigation. Anim. Behav. 2014, 97, 13–23. [Google Scholar] [CrossRef]
- Perreault, M.; Plowright, C.M.S. Facilitation of learning of a simultaneous discrimination between rotated patterns by bumblebees. Learn. Motiv. 2009, 40, 234–256. [Google Scholar] [CrossRef]
- Kamil, A.C.; Bond, A.B. Selective attention, priming and foraging behavior. In Comparative Cognition: Experimental Explorations of Animal Intelligence; Wasserman, E.A., Zentall, T.R., Eds.; Oxford University Press: Oxford, UK, 2006; pp. 106–126. [Google Scholar]
- Reinhard, J.; Srinivasan, M.V.; Guez, D.; Zhang, S.W. Floral scents induce recall of navigational and visual memories in honeybees. J. Exp. Biol. 2004, 207, 4371–4381. [Google Scholar] [CrossRef] [PubMed]
- McLinn, C.M.; Stephens, D.W. What makes information valuable: Signal reliability and environmental uncertainty. Anim. Behav. 2006, 71, 1119–1129. [Google Scholar] [CrossRef]
- Defrize, J.; Llandres, A.L.; Casas, J. Indirect cues in selecting a hunting site in a sit-and-wait predator. Physiol. Entomol. 2014, 39, 53–59. [Google Scholar] [CrossRef]
- Chittka, L.; Thomson, J.D.; Waser, N.M. Flower constancy, insect psychology, and plant evolution. Naturwissenchaften 1999, 86, 361–377. [Google Scholar] [CrossRef]
- Collett, T.S. Invertebrate memory: Honeybees with a sense of déjà vu. Curr. Biol. 2005, 15, R419–R421. [Google Scholar] [CrossRef] [PubMed]
- Menzel, R. Searching for the memory trace in a mini-brain, the honeybee. Learn. Mem. 2001, 8, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Lotto, R.B.; Chittka, L. Seeing the light: Illumination as a contextual cue to color choice behavior in bumblebees. Proc. Natl. Acad. Sci. USA 2005, 102, 3852–3856. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thompson, E.; Plowright, C. Learned Use of Picture Cues by Bumblebees (Bombus impatiens) in a Delayed Matching Task. Behav. Sci. 2016, 6, 22. https://doi.org/10.3390/bs6040022
Thompson E, Plowright C. Learned Use of Picture Cues by Bumblebees (Bombus impatiens) in a Delayed Matching Task. Behavioral Sciences. 2016; 6(4):22. https://doi.org/10.3390/bs6040022
Chicago/Turabian StyleThompson, Emma, and Catherine Plowright. 2016. "Learned Use of Picture Cues by Bumblebees (Bombus impatiens) in a Delayed Matching Task" Behavioral Sciences 6, no. 4: 22. https://doi.org/10.3390/bs6040022
APA StyleThompson, E., & Plowright, C. (2016). Learned Use of Picture Cues by Bumblebees (Bombus impatiens) in a Delayed Matching Task. Behavioral Sciences, 6(4), 22. https://doi.org/10.3390/bs6040022