Adrenal Steroids Uniquely Influence Sexual Motivation Behavior in Male Rats

{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
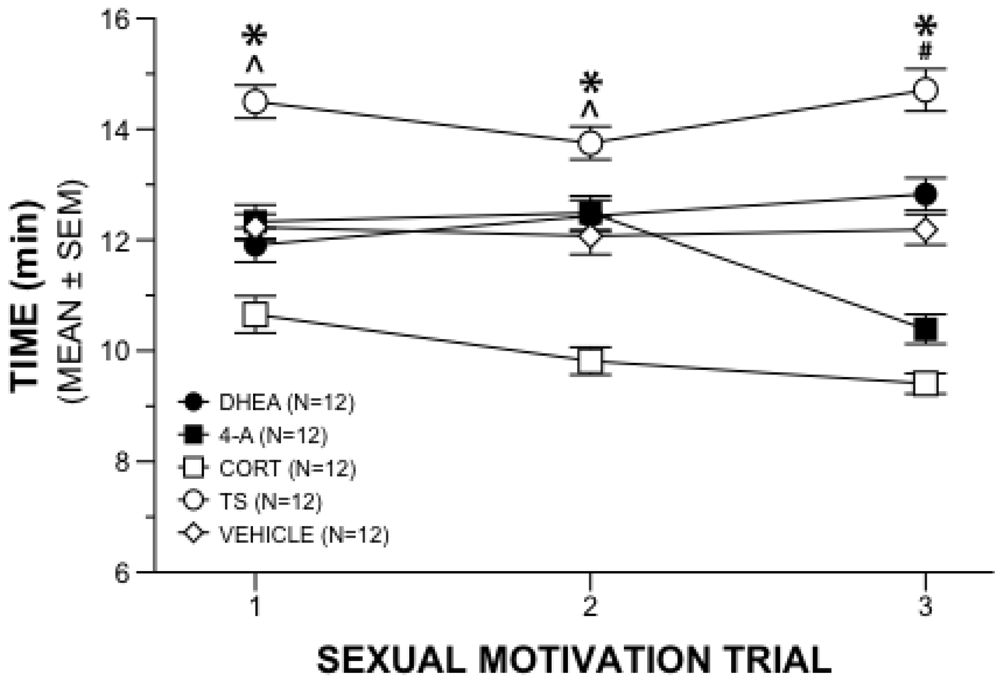
2.1. Sexual Motivation
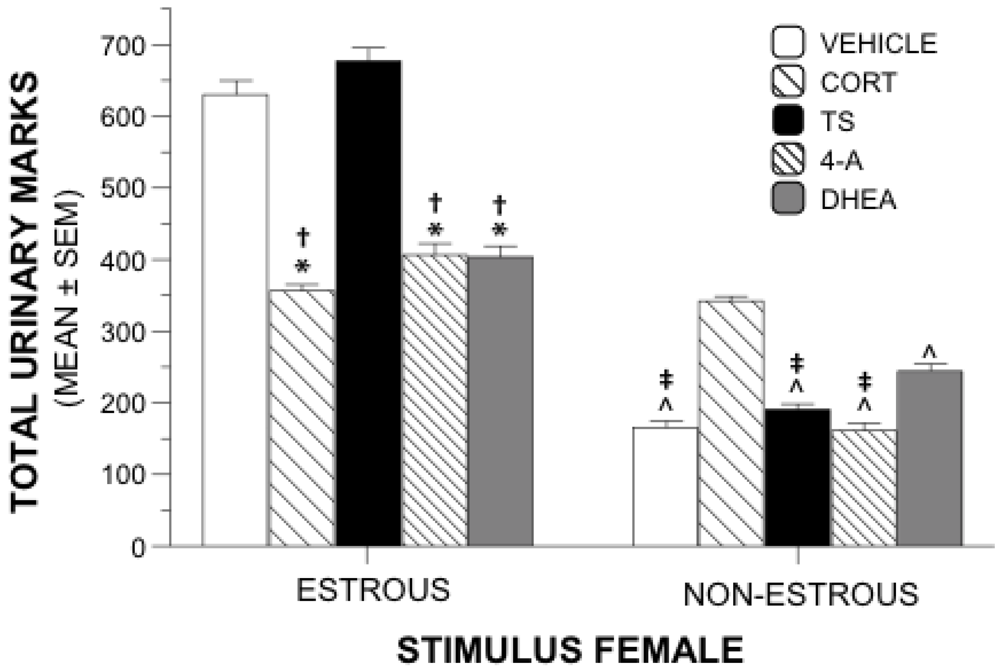
2.2. Urinary Marking and Open Field Activity
2.3. Discussion
3. Experimental Section
3.1. Subjects
3.2. Materials
3.2.1. Apparatus
3.2.2. Hormones
3.3. Experimental Design
3.4. Procedures
3.4.1. Surgery
3.4.2. Hormone Administration
3.4.3. Sexual Experience
3.4.4. Sexual Motivation and Urinary Marking
3.4.5. Open Field
3.4.6. Statistical Analyses
4. Conclusions
References
- Kanayama, G.; Hudson, J.I.; Pope, H.G., Jr. Illicit anabolic-androgenic steroid use. Horm. Behav. 2010, 58, 111–121. [Google Scholar] [CrossRef]
- Sakuma, Y. Neural substrates for sexual preference and motivation in the female and male rat. Ann. N. Y. Acad. Sci. 2008, 1129, 55–60. [Google Scholar]
- Rivier, C.; Rivest, S. Effect of stress on the activity of the hypothalamic-pituitary-gonadal axis: Peripheral and central mechanisms. Biol. Reprod. 1991, 45, 523–532. [Google Scholar] [CrossRef]
- McIntosh, M.; Bao, H.; Lee, C. Opposing actions of dehydroepiandrosterone and corticosterone in rats. Proc. Soc. Exp. Biol. Med. 1999, 221, 198–206. [Google Scholar]
- Taylor, G.T.; Womack, S.; Weiss, J.; Pitha, J. Behavior and physiological effects of supplemental episodes of testosterone, its precursors and metabolite in rats. Life Sci. 1990, 47, 1965–1971. [Google Scholar]
- Taylor, G.T.; Scherrer, J.; Weiss, J.; Pitha, J. Endocrine interactions: Adrenal steroids and precursors. Am. J. Physiol. 1994, 266, E676–E681. [Google Scholar]
- Taylor, G.T.; Weiss, J.; Haller, J. Chronic changes in male rats’ hormone-sensitive systems after suprathreshold pulses of testosterone. Anat. Rec. 1985, 211, 304–310. [Google Scholar] [CrossRef]
- Taylor, G.T.; Weiss, J.; Rupich, R. Suprathreshold manipulations of testosterone and reproductive functioning in gonadally intact sexually experienced and inexperienced male rats. Physiol. Behav. 1985, 35, 735–739. [Google Scholar]
- Alexander, G.M.; Swerdloff, R.S.; Wang, C.; Davidson, T.; McDonald, V.; Steiner, B.; Hines, M. Androgen-Behavior correlations in hypogonadal men and eugonadal men. I. Mood and response to auditory sexual stimuli. Horm. Behav. 1997, 31, 110–119. [Google Scholar] [CrossRef]
- Retana-Marquez, S.; Bonilla-Jaime, H.; Vazquez-Palacios, G.; Martinez-Garcia, R.; Velzquez-Moctezuma, J. Changes in masculine sexual behavior, corticosterone and testosterone in response to acute and chronic stress in male rats. Horm. Behav. 2003, 44, 327–337. [Google Scholar] [CrossRef]
- Gronli, J.; Murison, R.; Fiske, E.; Bjorvatn, B.; Sorensen, E.; Portas, C.M.; Ursin, R. Effects of chronic mild stress on sexual behavior, locomotor activity and consumption of sucrose and saccharine solutions. Physiol. Behav. 2005, 84, 571–577. [Google Scholar] [CrossRef]
- Roselli, C.E.; Cross, E.; Poonyagariyagorn, H.K.; Stadelman, H.L. Role of aromatization in anticipatory and consummatory aspects of sexual behavior in male rats. Horm. Behav. 2003, 44, 146–151. [Google Scholar] [CrossRef]
- Breuer, M.E.; McGinnis, M.Y.; Lumia, A.R.; Possidente, B.P. Aggression in male rats receiving anabolic androgenic steroids: Effects of social and environmental provocation. Horm. Behav. 2001, 40, 409–418. [Google Scholar] [CrossRef]
- Taylor, G.T.; Maloney, S.; Dearborn, J.; Weiss, J. Hormones in the mentally disturbed brain: Steroids and peptides in the development and treatment of psychopathology. Cent. Nerv. Syst. Agents Med. Chem. 2009, 9, 331–360. [Google Scholar]
- Brown, R.E. Hormonal control of odor preferences and urine-marking in male and female rats. Physiol. Behav. 1978, 20, 21–24. [Google Scholar] [CrossRef]
- Arteaga-Silva, M.; Marquez-Villanueva, Y.; Martinez-Garcia, R.; Hernandez-Gonzalez, M.; Bonilla-Jaime, H.; Retana-Marquez, S. Effects of hormonal replacement with androgens and estrogens on male sexual behavior and plasma levels of these steroids in gonadectomized golden hamsters (Mesocricetus auratus). Physiol. Behav. 2005, 85, 571–580. [Google Scholar] [CrossRef]
- Arteaga-Silva, M.; Vigueras-Villasenor, R.M.; Retana-Marquez, S.; Hernandez-Gonzalez, M.; Chihuahua-Serrano, C.; Bonilla-Jaime, H.; Contreras, J.L.; Morali, G. Testosterone, androstenedione, and 5alpha-dihydrotestosterone on male sexual behavior and penile spines in the hamster. Physiol. Behav. 2008, 94, 412–421. [Google Scholar] [CrossRef]
- Whalen, R.E.; Luttge, W.G. Testosterone, androstenedione, and dihydrotestosterone: Effects on mating behavior of male rats. Horm. Behav. 1971, 2, 117–125. [Google Scholar] [CrossRef]
- Beyer, C.; Larsson, K.; Perez-Palacios, G.; Morali, G. Androgen structure and male sexual behavior in the castrated rat. Horm. Behav. 1973, 4, 99–108. [Google Scholar] [CrossRef]
- Retana-Marquez, S.; Bonilla-Jaime, H.; Velazquez-Moctezuma, J. Lack of effect of corticosterone administration on male sexual behavior of rats. Physiol. Behav. 1998, 63, 367–370. [Google Scholar] [CrossRef]
- Taylor, G.T.; Komitowski, D.; Weiss, J. Light and scanning electron microscopic study of testosterone-restored penile papillae in castrated rats. Anat. Rec. 1983, 205, 277–286. [Google Scholar] [CrossRef]
- Maninger, N.; Wolkowitz, O.M.; Reus, V.I.; Epel, E.S.; Mellon, S.H. Neurobiological and neuropsychiatric effects of dehydroepiandrosterone (DHEA) and DHEA sulfate (DHEAS). Front. Neuroendocrinol. 2009, 30, 65–91. [Google Scholar] [CrossRef]
- Kaasik, A.; Safiulina, D.; Kalda, A.; Zharkovsky, A. Dehydroepiandrosterone with other neurosteroids preserve neuronal mitochondria from calcium overload. J. Steroid Biochem. Mol. Biol. 2003, 87, 97–103. [Google Scholar] [CrossRef]
- Akk, G.; Shu, H.J.; Wang, C.; Steinbach, J.H.; Zorumski, C.F.; Covey, D.F.; Mennerick, S. Neurosteroid access to the GABAA receptor. J. Neurosci. 2005, 25, 11605–11613. [Google Scholar]
- Chisari, M.; Eisenman, L.N.; Covey, D.F.; Mennerick, S.; Zorumski, C.F. The sticky issue of neurosteroids and GABA(A) receptors. Trends Neurosci. 2010, 33, 299–306. [Google Scholar]
- Taylor, G.T.; Griffin, M.G.; Bardgett, M. Search for a male contraceptive: The effect of gossypol on sexual motivation and epididymal sperm. J. Med. 1991, 22, 29–44. [Google Scholar]
- Taylor, G.T.; Komitowski, D.; Muto, S.; Weiss, J. Behavioral endocrinology of male rats with periodic sexual contacts with exclusive or varied females. Proc. Soc. Exp. Biol. Med. 1989, 192, 236–241. [Google Scholar]
- Farr, S.A.; Flood, J.F.; Scherrer, J.F.; Kaiser, F.E.; Taylor, G.T.; Morley, J.E. Effect of ovarian steroids on footshock avoidance learning and retention in female mice. Physiol. Behav. 1995, 58, 715–723. [Google Scholar]
- Lefevre, J.; McClintock, M.K. Reproductive senescence in female rats; a longitudial study of individual differences in estrous cycles and behavior. Biol. Reprod. 1988, 38, 780–789. [Google Scholar] [CrossRef]
- Taylor, G.T.; Haller, J.; Regan, D. Female rats prefer an area vacated by a high testosterone male. Physiol. Behav. 1982, 28, 953–958. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Taylor, G.T.; Dearborn, J.T.; Maloney, S.E. Adrenal Steroids Uniquely Influence Sexual Motivation Behavior in Male Rats. Behav. Sci. 2012, 2, 195-206. https://doi.org/10.3390/bs2030195
Taylor GT, Dearborn JT, Maloney SE. Adrenal Steroids Uniquely Influence Sexual Motivation Behavior in Male Rats. Behavioral Sciences. 2012; 2(3):195-206. https://doi.org/10.3390/bs2030195
Chicago/Turabian StyleTaylor, George T., Joshua T. Dearborn, and Susan E. Maloney. 2012. "Adrenal Steroids Uniquely Influence Sexual Motivation Behavior in Male Rats" Behavioral Sciences 2, no. 3: 195-206. https://doi.org/10.3390/bs2030195
APA StyleTaylor, G. T., Dearborn, J. T., & Maloney, S. E. (2012). Adrenal Steroids Uniquely Influence Sexual Motivation Behavior in Male Rats. Behavioral Sciences, 2(3), 195-206. https://doi.org/10.3390/bs2030195