
Effects of Early Weaning Associated with Alimentary Stress on Emotional and Feeding Behavior of Female Adult Wistar Rats

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Animals and Experimental Protocol
2.2.1. Animals
2.2.2. Early Weaning and Experimental Groups
- Control Group—weaned on a period natural for its species (30th post-natal day, n = 8);
- Early Weaning Group—weaned on the 15th post-natal day (n = 8).
2.3. Behavioral Procedures
2.3.1. Alimentary Stress
2.3.2. Cafeteria Diet Intake before and after Stress
2.3.3. Body Weight
2.3.4. Experimental Anxiety before and after Alimentary Stress
2.3.5. Standard Diet Intake after Alimentary Stress
2.3.6. Water Intake after Alimentary Stress
2.4. Statistical Analysis
3. Results
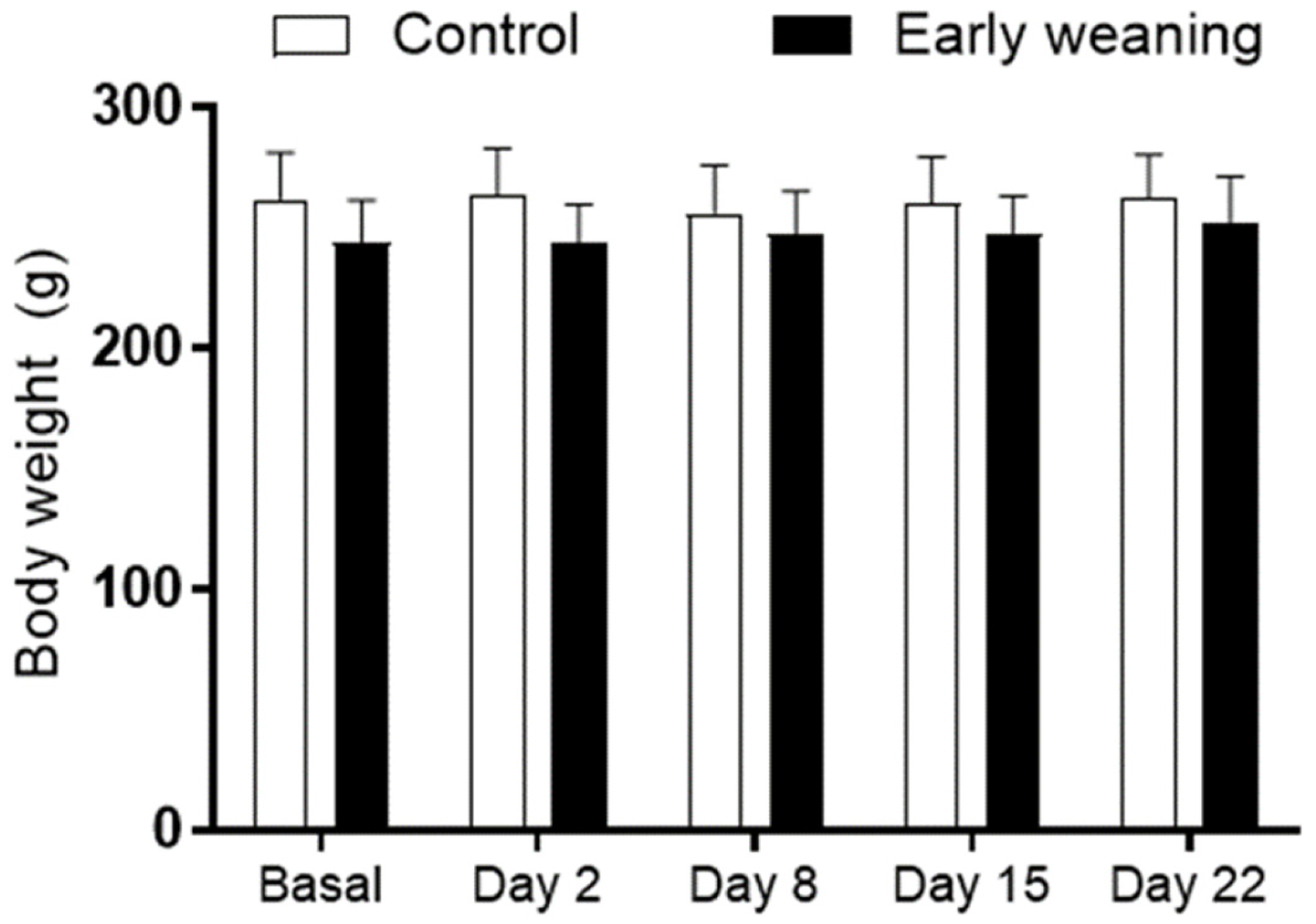
3.1. Body Weight
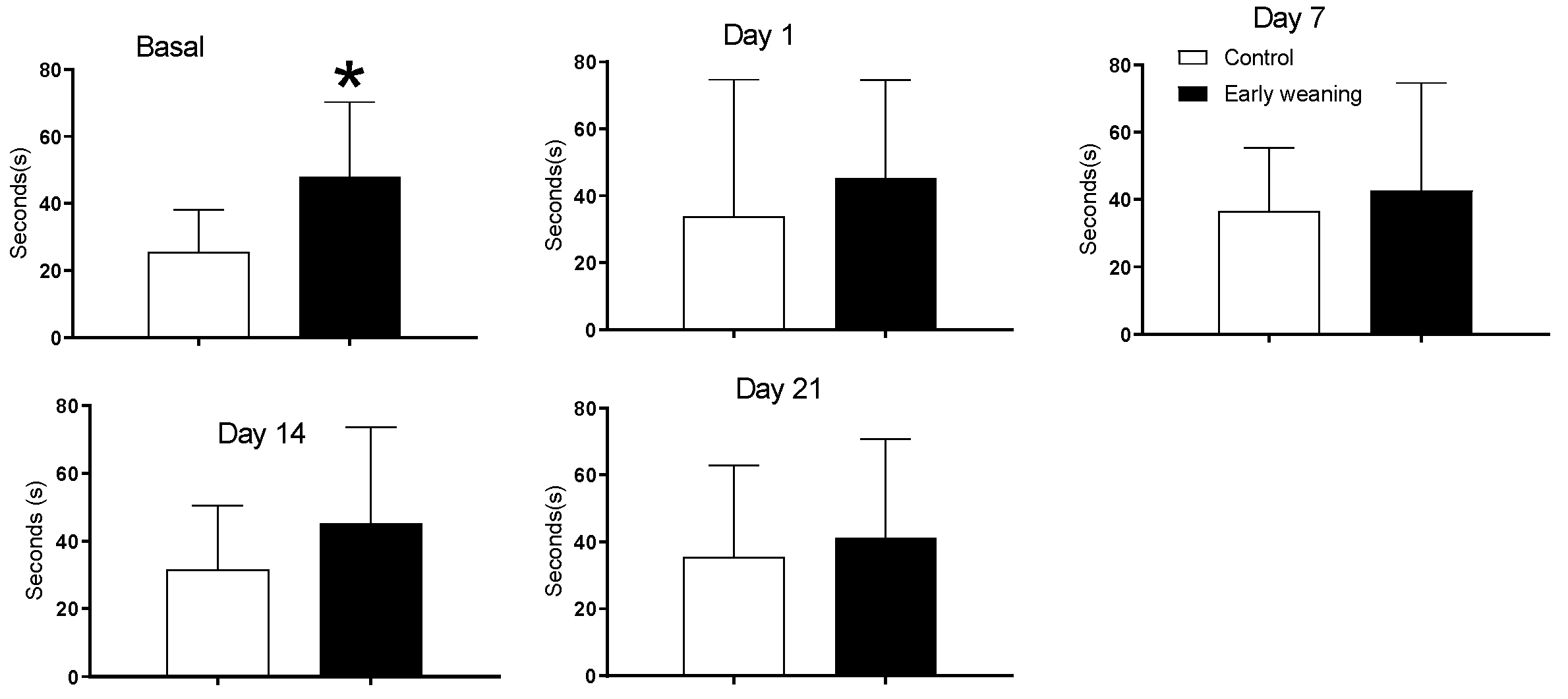
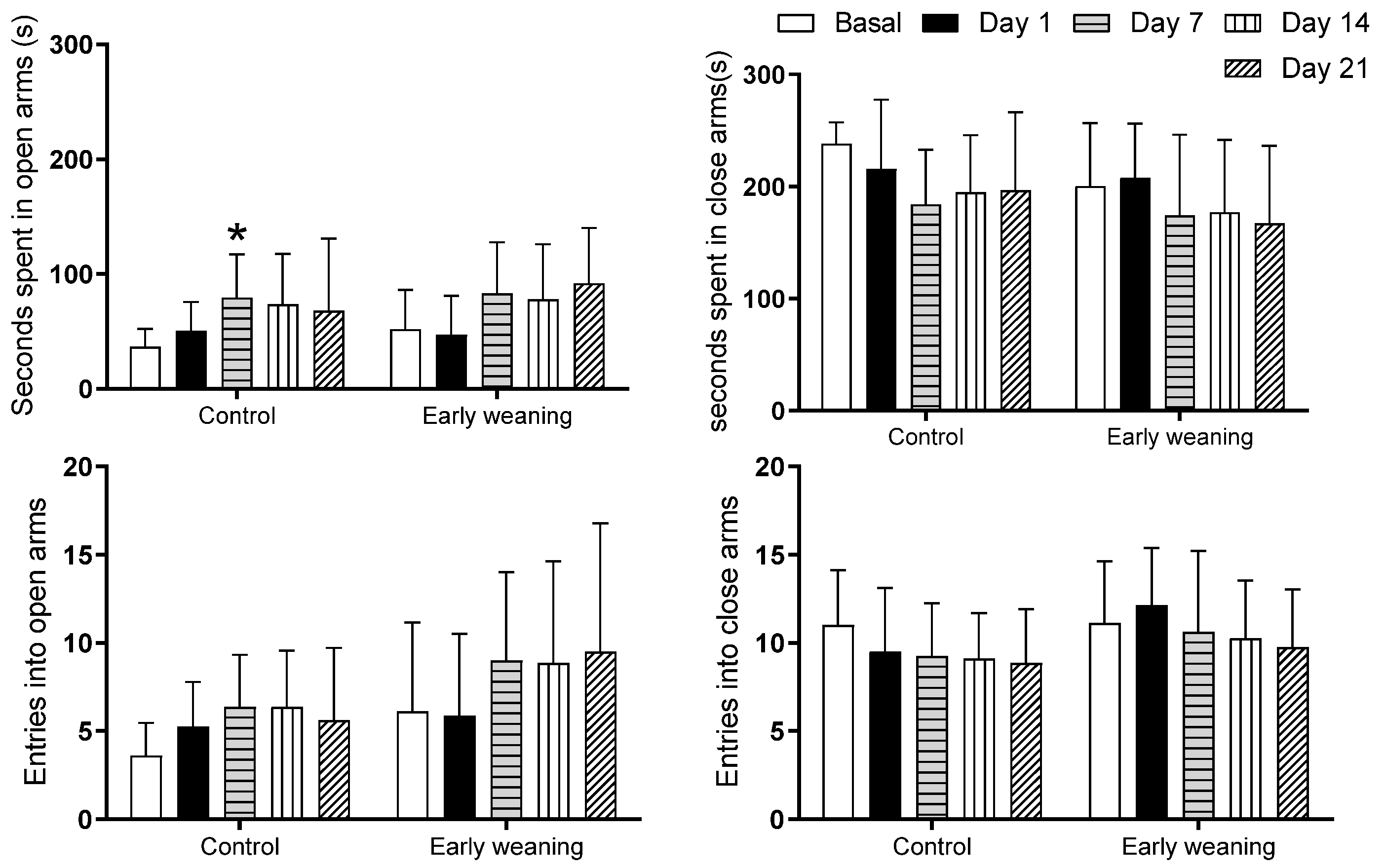
3.2. Elevated plus Maze
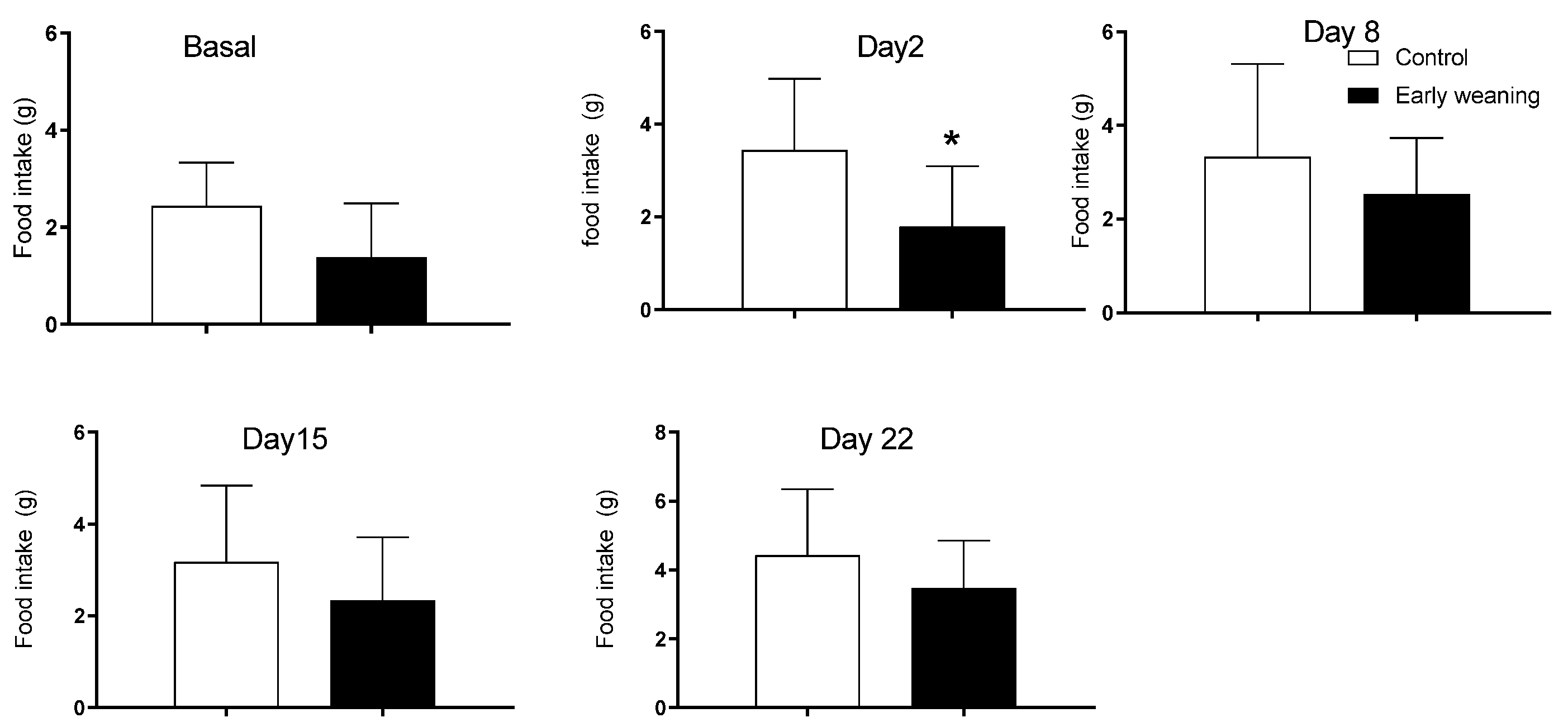
3.3. Cafeteria Diet Intake
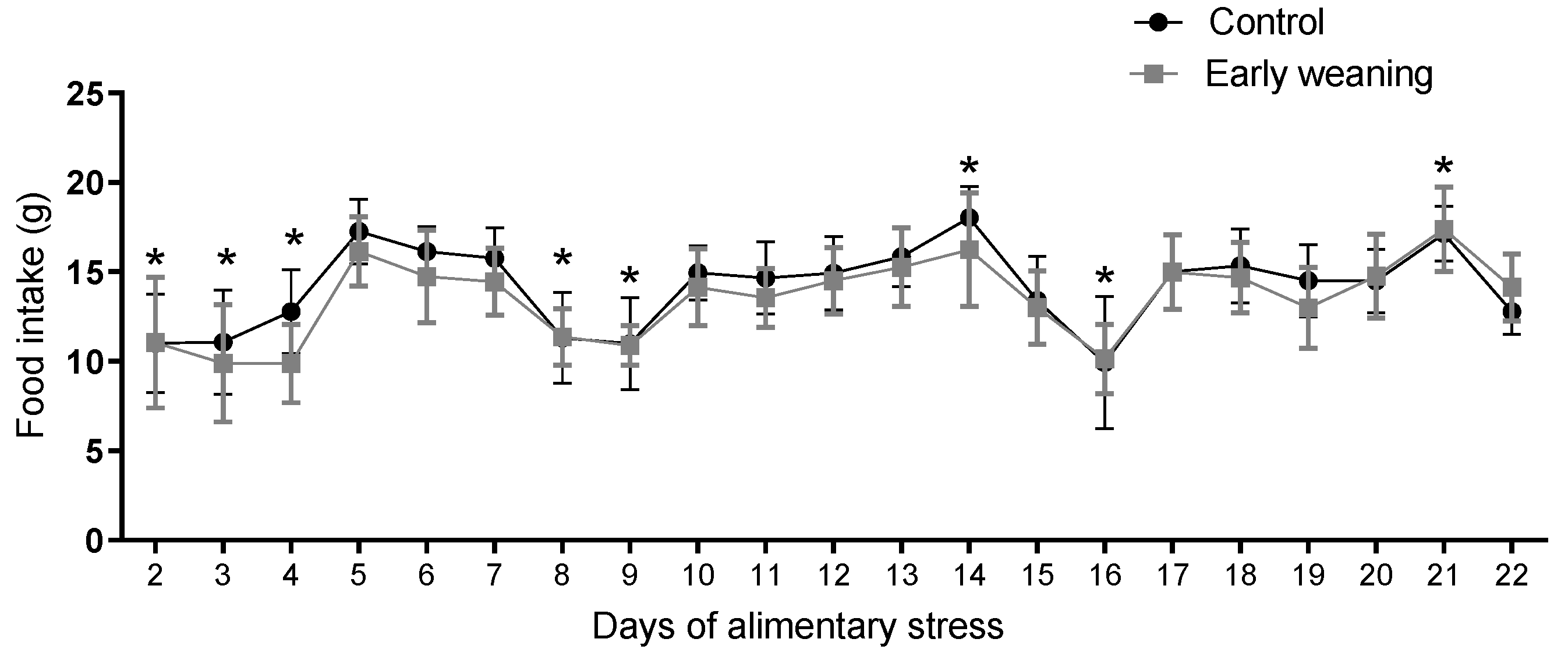
3.4. Standard Diet Intake
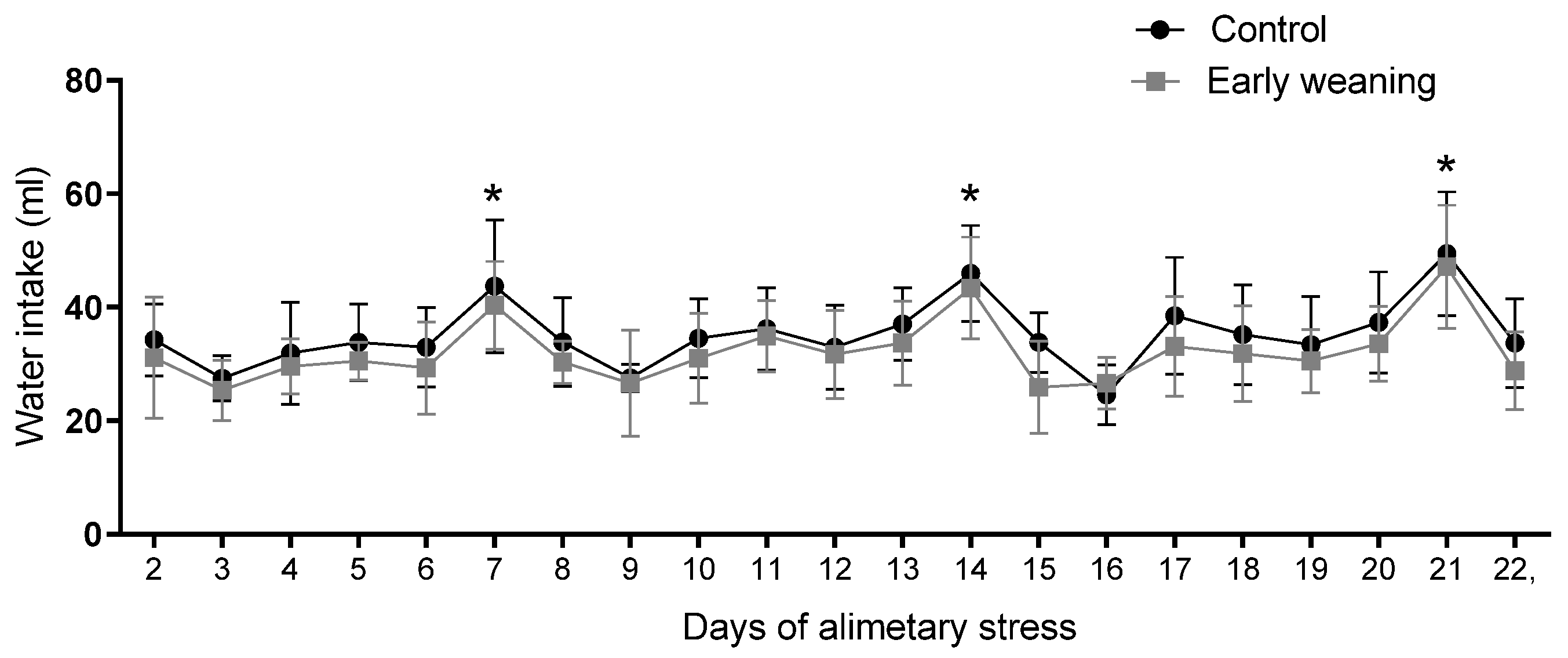
3.5. Water Intake
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EW | Early Weaning |
| E2 | 17β-Estradiol |
| EPM | Elevated Plus Maze |
| NPY | Neuropeptide Y |
| AgRp | Agouti-Related Protein |
| GABA | γ-aminobutyric Acid |
| mGluR5 | Metabotropic Glutamatergic Receptor 5 |
| ERα | Estrogen Receptor Alpha |
| OECD | Organization for Economic Cooperation and Development |
| UFPE | Federal University of Pernambuco |
| CEUA | Ethics Committee on the Use of Animals |
| CAPES | Coordination for Improvement of Higher Education Personnel |
| CNPq | Brazilian National Council for Scientific and Technological Development |
| PROCAD | National Academic Cooperation Program |
References
- Report of the Task Force on the Assessment of the Scientific Evidence Relating to Infant-Feeding Practices and Infant Health. Pediatrics 1984, 74, 579–762.
- Giugliani, E.R.J.; Horta, B.L.; Loret de Mola, C.; Lisboa, B.O.; Victora, C.G. Effect of breastfeeding promotion interventions on child growth: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Bowatte, G.; Tham, R.; Allen, K.J.; Tan, D.J.; Lau, M.; Dai, X.; Lodge, C.J. Breastfeeding and childhood acute otitis media: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 85–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sankar, M.J.; Sinha, B.; Chowdhury, R.; Bhandari, N.; Taneja, S.; Martines, J.; Bahl, R. Optimal breastfeeding practices and infant and child mortality: A systematic review and meta-analysis. Acta Paediatr. 2015, 104, 3–13. [Google Scholar] [CrossRef]
- Ventura, A.K. Does Breastfeeding Shape Food Preferences? Links to Obesity. Ann. Nutr. Metab. 2017, 70 (Suppl. S3), 8–15. [Google Scholar] [CrossRef]
- Kanari, K.; Kikusui, T.; Takeuchi, Y.; Mori, Y. Multidimensional structure of anxiety-related behavior in early-weaned rats. Behav. Brain Res. 2005, 156, 45–52. [Google Scholar] [CrossRef]
- Lähdepuro, A.; Savolainen, K.; Lahti-Pulkkinen, M.; Eriksson, J.G.; Lahti, J.; Tuovinen, S.; Kajantie, E.; Pesonen, A.-K.; Heinonen, K.; Räikkönen, K. The Impact of Early Life Stress on Anxiety Symptoms in Late Adulthood. Sci. Rep. 2019, 9, 4395. [Google Scholar] [CrossRef] [Green Version]
- Du Percie Sert, N.; Hurst, V.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; et al. The ARRIVE guidelines 2.0: Updated guidelines for reporting animal research. PLoS Biol. 2020, 18, e3000410. [Google Scholar] [CrossRef]
- Lima, N.d.S.; Moura, E.G.d.; Passos, M.C.F.; Nogueira Neto, F.J.; Reis, A.M.; Oliveira, E.d.; Lisboa, P.C. Early weaning causes undernutrition for a short period and programmes some metabolic syndrome components and leptin resistance in adult rat offspring. Br. J. Nutr. 2011, 105, 1405–1413. [Google Scholar] [CrossRef] [Green Version]
- Baquero, A.F.; Solis, A.J.d.; Lindsley, S.R.; Kirigiti, M.A.; Smith, M.S.; Cowley, M.A.; Zeltser, L.M.; Grove, K.L. Developmental switch of leptin signaling in arcuate nucleus neurons. J. Neurosci. 2014, 34, 9982–9994. [Google Scholar] [CrossRef] [Green Version]
- Tavares, G.A.; do Amaral Almeida, L.C.; Souza, J.A.d.; Farias, V.V.d.; Souza, F.L.d.; Andrade Silva, S.C.d.; Lagranha, C.J.; Kaeffer, B.; Souza, S.L.d. Early weaning leads to disruption of homeostatic and hedonic eating behaviors and modulates serotonin (5HT) and dopamine (DA) systems in male adult rats. Behav. Brain Res. 2020, 383, 112531. [Google Scholar] [CrossRef] [PubMed]
- Tarttelin, M.F.; Gorski, R.A. The effects of ovarian steroids on food and water intake and body weight in the female rat. Acta Endocrinol. 1973, 72, 551–568. [Google Scholar] [CrossRef] [PubMed]
- Tarttelin, M.F. Cyclical variations in food and water intakes in ewes. J. Physiol. 1968, 195, 29P–31P. [Google Scholar] [PubMed]
- Tavares, G.A.; Almeida, L.C.d.A.; Souza, J.A.d.; Braz, G.R.F.; da Silva, M.C.; Lagranha, C.J.; do Nascimento, E.; Souza, S.L.d. Early weaning disrupts feeding patterns in female juvenile rats through 5HT-system modulations. Behav. Processes 2020, 170, 103981. [Google Scholar] [CrossRef]
- Steger, R.W.; Peluso, J.J. Effects of age on hormone levels and in vitro steroidogenesis by rat ovary and adrenal. Exp. Aging Res. 1982, 8, 203–208. [Google Scholar] [CrossRef]
- Asarian, L.; Geary, N. Cyclic estradiol treatment normalizes body weight and restores physiological patterns of spontaneous feeding and sexual receptivity in ovariectomized rats. Horm. Behav. 2002, 42, 461–471. [Google Scholar] [CrossRef]
- Wallen, W.J.; Belanger, M.P.; Wittnich, C. Sex hormones and the selective estrogen receptor modulator tamoxifen modulate weekly body weights and food intakes in adolescent and adult rats. J. Nutr. 2001, 131, 2351–2357. [Google Scholar] [CrossRef]
- Wade, G.N. Some effects of ovarian hormones on food intake and body weight in female rats. J. Comp. Physiol. Psychol. 1975, 88, 183–193. [Google Scholar] [CrossRef]
- Eckel, L.A.; Langhans, W.; Kahler, A.; Campfield, L.A.; Smith, F.J.; Geary, N. Chronic administration of OB protein decreases food intake by selectively reducing meal size in female rats. Am. J. Physiol. 1998, 275, R186–R193. [Google Scholar] [CrossRef]
- Martínez de Morentin, P.B.; González-García, I.; Martins, L.; Lage, R.; Fernández-Mallo, D.; Martínez-Sánchez, N.; Ruíz-Pino, F.; Liu, J.; Morgan, D.A.; Pinilla, L.; et al. Estradiol regulates brown adipose tissue thermogenesis via hypothalamic AMPK. Cell Metab. 2014, 20, 41–53. [Google Scholar] [CrossRef] [Green Version]
- South, T.; Holmes, N.M.; Martire, S.I.; Westbrook, R.F.; Morris, M.J. Rats eat a cafeteria-style diet to excess but eat smaller amounts and less frequently when tested with chow. PLoS ONE 2014, 9, e93506. [Google Scholar] [CrossRef] [Green Version]
- Chu, S.-C.; Chou, Y.-C.; Liu, J.-Y.; Chen, C.-H.; Shyu, J.-C.; Chou, F.-P. Fluctuation of serum leptin level in rats after ovariectomy and the influence of estrogen supplement. Life Sci. 1999, 64, 2299–2306. [Google Scholar] [CrossRef]
- Rivera, H.M.; Stincic, T.L. Estradiol and the control of feeding behavior. Steroids 2018, 133, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Drewett, R.F. Oestrous and dioestrous components of the ovarian inhibition on hunger in the rat. Anim. Behav. 1973, 21, 772–780. [Google Scholar] [CrossRef]
- Eckel, L. Spontaneous meal patterns in female rats with and without access to running wheels. Physiol. Behav. 2000, 70, 397–405. [Google Scholar] [CrossRef]
- Ahdieh, H.B.; Wade, G.N. Effects of hysterectomy on sexual receptivity, food intake, running wheel activity, and hypothalamic estrogen and progestin receptors in rats. J. Comp. Physiol. Psychol. 1982, 96, 886–892. [Google Scholar] [CrossRef]
- Gruber, C.J.; Gruber, D.M.; Gruber, I.M.L.; Wieser, F.; Huber, J.C. Anatomy of the estrogen response element. Trends Endocrinol. Metab. 2004, 15, 73–78. [Google Scholar] [CrossRef]
- Qiu, J.; Bosch, M.A.; Tobias, S.C.; Krust, A.; Graham, S.M.; Murphy, S.J.; Korach, K.S.; Chambon, P.; Scanlan, T.S.; Rønnekleiv, O.K.; et al. A G-protein-coupled estrogen receptor is involved in hypothalamic control of energy homeostasis. J. Neurosci. 2006, 26, 5649–5655. [Google Scholar] [CrossRef] [Green Version]
- Carroll, M.E.; Morgan, A.D.; Lynch, W.J.; Campbell, U.C.; Dess, N.K. Intravenous cocaine and heroin self-administration in rats selectively bred for differential saccharin intake: Phenotype and sex differences. Psychopharmacology 2002, 161, 304–313. [Google Scholar] [CrossRef]
- Roberts, D.C.; Bennett, S.A.; Vickers, G.J. The estrous cycle affects cocaine self-administration on a progressive ratio schedule in rats. Psychopharmacology 1989, 98, 408–411. [Google Scholar] [CrossRef]
- Volkow, N.D.; Wang, G.-J.; Baler, R.D. Reward, dopamine and the control of food intake: Implications for obesity. Trends Cogn. Sci. 2011, 15, 37–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galea, L. High levels of estradiol disrupt conditioned place preference learning, stimulus response learning and reference memory but have limited effects on working memory. Behav. Brain Res. 2001, 126, 115–126. [Google Scholar] [CrossRef]
- Becker, J.B.; Hu, M. Sex differences in drug abuse. Front. Neuroendocrinol. 2008, 29, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richard, J.E.; López-Ferreras, L.; Anderberg, R.H.; Olandersson, K.; Skibicka, K.P. Estradiol is a critical regulator of food-reward behavior. Psychoneuroendocrinology 2017, 78, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.A.; Gross, K.S.; Himmler, B.T.; Emmitt, N.L.; Peterson, B.M.; Zlebnik, N.E.; Foster Olive, M.; Carroll, M.E.; Meisel, R.L.; Mermelstein, P.G. Estradiol Facilitation of Cocaine Self-Administration in Female Rats Requires Activation of mGluR5. Eneuro 2016, 3, 0140-16. [Google Scholar] [CrossRef]
- Cason, A.M.; Aston-Jones, G. Role of orexin/hypocretin in conditioned sucrose-seeking in female rats. Neuropharmacology 2014, 86, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Ito, A.; Kikusui, T.; Takeuchi, Y.; Mori, Y. Effects of early weaning on anxiety and autonomic responses to stress in rats. Behav. Brain Res. 2006, 171, 87–93. [Google Scholar] [CrossRef]
- Shimozuru, M.; Kodama, Y.; Iwasa, T.; Kikusui, T.; Takeuchi, Y.; Mori, Y. Early weaning decreases play-fighting behavior during the postweaning developmental period of Wistar rats. Dev. Psychobiol. 2007, 49, 343–350. [Google Scholar] [CrossRef]
- Campos, A.C.; Fogaça, M.V.; Aguiar, D.C.; Guimarães, F.S. Animal models of anxiety disorders and stress. Braz. J. Psychiatry 2013, 35 (Suppl. S2), S101–S111. [Google Scholar] [CrossRef] [Green Version]
- Schrader, A.J.; Taylor, R.M.; Lowery-Gionta, E.G.; Moore, N.L.T. Repeated elevated plus maze trials as a measure for tracking within-subjects behavioral performance in rats (Rattus norvegicus). PLoS ONE 2018, 13, e0207804. [Google Scholar] [CrossRef] [Green Version]
- Kastenberger, I.; Lutsch, C.; Schwarzer, C. Activation of the G-protein-coupled receptor GPR30 induces anxiogenic effects in mice, similar to oestradiol. Psychopharmacology 2012, 221, 527–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrow, A.P.; Handa, R.J. Estrogen Receptors Modulation of Anxiety-Like Behavior. Vitam. Horm. 2017, 103, 27–52. [Google Scholar] [CrossRef] [PubMed]
- Rico, J.L.; Hurtado-Parrado, C.; Vásquez-Sepúlveda, J.; Fonseca, J.; Cardona, Á. El tiempo en el área central del laberinto en cruz elevado correlaciona con medidas relacionadas a impulsividad durante una tarea operante. Univ. Psychol. 2017, 15, 1–9. [Google Scholar] [CrossRef]
- Stein, J.S.; Renda, C.R.; Barker, S.M.; Liston, K.J.; Shahan, T.A.; Madden, G.J. Impulsive choice predicts anxiety-like behavior, but not alcohol or sucrose consumption, in male Long-Evans rats. Alcohol. Clin. Exp. Res. 2015, 39, 932–940. [Google Scholar] [CrossRef] [Green Version]
- Jakuszkowiak-Wojten, K.; Landowski, J.; Wiglusz, M.S.; Cubała, W.J. Impulsivity in anxiety disorders. A critical review. Psychiatr. Danub. 2015, 27 (Suppl. S1), S452–S455. [Google Scholar]
- Fitzsimons, T.J.; Le Magnen, J. Eating as a regulatory control of drinking in the rat. J. Comp. Physiol. Psychol. 1969, 67, 273–283. [Google Scholar] [CrossRef]
- Shughrue, P.J.; Lane, M.V.; Merchenthaler, I. Comparative distribution of estrogen receptor-α and-β mRNA in the rat central nervous system. J. Comp. Neurol. 1997, 388, 507–525. [Google Scholar] [CrossRef]
- Suzuki, T.; Kohno, H.; Sakurada, T.; Tadano, T.; Kisara, K. Intracranial injection of thyrotropin releasing hormone (TRH) suppresses starvation-induced feeding and drinking in rats. Pharmacol. Biochem. Behav. 1982, 17, 249–253. [Google Scholar] [CrossRef]
- Choi, Y.-H.; Hartzell, D.; Azain, M.J.; Baile, C.A. TRH decreases food intake and increases water intake and body temperature in rats. Physiol. Behav. 2002, 77, 1–4. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz, V.I.M.; de Souza, J.A.; de Sousa, S.L. Effects of Early Weaning Associated with Alimentary Stress on Emotional and Feeding Behavior of Female Adult Wistar Rats. Behav. Sci. 2022, 12, 171. https://doi.org/10.3390/bs12060171
Díaz VIM, de Souza JA, de Sousa SL. Effects of Early Weaning Associated with Alimentary Stress on Emotional and Feeding Behavior of Female Adult Wistar Rats. Behavioral Sciences. 2022; 12(6):171. https://doi.org/10.3390/bs12060171
Chicago/Turabian StyleDíaz, Víctor Isaac Meléndez, Julliet Araújo de Souza, and Sandra Lopes de Sousa. 2022. "Effects of Early Weaning Associated with Alimentary Stress on Emotional and Feeding Behavior of Female Adult Wistar Rats" Behavioral Sciences 12, no. 6: 171. https://doi.org/10.3390/bs12060171
APA StyleDíaz, V. I. M., de Souza, J. A., & de Sousa, S. L. (2022). Effects of Early Weaning Associated with Alimentary Stress on Emotional and Feeding Behavior of Female Adult Wistar Rats. Behavioral Sciences, 12(6), 171. https://doi.org/10.3390/bs12060171