
The Impact of Chronic Unpredictable Mild Stress-Induced Depression on Spatial, Recognition and Reference Memory Tasks in Mice: Behavioral and Histological Study

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. CUMS Procedures
2.3. Experimental Design
2.4. Weight
2.5. Locomotor Behavioral Tests
OFT
2.6. Anxiety Behavioral Tests
2.6.1. Central Preference %
2.6.2. EZM
2.6.3. EPM
2.7. Depression Behavioral Tests
2.7.1. TST
2.7.2. FST
2.7.3. SPT
2.8. Memory Tests
2.8.1. NORT
2.8.2. Y-Maze
2.8.3. MWM
2.9. Tissue Harvesting and Processing
2.10. Statistical Analysis
3. Results
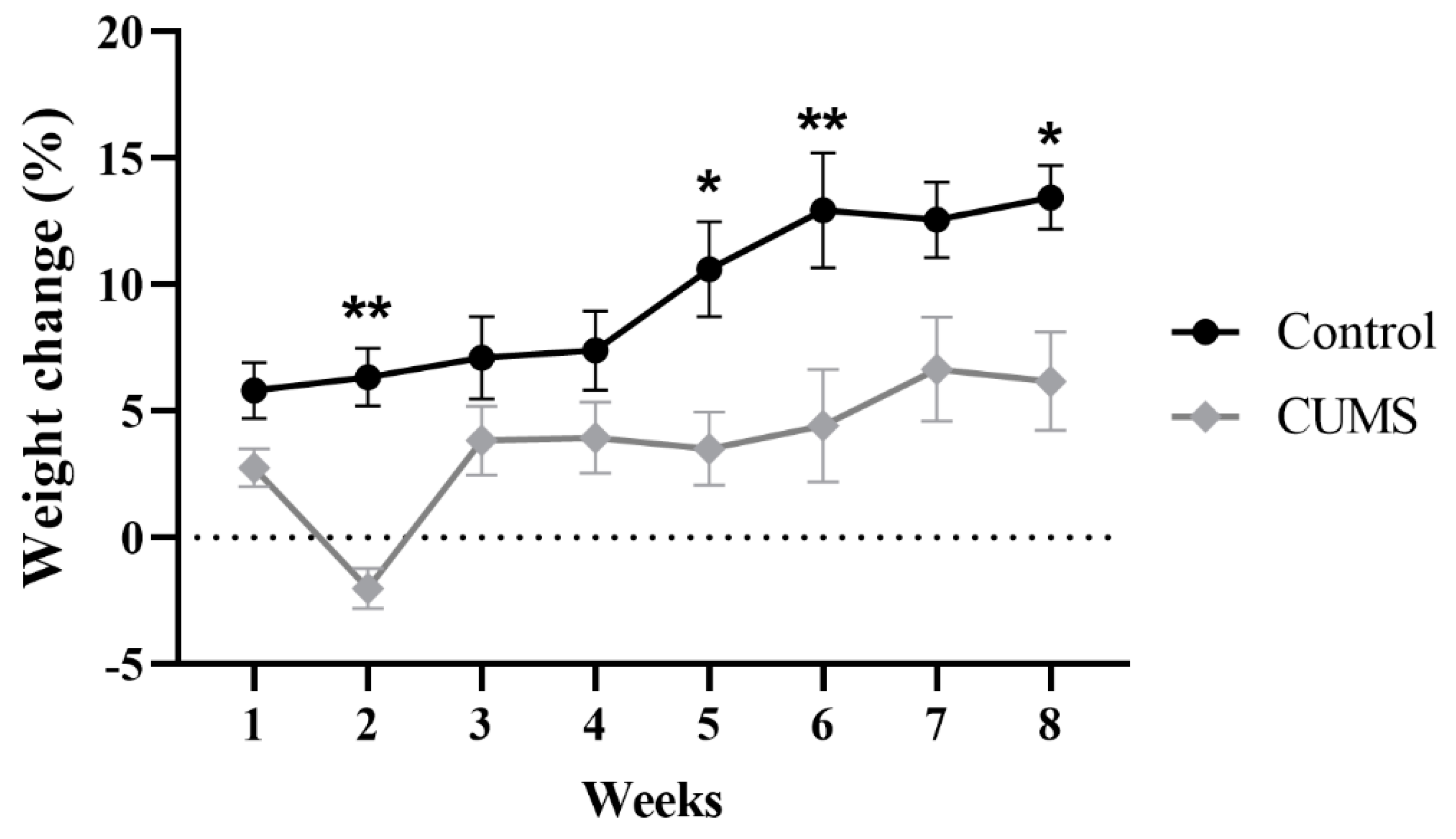
3.1. Effect of CUMS on Body Weight
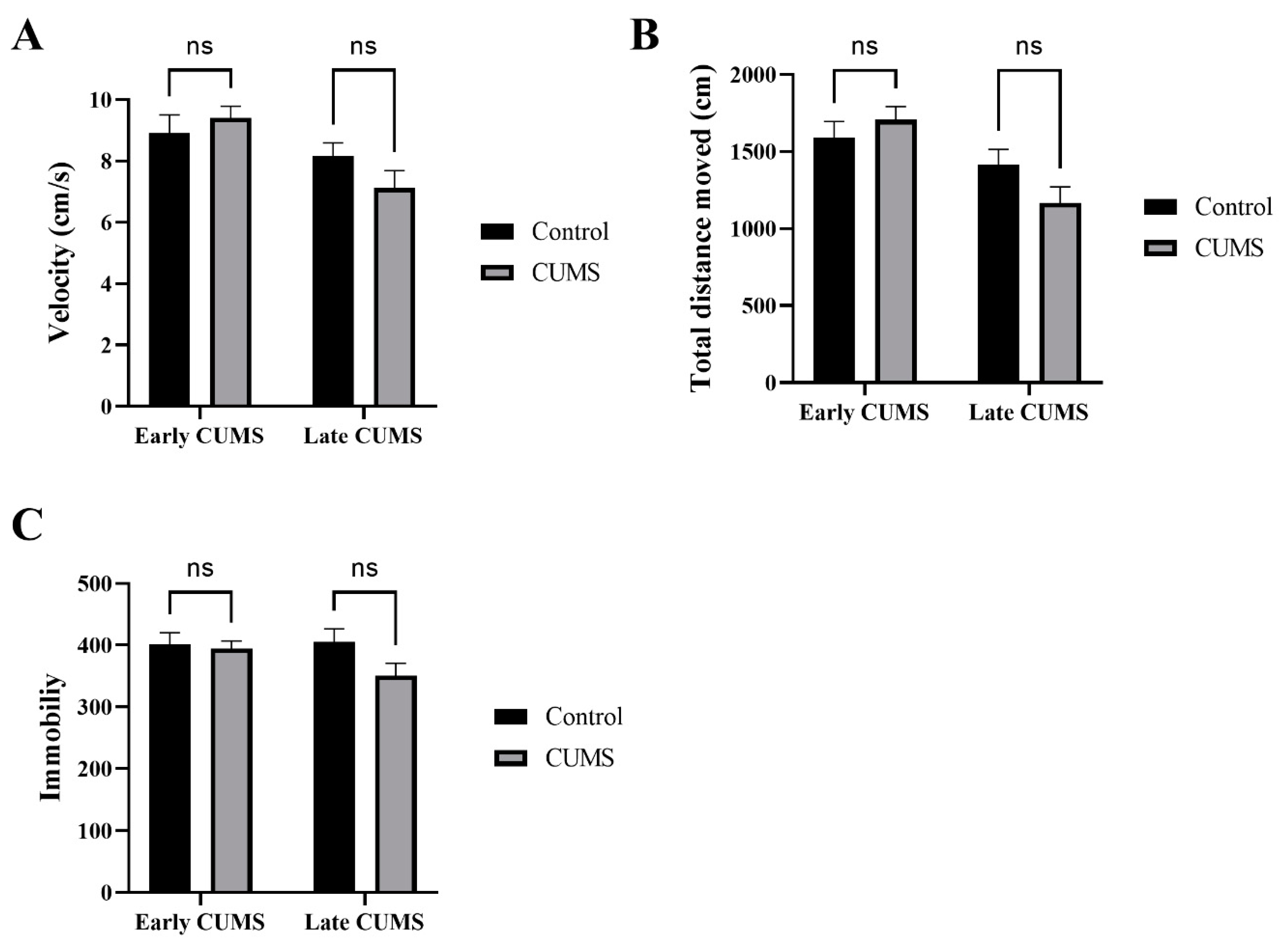
3.2. Effect of Early and Late CUMS on Locomotor Activity
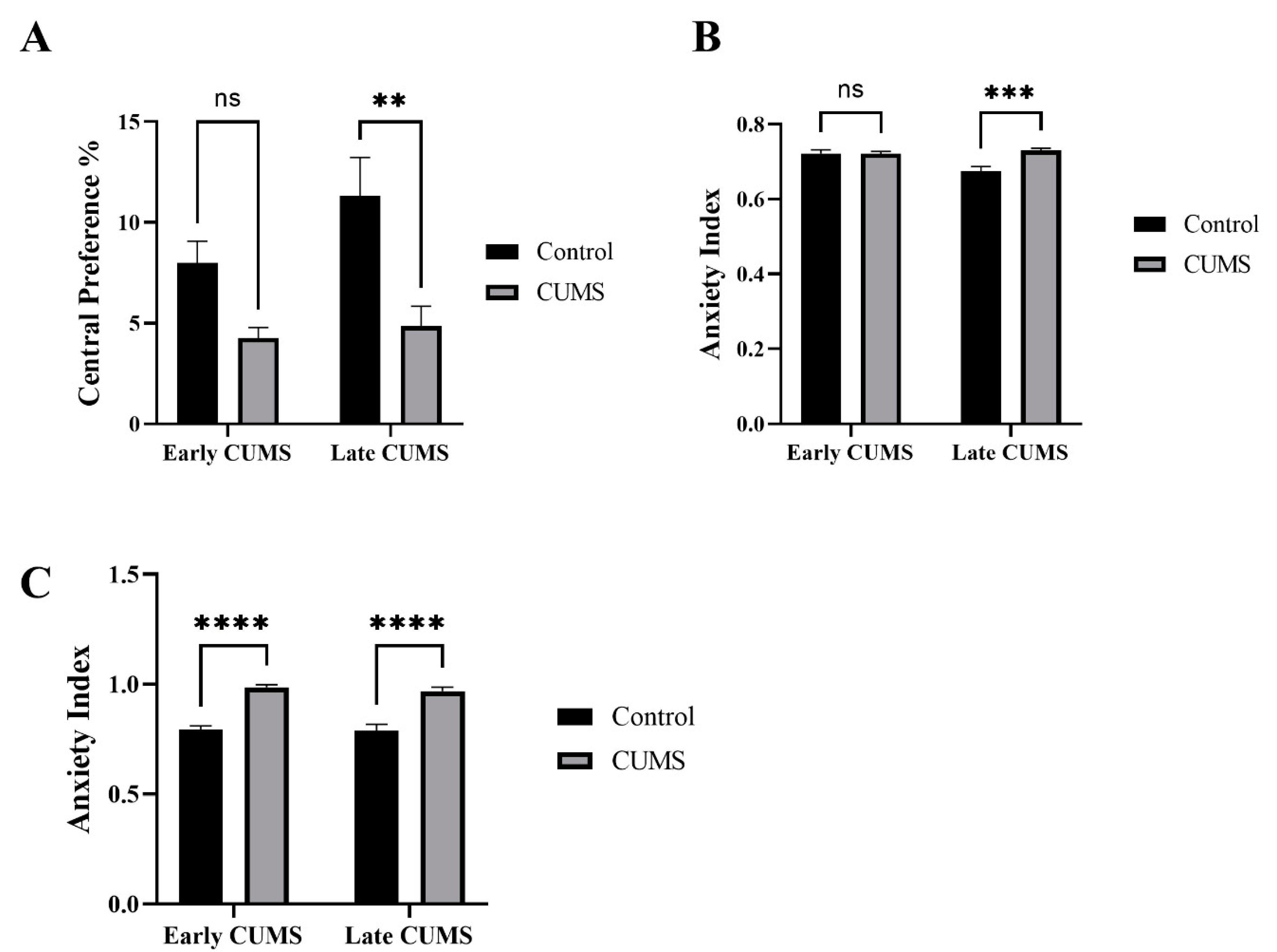
3.3. Effect of Early and Late CUMS on Anxiety Behavior
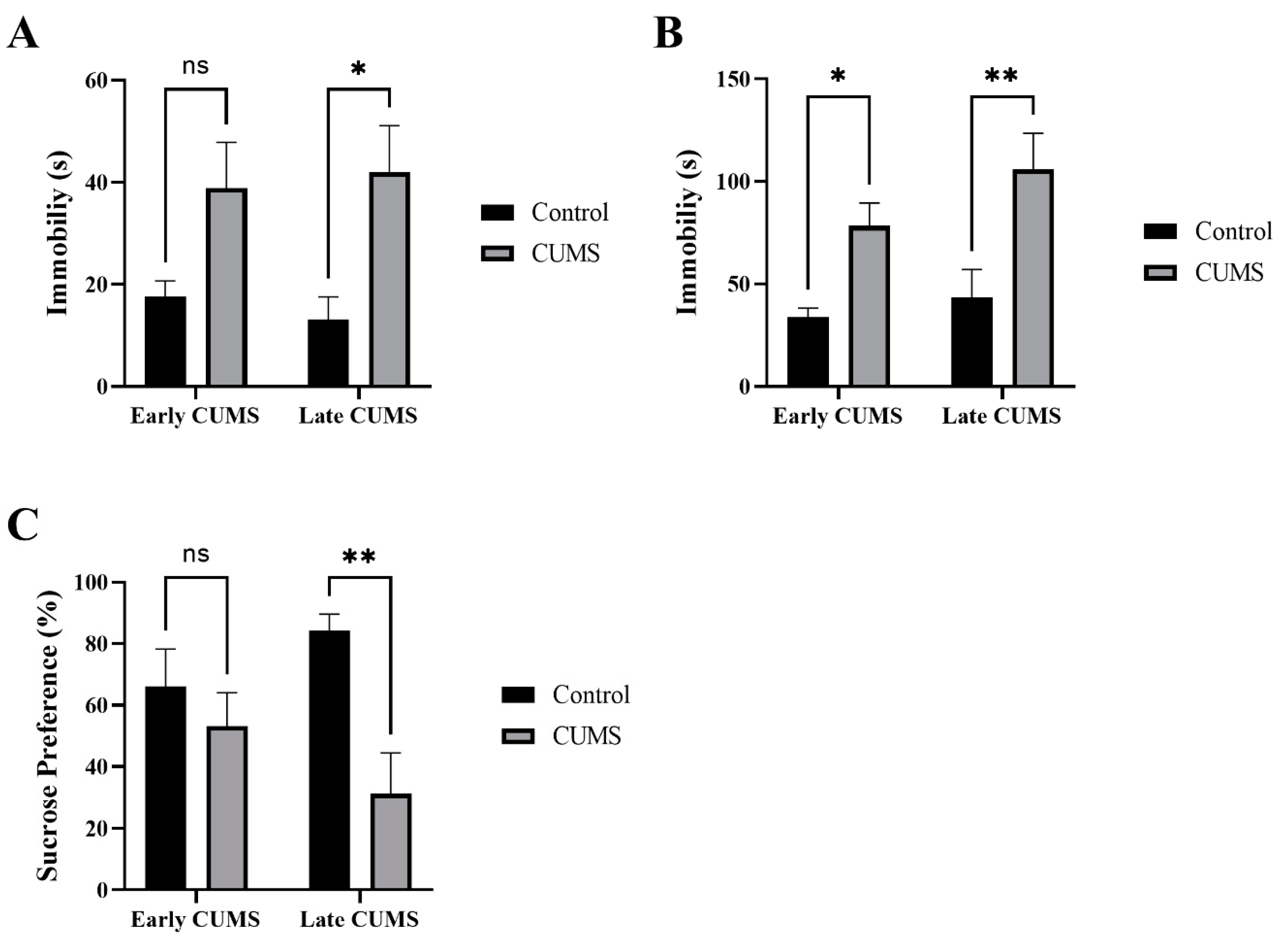
3.4. Effect of Early and Late CUMS on Depression Behavior
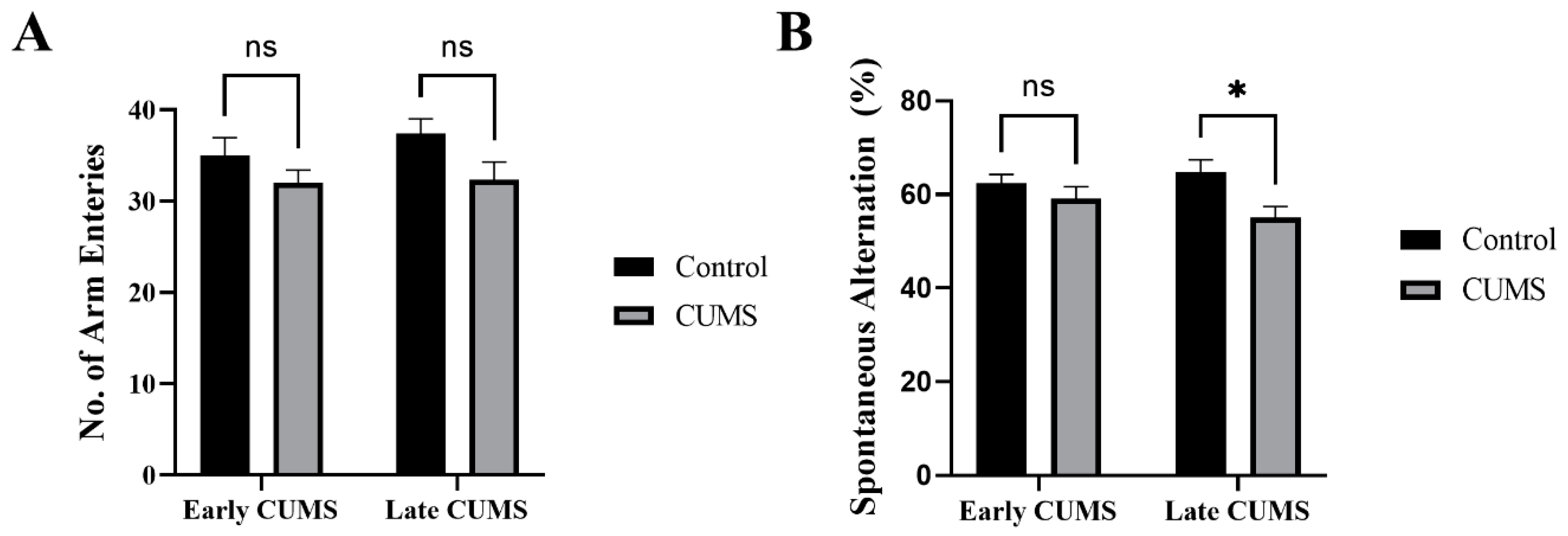
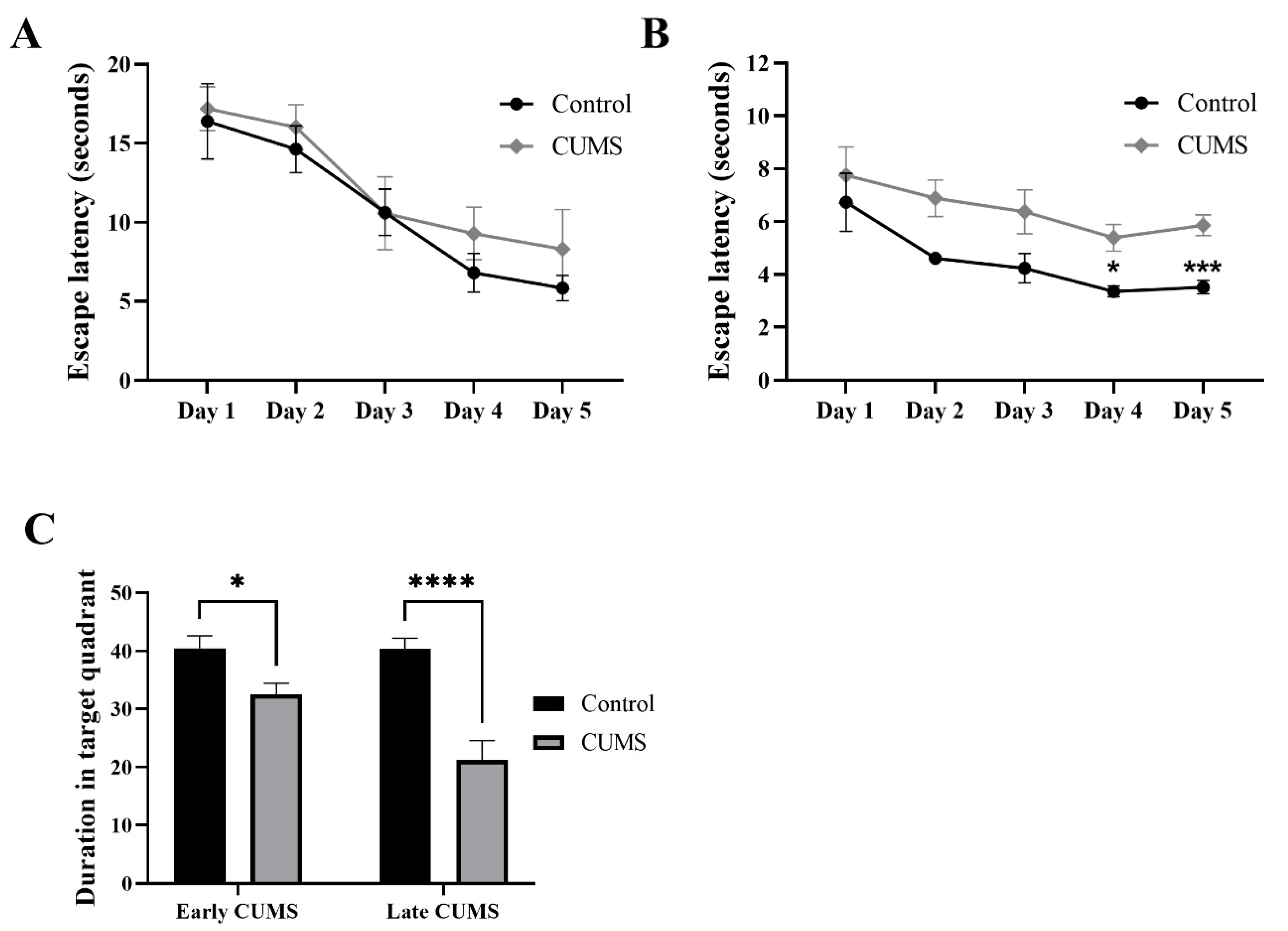
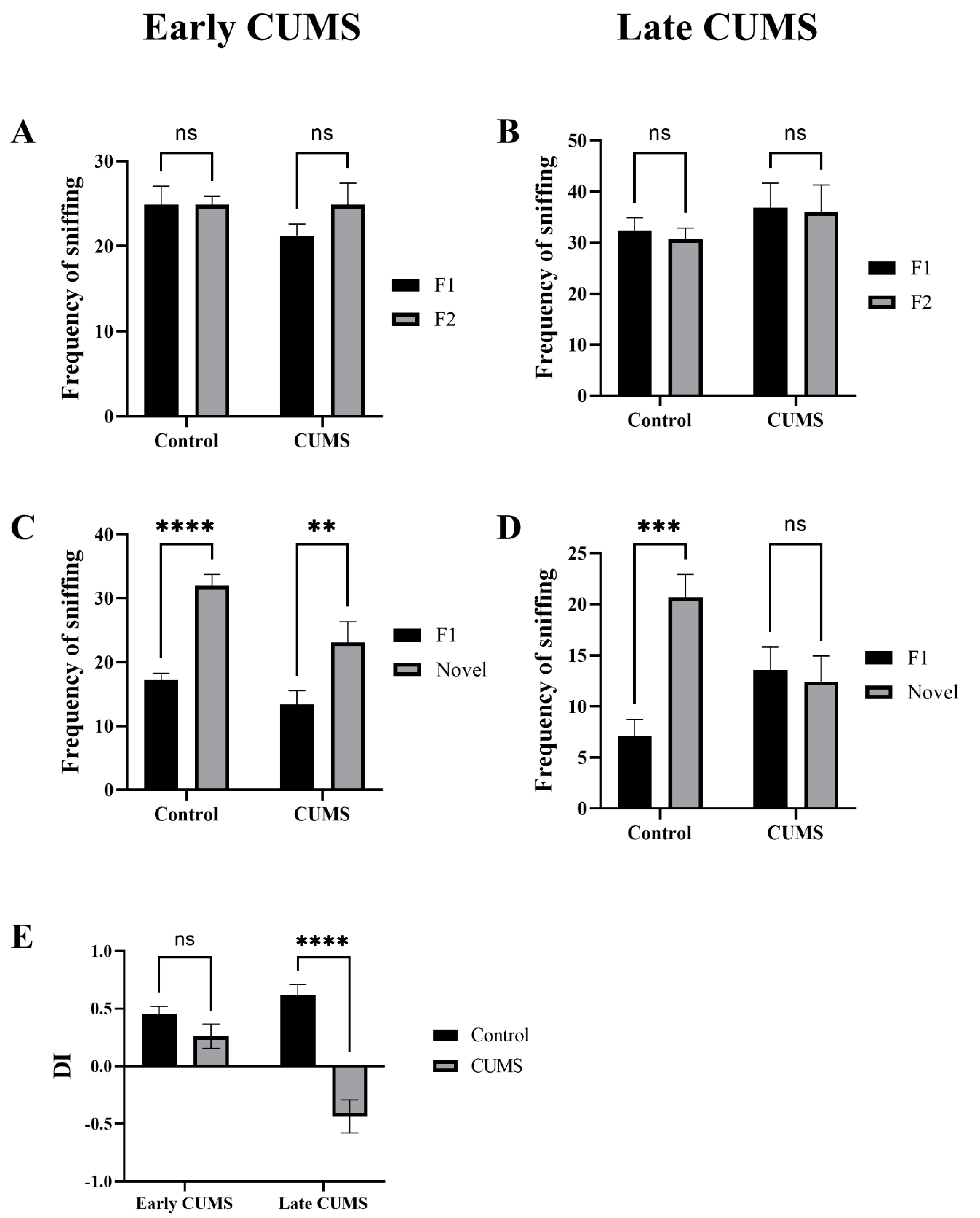
3.5. Effect of Early and Late CUMS on Memory
3.5.1. Effect of Early and Late CUMS on Spatial Memory
3.5.2. Effect of Early and Late CUMS on Recognition Memory
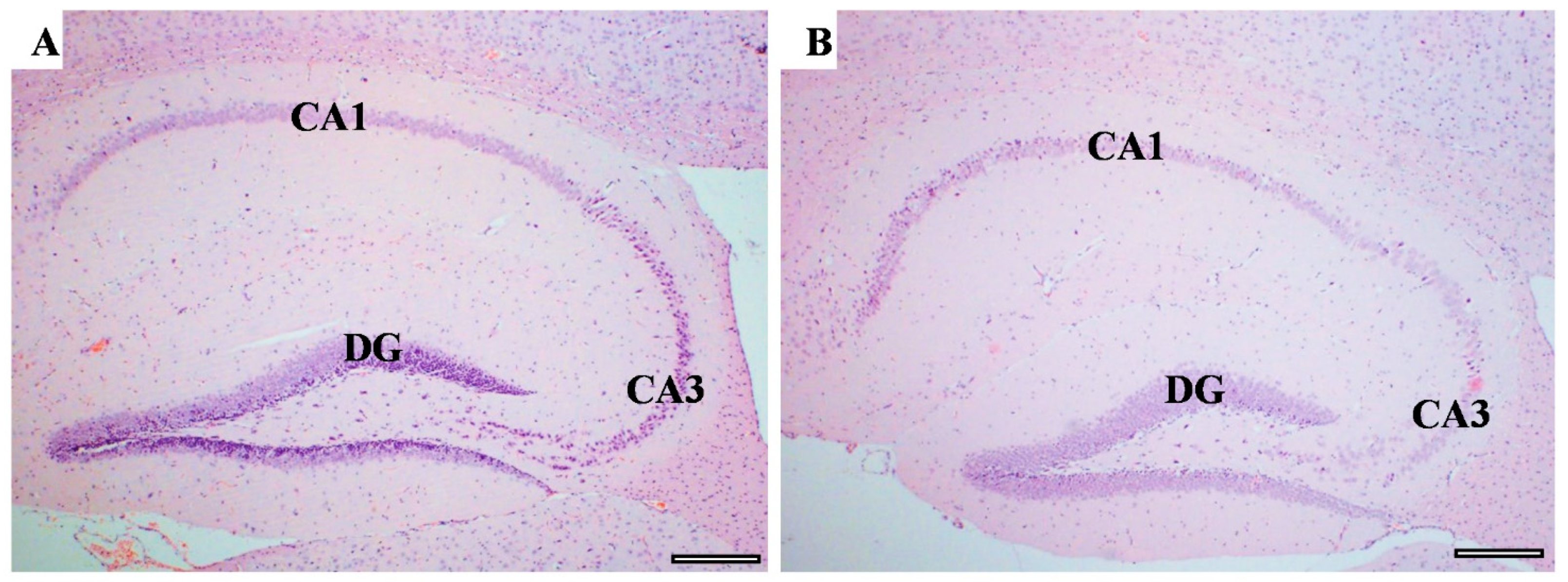
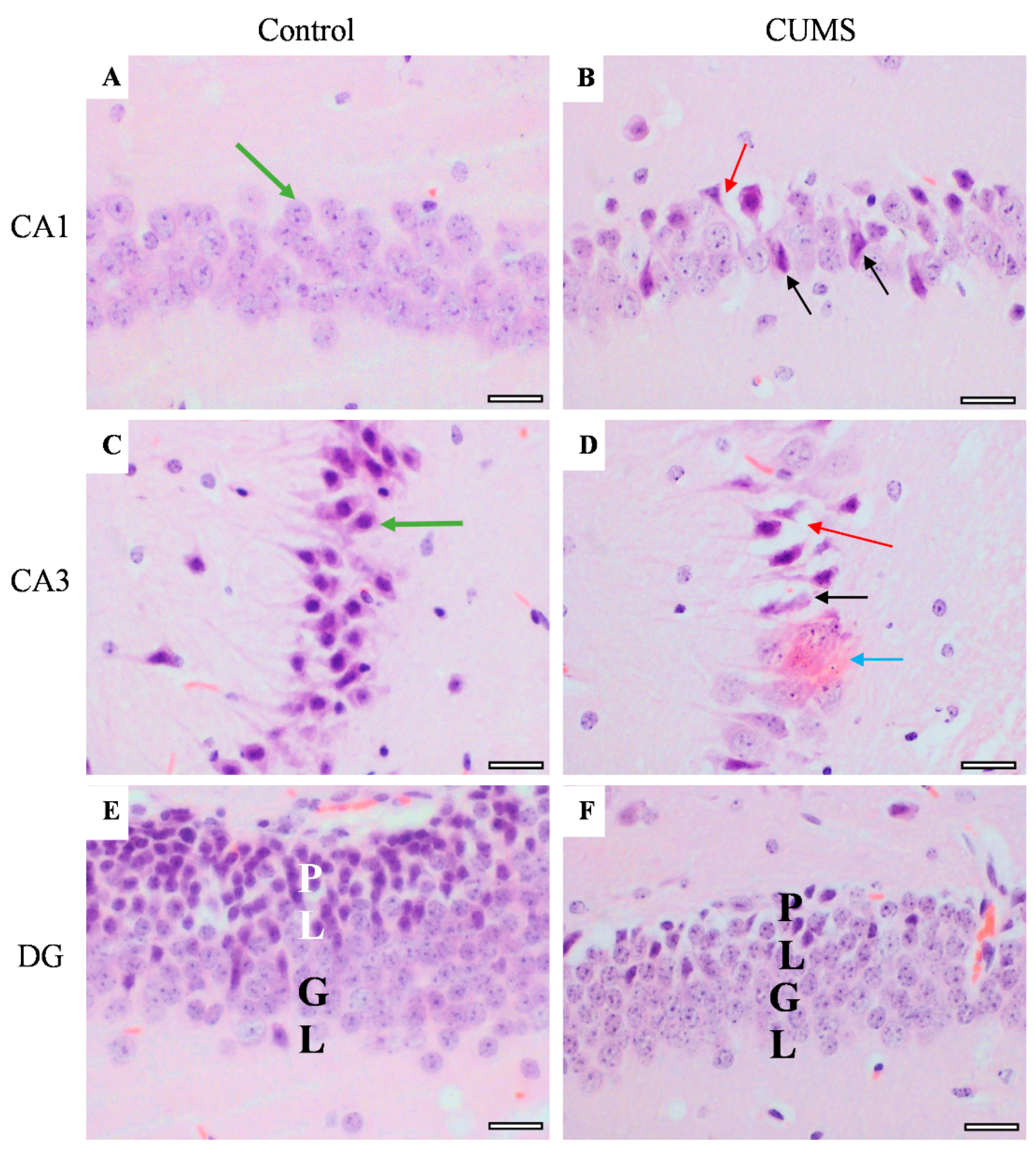
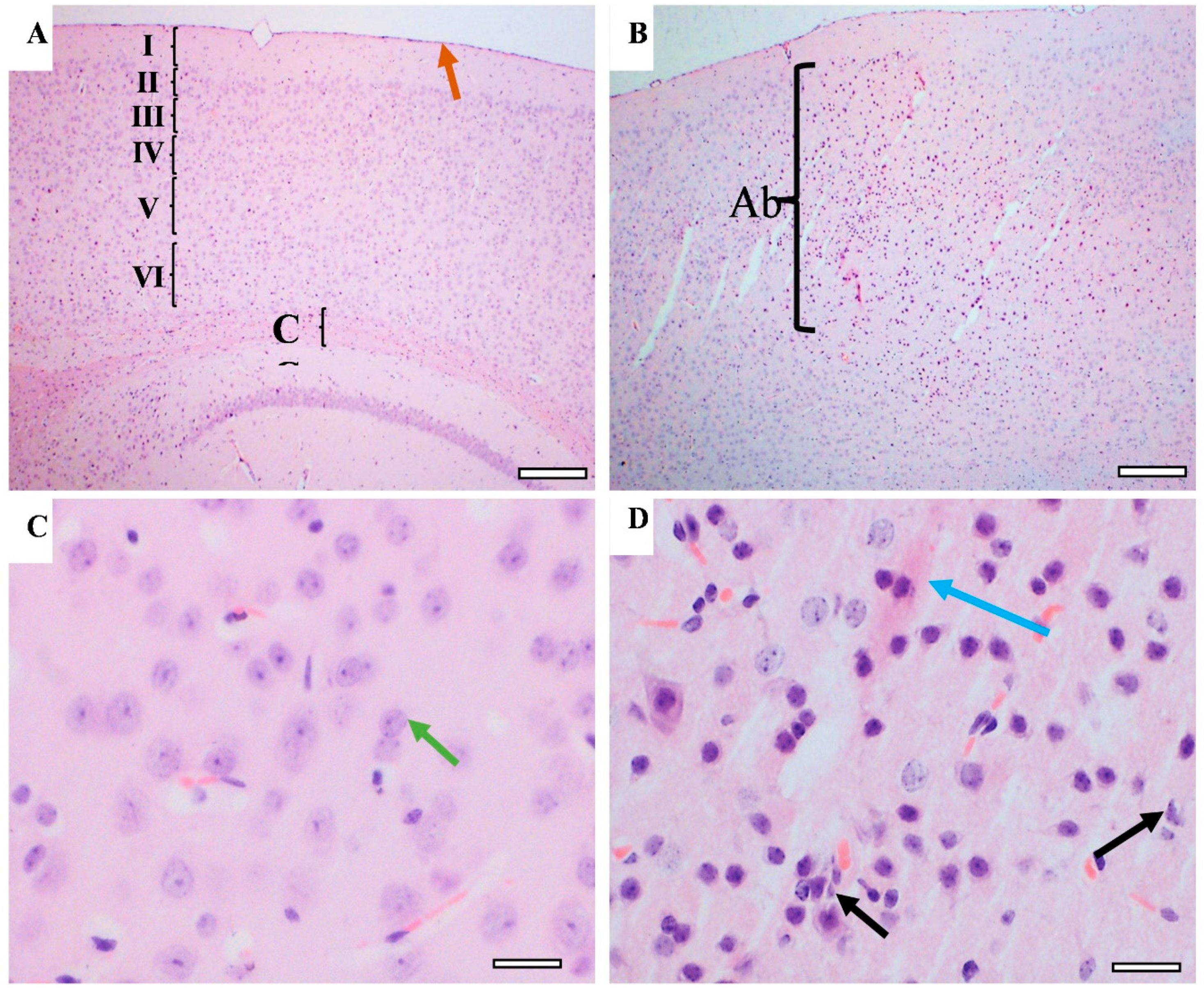
3.6. Effect of CUMS on the Brain Histological Feature
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Melchior, M.; Caspi, A.; Milne, B.J.; Danese, A.; Poulton, R.; Moffitt, T.E. Work stress precipitates depression and anxiety in young, working women and men. Psychol. Med. 2007, 37, 1119–1129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolentino, J.C.; Schmidt, S.L. DSM-5 Criteria and Depression Severity: Implications for Clinical Practice. Front. Psychiatry 2018, 9, 450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, R.C.; Bromet, E.J. The Epidemiology of Depression across Cultures. Annu. Rev. Public Health 2013, 34, 119–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verbeek, E.C.; Bevova, M.R.; Hoogendijk, W.J.G.; Heutink, P. The genetics of MDD—A review of challenges and opportunities. J. Depress. Anxiety 2014, 3, 150. [Google Scholar]
- Bromet, E.; Andrade, L.H.; Hwang, I.; Sampson, N.A.; Alonso, J.; De Girolamo, G.; De Graaf, R.; Demyttenaere, K.; Hu, C.; Iwata, N.; et al. Cross-national epidemiology of DSM-IV major depressive episode. BMC Med. 2011, 9, 90. [Google Scholar] [CrossRef]
- Hirschfeld, R.M.A. The Epidemiology of Depression and the Evolution of Treatment. J. Clin. Psychiatry 2012, 73, 5–9. [Google Scholar] [CrossRef]
- Razmara, A.; Valle, N.; Markovic, D.; Sanossian, N.; Ovbiagele, B.; Dutta, T.; Towfighi, A. Depression Is Associated with a Higher Risk of Death among Stroke Survivors. J. Stroke Cerebrovasc. Dis. 2017, 26, 2870–2879. [Google Scholar] [CrossRef]
- Campayo, A.; Gómez-Biel, C.H.; Lobo, A. Diabetes and depression. Curr. Psychiatry Rep. 2011, 13, 26–30. [Google Scholar] [CrossRef]
- Jiang, X.; Asmaro, R.; O’Sullivan, D.M.; Modi, J.; Budnik, E.; Schnatz, P.F. Depression may be a risk factor for coronary heart disease in midlife women <65 years: A 9-year prospective cohort study. Int. J. Cardiol. 2018, 271, 8–12. [Google Scholar] [CrossRef]
- Gross, A.L.; Gallo, J.J.; Eaton, W.W. Depression and cancer risk: 24 years of follow-up of the Baltimore Epidemiologic Catchment Area sample. Cancer Causes Control. 2009, 21, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Voinov, B.; Richie, W.D.; Bailey, R.K. Depression and chronic diseases: It is time for a synergistic mental health and primary care approach. Prim. Care Companion CNS Disord. 2013, 15, 26226. [Google Scholar] [CrossRef] [PubMed]
- Cummins, N.; Scherer, S.; Krajewski, J.; Schnieder, S.; Epps, J.; Quatieri, T.F. A review of depression and suicide risk assessment using speech analysis. Speech Commun. 2015, 71, 10–49. [Google Scholar] [CrossRef]
- Cuijpers, P.; Vogelzangs, N.; Twisk, J.; Kleiboer, A.; Li, J.; Penninx, B.W. Comprehensive Meta-Analysis of Excess Mortality in Depression in the General Community Versus Patients with Specific Illnesses. Am. J. Psychiatry 2014, 171, 453–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shokrgozar, S.; Khesht-Masjedi, M.F.; Abdollahi, E.; Habibi, B.; Asghari, T.; Ofoghi, R.S.; Pazhooman, S. The relationship between gender, age, anxiety, depression, and academic achievement among teenagers. J. Fam. Med. Prim. Care 2019, 8, 799–804. [Google Scholar] [CrossRef]
- Breslau, J.; Miller, E.; Chung, W.-J.J.; Schweitzer, J.B. Childhood and adolescent onset psychiatric disorders, substance use, and failure to graduate high school on time. J. Psychiatr. Res. 2011, 45, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Ford, E.; Clark, C.; McManus, S.; Harris, J.; Jenkins, R.; Bebbington, P.; Brugha, T.; Meltzer, H.; Stansfeld, S. Common mental disorders, unemployment and welfare benefits in England. Public Health 2010, 124, 675–681. [Google Scholar] [CrossRef]
- Doan, S.N. Allostatic load: Developmental and conceptual considerations in a multi-system physiological indicator of chronic stress exposure. Dev. Psychobiol. 2021, 63, 825–836. [Google Scholar] [CrossRef]
- Guidi, J.; Lucente, M.; Sonino, N.; Fava, G.A. Allostatic Load and Its Impact on Health: A Systematic Review. Psychother. Psychosom. 2020, 90, 11–27. [Google Scholar] [CrossRef]
- McEwen, B.S.; Bowles, N.P.; Gray, J.D.; Hill, M.N.; Hunter, R.G.; Karatsoreos, I.N.; Nasca, C. Mechanisms of stress in the brain. Nat. Neurosci. 2015, 18, 1353–1363. [Google Scholar] [CrossRef]
- Kivimaki, M.; Steptoe, A. Effects of stress on the development and progression of cardiovascular disease. Nat. Rev. Cardiol. 2017, 15, 215–229. [Google Scholar] [CrossRef]
- Dai, S.; Mo, Y.; Wang, Y.; Xiang, B.; Liao, Q.; Zhou, M.; Li, X.; Li, Y.; Xiong, W.; Li, G.; et al. Chronic Stress Promotes Cancer Development. Front. Oncol. 2020, 10, 1492. [Google Scholar] [CrossRef] [PubMed]
- Willner, P.; Towell, A.; Sampson, D.; Sophokleous, S.; Muscat, R. Reduction of sucrose preference by chronic unpredictable mild stress, and its restoration by a tricyclic antidepressant. Psychopharmacology 1987, 93, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Willner, P. Validity, reliability and utility of the chronic mild stress model of depression: A 10-year review and evaluation. Psychopharmacology 1997, 134, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Willner, P.; Muscat, R.; Papp, M. Chronic mild stress-induced anhedonia: A realistic animal model of depression. Neurosci. Biobehav. Rev. 1992, 16, 525–534. [Google Scholar] [CrossRef]
- Antoniuk, S.; Bijata, M.; Ponimaskin, E.; Wlodarczyk, J. Chronic unpredictable mild stress for modeling depression in rodents: Meta-analysis of model reliability. Neurosci. Biobehav. Rev. 2018, 99, 101–116. [Google Scholar] [CrossRef]
- Guze, S.B. Diagnostic and Statistical Manual of Mental Disorders, 4th ed. (DSM-IV). Am. J. Psychiatry 1995, 152, 1228. [Google Scholar] [CrossRef]
- Listunova, L.; Roth, C.; Bartolovic, M.; Kienzle, J.; Bach, C.; Weisbrod, M.; Roesch-Ely, D. Cognitive Impairment Along the Course of Depression: Non-Pharmacological Treatment Options. Psychopathology 2018, 51, 295–305. [Google Scholar] [CrossRef]
- Roca, M.; Vives, M.; López-Navarro, E.; García-Campayo, J.; Gili, M. Cognitive impairments and depression: A critical review. Actas Esp. Psiquiatr. 2015, 43, 187–193. [Google Scholar]
- Mattingly, G.; Anderson, R.H.; Mattingly, S.G.; Anderson, E.Q. The impact of cognitive challenges in major depression: The role of the primary care physician. Postgrad. Med. 2016, 128, 665–671. [Google Scholar] [CrossRef]
- Fossati, P. Is major depression a cognitive disorder? Rev. Neurol. 2018, 174, 212–215. [Google Scholar] [CrossRef]
- Harvey, P.D. Cognitive impairments in major depression and bipolar disorders. Psychiatry 2007, 4, 12–14. [Google Scholar] [PubMed]
- Gualtieri, C.T.; Johnson, L.G.; Benedict, K.B. Neurocognition in depression: Patients on and off medication versus healthy comparison subjects. J. Neuropsychiatry Clin. Neurosci. 2006, 18, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Reppermund, S.; Zihl, J.; Lucae, S.; Horstmann, S.; Kloiber, S.; Holsboer, F.; Ising, M. Persistent Cognitive Impairment in Depression: The Role of Psychopathology and Altered Hypothalamic-Pituitary-Adrenocortical (HPA) System Regulation. Biol. Psychiatry 2007, 62, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Dolan, R.J. Emotion, cognition, and behavior. Science 2002, 298, 1191–1194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuzis, G.; Sabe, L.; Tiberti, C.; Leiguarda, R.; Starkstein, S.E. Cognitive functions in major depression and Parkinson disease. Arch. Neurol. 1997, 54, 982–986. [Google Scholar] [CrossRef]
- Ravnkilde, B.; Videbech, P.; Clemmensen, K.; Egander, A.; Rasmussen, N.A.; Rosenberg, R. Cognitive deficits in major depression. Scand. J. Psychol. 2002, 43, 239–251. [Google Scholar] [CrossRef]
- Song, L.; Che, W.; Min-Wei, W.; Murakami, Y.; Matsumoto, K. Impairment of the spatial learning and memory induced by learned helplessness and chronic mild stress. Pharmacol. Biochem. Behav. 2006, 83, 186–193. [Google Scholar] [CrossRef]
- Lu, Q.; Mouri, A.; Yang, Y.; Kunisawa, K.; Teshigawara, T.; Hirakawa, M.; Mori, Y.; Yamamoto, Y.; Libo, Z.; Nabeshima, T.; et al. Chronic unpredictable mild stress-induced behavioral changes are coupled with dopaminergic hyperfunction and serotonergic hypofunction in mouse models of depression. Behav. Brain Res. 2019, 372, 112053. [Google Scholar] [CrossRef]
- Yan, T.; He, B.; Wan, S.; Xu, M.; Yang, H.; Xiao, F.; Bi, K.; Jia, Y. Antidepressant-like effects and cognitive enhancement of Schisandra chinensis in chronic unpredictable mild stress mice and its related mechanism. Sci. Rep. 2017, 7, 6903. [Google Scholar] [CrossRef] [Green Version]
- Willner, P. The chronic mild stress procedure as an animal model of depression: Valid, reasonably reliable, and useful. Psychopharmacology 1997, 134, 371–377. [Google Scholar] [CrossRef]
- Alghamdi, B.S. The Effect of Melatonin and Exercise on Social Isolation-Related Behavioral Changes in Aged Rats. Front. Aging Neurosci. 2022, 14, 22. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, B.S.; AboTaleb, H.A. Melatonin improves memory defects in a mouse model of multiple sclerosis by up-regulating cAMP-response element-binding protein and synapse-associated proteins in the prefrontal cortex. J. Integr. Neurosci. 2020, 19, 229–237. [Google Scholar] [PubMed]
- Braun, A.; Skelton, M.; Vorhees, C.V.; Williams, M.T. Comparison of the elevated plus and elevated zero mazes in treated and untreated male Sprague–Dawley rats: Effects of anxiolytic and anxiogenic agents. Pharmacol. Biochem. Behav. 2011, 97, 406–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zu, X.; Zhang, M.; Li, W.; Xie, H.; Lin, Z.; Yang, N.; Liu, X.; Zhang, W. Antidepressant-like Effect of Bacopaside I in Mice Exposed to Chronic Unpredictable Mild Stress by Modulating the Hypothalamic–Pituitary–Adrenal Axis Function and Activating BDNF Signaling Pathway. Neurochem. Res. 2017, 42, 3233–3244. [Google Scholar] [CrossRef]
- Labban, S.; Alghamdi, B.S.; Alshehri, F.S.; Kurdi, M. Effects of melatonin and resveratrol on recognition memory and passive avoidance performance in a mouse model of Alzheimer’s disease. Behav. Brain Res. 2021, 402, 113100. [Google Scholar] [CrossRef]
- Jiang, H.; Ashraf, G.M.; Liu, M.; Zhao, K.; Wang, Y.; Wang, L.; Xing, J.; Alghamdi, B.S.; Li, Z.; Liu, R. Tilianin Ameliorates Cognitive Dysfunction and Neuronal Damage in Rats with Vascular Dementia via p-CaMKII/ERK/CREB and ox-CaMKII-Dependent MAPK/NF-κB Pathways. Oxidative Med. Cell. Longev. 2021, 2021, 6673967. [Google Scholar] [CrossRef]
- Vorhees, C.V.; Williams, M. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [Green Version]
- Pałucha-Poniewiera, A.; Podkowa, K.; Rafało-Ulińska, A.; Brański, P.; Burnat, G. The influence of the duration of chronic unpredictable mild stress on the behavioural responses of C57BL/6J mice. Behav. Pharmacol. 2020, 31, 574–582. [Google Scholar] [CrossRef]
- Hu, C.; Luo, Y.; Wang, H.; Kuang, S.; Liang, G.; Yang, Y.; Mai, S.; Yang, J. Re-evaluation of the interrelationships among the behavioral tests in rats exposed to chronic unpredictable mild stress. PLoS ONE 2017, 12, e0185129. [Google Scholar] [CrossRef] [Green Version]
- Vitale, G.; Ruggieri, V.; Filaferro, M.; Frigeri, C.; Alboni, S.; Tascedda, F.; Brunello, N.; Guerrini, R.; Cifani, C.; Massi, M. Chronic treatment with the selective NOP receptor antagonist [Nphe1,Arg14,Lys15]N/OFQ-NH2 (UFP-101) reverses the behavioural and biochemical effects of unpredictable chronic mild stress in rats. Psychopharmacology 2009, 207, 173–189. [Google Scholar] [CrossRef]
- Strekalova, T. Optimization of the chronic stress depression model in C57 BL/6 mice: Evidences for improved validity. In Behavioral Models in Stress Research; Nova Science Publisher: Hauppauge, NY, USA, 2008. [Google Scholar]
- Slattery, D.A.; Cryan, J. The Ups and Downs of Modelling Mood Disorders in Rodents. ILAR J. 2014, 55, 297–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belovicova, K.; Bogi, E.; Csatlosova, K.; Dubovicky, M. Animal tests for anxiety-like and depression-like behavior in rats. Interdiscip. Toxicol. 2017, 10, 40–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harro, J. Animal models of depression: Pros and cons. Cell Tissue Res. 2018, 377, 5–20. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, E.; Takeshita, S. Effects of illumination and handling upon rat open field activity. Physiol. Behav. 1995, 57, 699–703. [Google Scholar] [CrossRef]
- Ellenbroek, B.; Youn, J. Rodent models in neuroscience research: Is it a rat race? Dis. Models Mech. 2016, 9, 1079–1087. [Google Scholar] [CrossRef] [Green Version]
- Strekalova, T.; Steinbusch, H.W. Measuring behavior in mice with chronic stress depression paradigm. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 348–361. [Google Scholar] [CrossRef]
- Ferreira, J.M.F.; Castanheira, L.; Sebastião, A.M.; Telles-Correia, D. Depression Assessment in Clinical Trials and Pre-clinical Tests: A Critical Review. Curr. Top. Med. Chem. 2018, 18, 1677–1703. [Google Scholar] [CrossRef]
- Maslova, L.; Bulygina, V.; Markel, A. Chronic stress during prepubertal development: Immediate and long-lasting effects on arterial blood pressure and anxiety-related behavior. Psychoneuroendocrinology 2002, 27, 549–561. [Google Scholar] [CrossRef]
- Tannenbaum, B.; Tannenbaum, G.; Sudom, K.; Anisman, H. Neurochemical and behavioral alterations elicited by a chronic intermittent stressor regimen: Implications for allostatic load. Brain Res. 2002, 953, 82–92. [Google Scholar] [CrossRef]
- Willner, P. Chronic Mild Stress (CMS) Revisited: Consistency and Behavioural-Neurobiological Concordance in the Effects of CMS. Neuropsychobiology 2005, 52, 90–110. [Google Scholar] [CrossRef]
- Strekalova, T.; Spanagel, R.; Dolgov, O.; Bartsch, D. Stress-induced hyperlocomotion as a confounding factor in anxiety and depression models in mice. Behav. Pharmacol. 2005, 16, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Mombereau, C.; Vassout, A. The tail suspension test as a model for assessing antidepressant activity: Review of pharmacological and genetic studies in mice. Neurosci. Biobehav. Rev. 2005, 29, 571–625. [Google Scholar] [CrossRef] [PubMed]
- Moretti, M.; Colla, A.; de Oliveira Balen, G.; dos Santos, D.B.; Budni, J.; de Freitas, A.E.; Farina, M.; Rodrigues, A.L.S. Ascorbic acid treatment, similarly to fluoxetine, reverses depressive-like behavior and brain oxidative damage induced by chronic unpredictable stress. J. Psychiatr. Res. 2012, 46, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Niu, C.; Wang, J.; Yang, H.; Du, Y.; Wei, L.; Li, C. The depressive-like behaviors of chronic unpredictable mild stress-treated mice ameliorated by Tibetan medicine Zuotai: Involvement in the hypothalamic–pituitary–adrenal (HPA) axis pathway. Neuropsychiatr. Dis. Treat. 2018, 14, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Nollet, M.; Le Guisquet, A.; Belzung, C. Models of Depression: Unpredictable Chronic Mild Stress in Mice. Curr. Protoc. Pharmacol. 2013, 61, 5–65. [Google Scholar] [CrossRef]
- Neis, V.B.; Bettio, L.E.; Moretti, M.; Rosa, P.B.; Ribeiro, C.M.; Freitas, A.E.; Gonçalves, F.M.; Leal, R.B.; Rodrigues, A.L.S. Acute agmatine administration, similar to ketamine, reverses depressive-like behavior induced by chronic unpredictable stress in mice. Pharmacol. Biochem. Behav. 2016, 150, 108–114. [Google Scholar] [CrossRef]
- Strekalova, T.; Spanagel, R.; Bartsch, D.; Henn, F.; Gass, P. Stress-Induced Anhedonia in Mice is Associated with Deficits in Forced Swimming and Exploration. Neuropsychopharmacology 2004, 29, 2007–2017. [Google Scholar] [CrossRef]
- Forbes, N.F.; Stewart, C.A.; Matthews, K.; Reid, I.C. Chronic Mild Stress and Sucrose Consumption: Validity as a Model of Depression. Physiol. Behav. 1996, 60, 1481–1484. [Google Scholar] [CrossRef]
- Bekris, S.; Antoniou, K.; Daskas, S.; Papadopoulou-Daifoti, Z. Behavioural and neurochemical effects induced by chronic mild stress applied to two different rat strains. Behav. Brain Res. 2005, 161, 45–59. [Google Scholar] [CrossRef]
- Papp, M.; Gruca, P.; Boyer, P.-A.; Mocaër, E. Effect of Agomelatine in the Chronic Mild Stress Model of Depression in the Rat. Neuropsychopharmacology 2002, 28, 694–703. [Google Scholar] [CrossRef]
- Lu, Y.; Ho, C.S.; McIntyre, R.S.; Wang, W.; Ho, R. Effects of vortioxetine and fluoxetine on the level of Brain Derived Neurotrophic Factors (BDNF) in the hippocampus of chronic unpredictable mild stress-induced depressive rats. Brain Res. Bull. 2018, 142, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Sun, R.; Ji, Z.; Li, X.; Fu, Q.; Ma, S. Perilla aldehyde attenuates CUMS-induced depressive-like behaviors via regulating TXNIP/TRX/NLRP3 pathway in rats. Life Sci. 2018, 206, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yuan, S.; Pu, J.; Yang, L.; Zhou, X.; Liu, L.; Jiang, X.; Zhang, H.; Teng, T.; Tian, L.; et al. Integrated Metabolomics and Proteomics Analysis of Hippocampus in a Rat Model of Depression. Neuroscience 2018, 371, 207–220. [Google Scholar] [CrossRef]
- Fan, C.; Song, Q.; Wang, P.; Li, Y.; Yang, M.; Yu, S.Y. Neuroprotective Effects of Ginsenoside-Rg1 Against Depression-Like Behaviors via Suppressing Glial Activation, Synaptic Deficits, and Neuronal Apoptosis in Rats. Front. Immunol. 2018, 9, 2889. [Google Scholar] [CrossRef]
- Kreisel, T.; Frank, M.G.; Licht, T.; Reshef, R.; Ben-Menachem-Zidon, O.; Baratta, M.V.; Maier, S.F.; Yirmiya, R. Dynamic microglial alterations underlie stress-induced depressive-like behavior and suppressed neurogenesis. Mol. Psychiatry 2013, 19, 699–709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, G.; Hui, J.; Zhang, Z.; Liu, S.; Zhang, X.; Teng, G.; Chan, K.C.W.; Wu, E.X.; Nie, B.; Shan, B.; et al. Learning and Memory Alterations Are Associated with Hippocampal N-acetylaspartate in a Rat Model of Depression as Measured by 1H-MRS. PLoS ONE 2011, 6, e28686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Ren, L.; Min, S.; Hao, X.; Chen, H.; Deng, J. Changes in synaptic plasticity are associated with electroconvulsive shock-induced learning and memory impairment in rats with depression-like behavior. Neuropsychiatr. Dis. Treat. 2018, 14, 1737–1746. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Zhang, Q.; Gu, J.; Wang, X.; Xie, K.; Xian, X.; Wang, J.; Jiang, H.; Wang, Z. Resveratrol prevents impaired cognition induced by chronic unpredictable mild stress in rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2014, 49, 21–29. [Google Scholar] [CrossRef]
- Twig, G.; Elorza, A.; Molina, A.J.A.; Mohamed, H.; Wikstrom, J.D.; Walzer, G.; Stiles, L.; Haigh, S.E.; Katz, S.; Las, G.; et al. Fission and selective fusion govern mitochondrial segregation and elimination by autophagy. EMBO J. 2008, 27, 433–446. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Shui, L.; Wang, S.; Song, Z.; Tai, F. Bilobalide alleviates depression-like behavior and cognitive deficit induced by chronic unpredictable mild stress in mice. Behav. Pharmacol. 2016, 27, 596–605. [Google Scholar] [CrossRef]
- Femenía, T.; Magara, S.; DuPont, C.M.; Lindskog, M. Hippocampal-Dependent Antidepressant Action of the H3Receptor Antagonist Clobenpropit in a Rat Model of Depression. Int. J. Neuropsychopharmacol. 2015, 18, pyv032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berlyne, D.E. Novelty and Curiosity as Determinants of Exploratory Behaviour1. Br. J. Psychol. Gen. Sect. 1950, 41, 68–80. [Google Scholar] [CrossRef]
- Aguilar-Valles, A.; Sánchez, E.; De Gortari, P.; Balderas, I.; Ramírez-Amaya, V.; Bermudez-Rattoni, F.; Joseph-Bravo, P. Analysis of the Stress Response in Rats Trained in the Water-Maze: Differential Expression of Corticotropin-Releasing Hormone, CRH-R1, Glucocorticoid Receptors and Brain-Derived Neurotrophic Factor in Limbic Regions. Neuroendocrinology 2005, 82, 306–319. [Google Scholar] [CrossRef]
- Anisman, H.; Hayley, S.; Kelly, O.; Borowski, T.; Merali, Z. Psychogenic, neurogenic, and systemic stressor effects on plasma corticosterone and behavior: Mouse strain-dependent outcomes. Behav. Neurosci. 2001, 115, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.J.; Diamond, D.M. The stressed hippocampus, synaptic plasticity and lost memories. Nat. Rev. Neurosci. 2002, 3, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Leussis, M.; Bolivar, V.J. Habituation in rodents: A review of behavior, neurobiology, and genetics. Neurosci. Biobehav. Rev. 2006, 30, 1045–1064. [Google Scholar] [CrossRef] [PubMed]
- Hurst, J.L.; West, R.S. Taming anxiety in laboratory mice. Nat. Methods 2010, 7, 825–826. [Google Scholar] [CrossRef]
- Dulawa, S.C.; Hen, R. Recent advances in animal models of chronic antidepressant effects: The novelty-induced hypophagia test. Neurosci. Biobehav. Rev. 2005, 29, 771–783. [Google Scholar] [CrossRef]
- Zhu, S.; Shi, R.; Wang, J.; Wang, J.-F.; Li, X.-M. Unpredictable chronic mild stress not chronic restraint stress induces depressive behaviours in mice. NeuroReport 2014, 25, 1151–1155. [Google Scholar] [CrossRef]
- Videbech, P.; Ravnkilde, B. Hippocampal volume and depression: A meta-analysis of MRI studies. Am. J. Psychiatry 2004, 161, 1957–1966. [Google Scholar] [CrossRef]
- Luo, Y.; Cao, Z.; Wang, D.; Wu, L.; Li, Y.; Sun, W.; Zhu, Y. Dynamic study of the hippocampal volume by structural MRI in a rat model of depression. Neurol. Sci. 2014, 35, 1777–1783. [Google Scholar] [CrossRef] [PubMed]
- Grønli, J.; Bramham, C.; Murison, R.; Kanhema, T.; Fiske, E.; Bjorvatn, B.; Ursin, R.; Portas, C.M. Chronic mild stress inhibits BDNF protein expression and CREB activation in the dentate gyrus but not in the hippocampus proper. Pharmacol. Biochem. Behav. 2006, 85, 842–849. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, M.; Banasr, M.; Duric, V.; Fournier, N.M.; Licznerski, P.; Duman, R.S. Antidepressant Effects of Fibroblast Growth Factor-2 in Behavioral and Cellular Models of Depression. Biol. Psychiatry 2012, 72, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Bessa, J.; Ferreira, D.; Melo, I.; Marques, F.; Cerqueira, J.; Palha, J.; Almeida, O.F.; Sousa, N. The mood-improving actions of antidepressants do not depend on neurogenesis but are associated with neuronal remodeling. Mol. Psychiatry 2008, 14, 764–773. [Google Scholar] [CrossRef] [Green Version]
- Jayatissa, M.N.; Henningsen, K.; Nikolajsen, G.; West, M.J.; Wiborg, O. A reduced number of hippocampal granule cells does not associate with an anhedonia-like phenotype in a rat chronic mild stress model of depression. Stress 2009, 13, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Mineur, Y.; Belzung, C.; Crusio, W. Functional implications of decreases in neurogenesis following chronic mild stress in mice. Neuroscience 2007, 150, 251–259. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stressor | Duration | Description | Days of CUMS Experiment | |
|---|---|---|---|---|
| 1 | Tail clamping | 1 min | Tail pinch 1 cm from the distal part of the tail | 1, 8, 13, 23, 27, 32, 37, 47, 51, 55 |
| 2 | Restraining | 4 h | Restrain in an air-permeability tube | 2, 7, 18, 21, 25, 33, 40, 38, 44, 49 |
| 3 | Wet cage | 24 h | A cage with damp bedding with water | 3, 8, 13, 22, 26, 31, 39, 46, 51, 56 |
| 4 | Food or water deprivation | 24 h | No water bottles or food pellets | 4, 7, 14, 20, 25, 32, 42, 45, 49, 55 |
| 5 | Tilted cage (30° degree) | 24 h | Cages are elevated from one side and kept tilted at 30° degree. | 1, 10, 16, 19, 28, 35,39, 43, 50, 56 |
| 6 | Illumination | 12 h | Overnight illumination (lights on overnight) | 2, 9, 18, 24, 27, 34, 38, 48, 52 |
| 7 | Isolation | 24 h | Housing in separate cages (1 mouse/cage) | 6, 11, 15, 21, 30, 36, 40, 44, 53 |
| 8 | Cage shaking | 10 min | Cage shaking (200 rpm) | 3, 9, 15, 19, 26, 31, 37, 43, 52 |
| 9 | Predator sounds | 10 min | Exposure to loud predator sounds | 4, 12, 16, 20, 29, 35, 42, 45, 50 |
| 10 | An empty cage | 24 h | Empty cage with no bedding | 5, 12, 17, 23, 29, 33, 41, 47, 54 |
| 11 | Exposure to an empty water bottle | 1 h | Water bottle is replaced by an empty one | 6, 10, 14, 24, 28, 36, 40, 48, 53 |
| 12 | Swimming in cold water (4 ℃) | 3 min | Forced swimming in cold water (4 ℃) in cylinder. | 5, 11, 17, 22, 30, 34, 41, 46, 54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqurashi, G.K.; Hindi, E.A.; Zayed, M.A.; Abd El-Aziz, G.S.; Alturkistani, H.A.; Ibrahim, R.F.; Al-thepyani, M.A.; Bakhlgi, R.; Alzahrani, N.A.; Ashraf, G.M.; et al. The Impact of Chronic Unpredictable Mild Stress-Induced Depression on Spatial, Recognition and Reference Memory Tasks in Mice: Behavioral and Histological Study. Behav. Sci. 2022, 12, 166. https://doi.org/10.3390/bs12060166
Alqurashi GK, Hindi EA, Zayed MA, Abd El-Aziz GS, Alturkistani HA, Ibrahim RF, Al-thepyani MA, Bakhlgi R, Alzahrani NA, Ashraf GM, et al. The Impact of Chronic Unpredictable Mild Stress-Induced Depression on Spatial, Recognition and Reference Memory Tasks in Mice: Behavioral and Histological Study. Behavioral Sciences. 2022; 12(6):166. https://doi.org/10.3390/bs12060166
Chicago/Turabian StyleAlqurashi, Ghofran Khalid, Emad A. Hindi, Mohamed A. Zayed, Gamal S. Abd El-Aziz, Hani A. Alturkistani, Rabee F. Ibrahim, Mona Ali Al-thepyani, Refal Bakhlgi, Noor A. Alzahrani, Ghulam Md Ashraf, and et al. 2022. "The Impact of Chronic Unpredictable Mild Stress-Induced Depression on Spatial, Recognition and Reference Memory Tasks in Mice: Behavioral and Histological Study" Behavioral Sciences 12, no. 6: 166. https://doi.org/10.3390/bs12060166
APA StyleAlqurashi, G. K., Hindi, E. A., Zayed, M. A., Abd El-Aziz, G. S., Alturkistani, H. A., Ibrahim, R. F., Al-thepyani, M. A., Bakhlgi, R., Alzahrani, N. A., Ashraf, G. M., & Alghamdi, B. S. (2022). The Impact of Chronic Unpredictable Mild Stress-Induced Depression on Spatial, Recognition and Reference Memory Tasks in Mice: Behavioral and Histological Study. Behavioral Sciences, 12(6), 166. https://doi.org/10.3390/bs12060166