Strangers, Friends, and Lovers Show Different Physiological Synchrony in Different Emotional States

 , , , , , , ,
, , , , , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Procedure
2.3. Stimuli
- A scene from the movie “When Harry met Sally” was used to elicit the emotion of embarrassment (EMBARRASS);
- A scene from the movie “Titanic” was used to elicit the emotion of sadness (SAD);
- A scene from the TV series “The Walking Dead” was used to elicit the emotion of fear (FEAR);
- A scene of a beach with a relaxing music playing in the background was used to induce calmness (CALMNESS);
- A scene from the movie “Notting Hill” was used to elicit romantic love (ROMANCE);
- A scene from the penalty-kick session in the 2006 FIFA World Cup Finals which led to the win of the Italian football team. Since all the participants were of Italian nationality, this stimulus was used to elicit the emotion of pride (PRIDE).
2.4. Physiological Measures
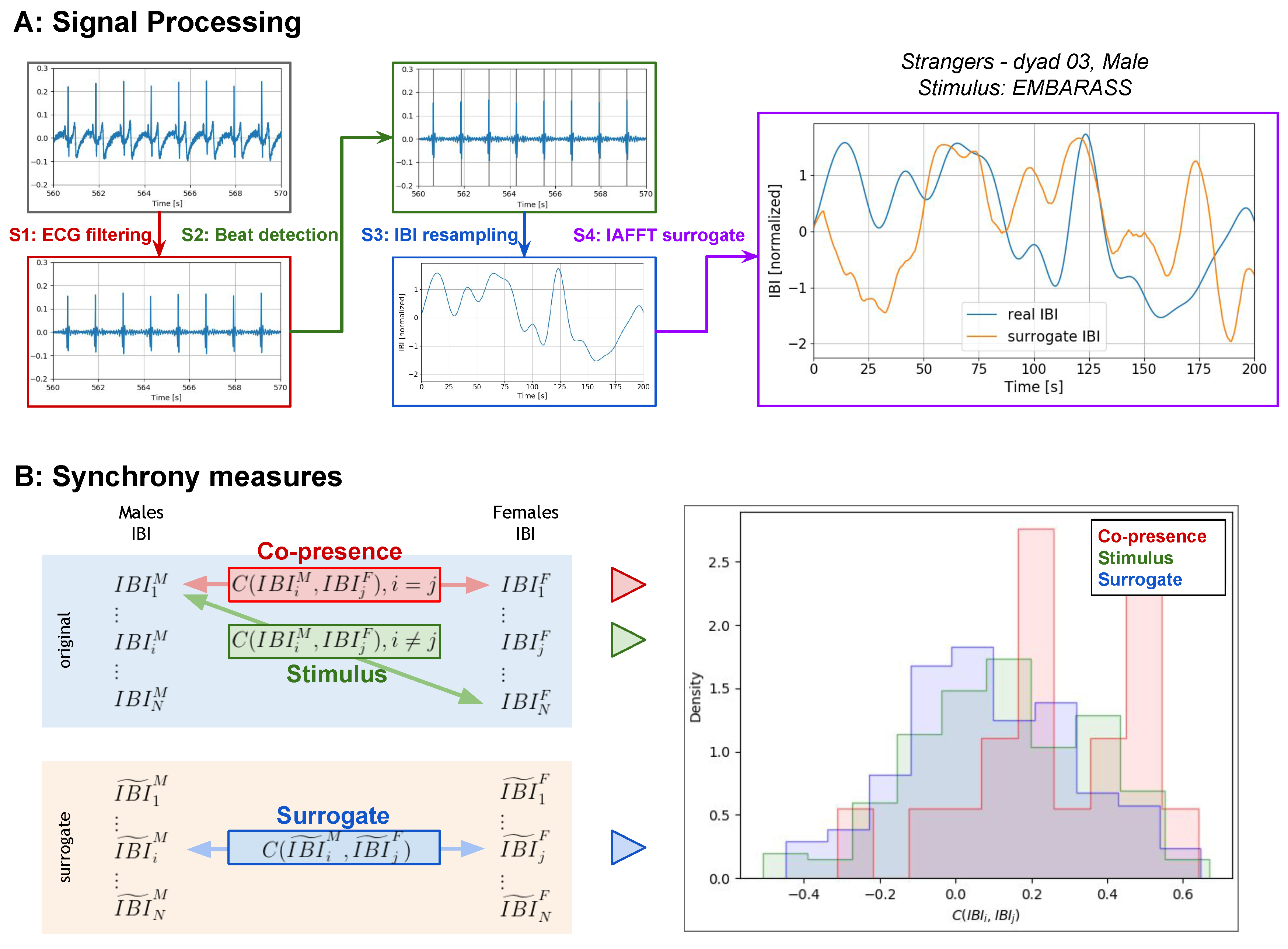
2.5. Synchrony Measures
- Copresence synchrony between the real IBI series of the male and female of the dyad, who watched the videos together: This synchrony measure is influenced by both stimulus and copresence, i.e., the effect of watching the stimulus with the other member of the dyad. The copresence synchrony is computed between all the dyads of each group.
- Stimulus synchrony between the real IBI series of a male and a female belonging to different dyads who did not watch the videos together: This synchrony measure is therefore only influenced by the stimulus, as the male and the female did not watch the stimulus together. The stimulus synchrony is computed between all possible male–female pairs of each group, excluding the real dyad.
- Surrogate synchrony between surrogate signals of a male and a female: This synchrony measure is not influenced by the stimulus or by the copresence and is used to compose the distribution of the null hypothesis that there is no effect of synchrony due to stimulus or copresence. The surrogate synchrony is computed between all possible male–female pairs of each group.
2.6. Analysis Plan
- To replicate and extend the results of Golland and colleagues [9] by investigating the effects of copresence between dyads of strangers with different elicited emotions;
- To investigate the association between type relationship and physiological synchrony;
- To investigate the influence of emotional state on physiological synchrony across different relationship types.
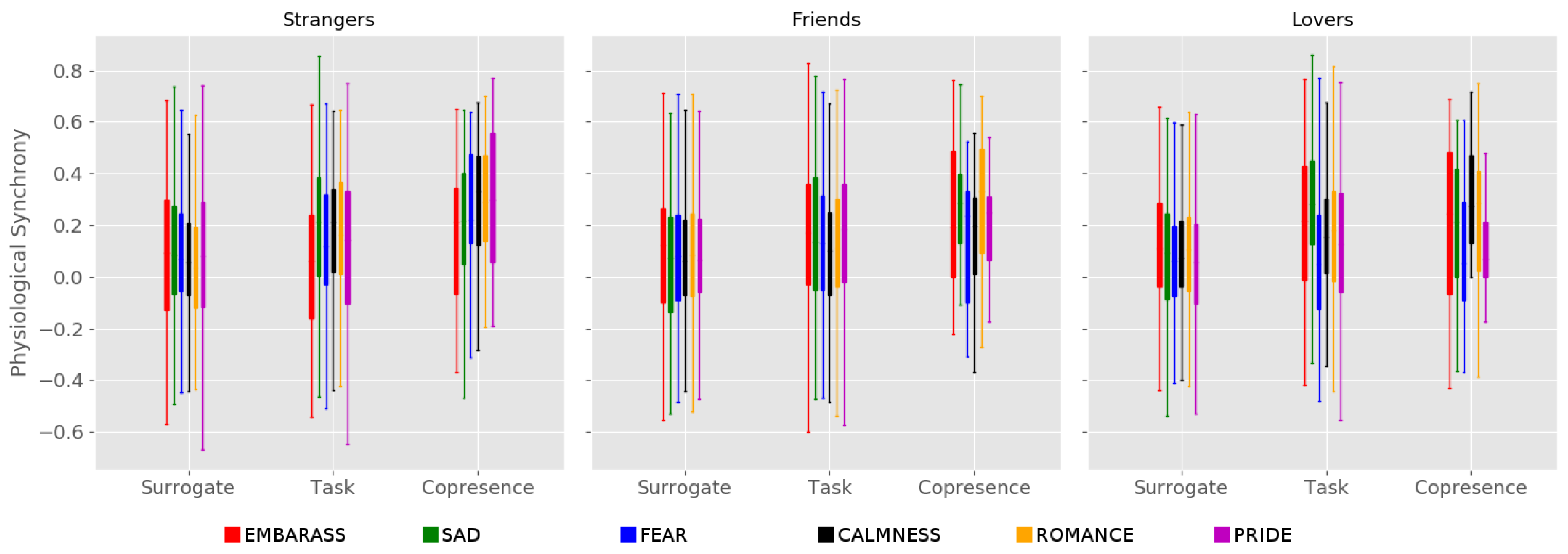
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Leclère, C.; Viaux, S.; Avril, M.; Achard, C.; Chetouani, M.; Missonnier, S.; Cohen, D. Why synchrony matters during mother-child interactions: A systematic review. PLoS ONE 2014, 9, e113571. [Google Scholar] [CrossRef] [PubMed]
- Hatfield, E.; Cacioppo, J.T.; Rapson, R.L. Emotional contagion. Curr. Dir. Psychol. Sci. 1993, 2, 96–100. [Google Scholar] [CrossRef]
- Barsade, S.G. The ripple effect: Emotional contagion and its influence on group behavior. Adm. Sci. Q. 2002, 47, 644–675. [Google Scholar] [CrossRef]
- Park, S.; Choi, S.J.; Mun, S.; Whang, M. Measurement of emotional contagion using synchronization of heart rhythm pattern between two persons: Application to sales managers and sales force synchronization. Physiol. Behav. 2019, 200, 148–158. [Google Scholar] [CrossRef]
- Chikersal, P.; Tomprou, M.; Kim, Y.J.; Woolley, A.W.; Dabbish, L. Deep structures of collaboration: Physiological correlates of collective intelligence and group satisfaction. In Proceedings of the 2017 ACM Conference on Computer Supported Cooperative Work and Social Computing, Portland, OR, USA, 25 February–1 March 2017; ACM: New York, NY, USA, 2017; pp. 873–888. [Google Scholar]
- Kelsey, R.M. Beta-adrenergic cardiovascular reactivity and adaptation to stress: The cardiac pre-ejection period as an index of effort. In How motivation Affects Cardiovascular Response: Mechanisms and Applications; Wright, R.A., Gendolla, G.H.E., Eds.; American Psychological Association: Worcester, MA, USA, 2012; pp. 43–60. [Google Scholar]
- Cacioppo, J.T.; Sandman, C.A.; Walker, B.B. The effects of operant heart rate conditioning on cognitive elaboration and attitude change. Psychophysiology 1978, 15, 330–338. [Google Scholar] [CrossRef]
- Suess, P.E.; Porges, S.W.; Plude, D.J. Cardiac vagal tone and sustained attention in school-age children. Psychophysiology 1994, 31, 17–22. [Google Scholar] [CrossRef]
- Golland, Y.; Arzouan, Y.; Levit-Binnun, N. The mere copresence: Synchronization of autonomic signals and emotional responses across co-present individuals not engaged in direct interaction. PLoS ONE 2015, 10, e0125804. [Google Scholar] [CrossRef]
- Danyluck, C.; Page-Gould, E. Social and physiological context can affect the meaning of physiological synchrony. Sci. Rep. 2019, 9, 8222. [Google Scholar] [CrossRef]
- Feldman, R. Bio-behavioral synchrony: A model for integrating biological and microsocial behavioral processes in the study of parenting. Parenting 2012, 12, 154–164. [Google Scholar] [CrossRef]
- Helm, J.L.; Sbarra, D.; Ferrer, E. Assessing cross-partner associations in physiological responses via coupled oscillator models. Emotion 2012, 12, 748. [Google Scholar] [CrossRef]
- Schneiderman, I.; Kanat-Maymon, Y.; Zagoory-Sharon, O.; Feldman, R. Mutual influences between partners’ hormones shape conflict dialog and relationship duration at the initiation of romantic love. Soc. Neurosci. 2014, 9, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Papp, L.M.; Pendry, P.; Simon, C.D.; Adam, E.K. Spouses’ cortisol associations and moderators: Testing physiological synchrony and connectedness in everyday life. Fam. Process 2013, 52, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Cirelli, L.K. How interpersonal synchrony facilitates early prosocial behavior. Curr. Opin. Psychol. 2018, 20, 35–39. [Google Scholar] [CrossRef] [PubMed]
- Leong, V.; Byrne, E.; Clackson, K.; Georgieva, S.; Lam, S.; Wass, S. Speaker gaze increases information coupling between infant and adult brains. Proc. Natl. Acad. Sci. USA 2017, 114, 13290–13295. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R. Parent–infant synchrony: A biobehavioral model of mutual influences in the formation of affiliative bonds. Monogr. Soc. Res. Child Dev. 2012, 77, 42–51. [Google Scholar] [CrossRef]
- Ostlund, B.D.; Measelle, J.R.; Laurent, H.K.; Conradt, E.; Ablow, J.C. Shaping emotion regulation: Attunement, symptomatology, and stress recovery within mother–infant dyads. Dev. Psychobiol. 2017, 59, 15–25. [Google Scholar] [CrossRef]
- Wilson, S.J.; Bailey, B.E.; Jaremka, L.M.; Fagundes, C.P.; Andridge, R.; Malarkey, W.B.; Gates, K.M.; Kiecolt-Glaser, J.K. When couples’ hearts beat together: Synchrony in heart rate variability during conflict predicts heightened inflammation throughout the day. Psychoneuroendocrinology 2018, 93, 107–116. [Google Scholar] [CrossRef]
- Levenson, R.W.; Gottman, J.M. Marital interaction: Physiological linkage and affective exchange. J. Personal. Soc. Psychol. 1983, 45, 587–597. [Google Scholar] [CrossRef]
- Coutinho, J.; Oliveira-Silva, P.; Fernandes, E.; Gonçalves, O.F.; Correia, D.; Perrone Mc-Govern, K.; Tschacher, W. Psychophysiological synchrony during verbal interaction in romantic relationships. Family Process 2018, 58, 716–733. [Google Scholar] [CrossRef]
- Azhari, A.; Leck, W.; Gabrieli, G.; Bizzego, A.; Rigo, P.; Setoh, P.; Bornstein, M.H.; Esposito, G. Parenting stress undermines mother-child brain-to-brain synchrony: A hyperscanning study. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Kaplan, H.B.; Burch, N.R.; Bloom, S.W.; Edelberg, R. Affective orientation and physiological activity (GSR) in small peer groups. Psychosom. Med. 1963, 25, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Gallotti, M.; Frith, C.D. Social cognition in the we-mode. Trends Cogn. Sci. 2013, 17, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Atmaca, S.; Sebanz, N.; Prinz, W.; Knoblich, G. Action co-representation: The joint SNARC effect. Soc. Neurosci. 2008, 3, 410–420. [Google Scholar] [CrossRef] [PubMed]
- Sebanz, N.; Knoblich, G.; Prinz, W. Representing others’ actions: Just like one’s own? Cognition 2003, 88, B11–B21. [Google Scholar] [CrossRef]
- Malik, M.; Bigger, J.T.; Camm, A.J.; Kleiger, R.E.; Malliani, A.; Moss, A.J.; Schwartz, P.J. Heart rate variability: Standards of measurement, physiological interpretation, and clinical use. Eur. Heart J. 1996, 17, 354–381. [Google Scholar] [CrossRef]
- Schreiber, T.; Schmitz, A. Surrogate time series. Phys. D Nonlinear Phenom. 2000, 142, 346–382. [Google Scholar] [CrossRef]
- Golland, Y.; Keissar, K.; Levit-Binnun, N. Studying the dynamics of autonomic activity during emotional experience. Psychophysiology 2014, 51, 1101–1111. [Google Scholar] [CrossRef]
- Mauss, I.B.; Levenson, R.W.; McCarter, L.; Wilhelm, F.H.; Gross, J.J. The tie that binds? Coherence among emotion experience, behavior, and physiology. Emotion 2005, 5, 175. [Google Scholar] [CrossRef]
- Kettunen, J.; Ravaja, N.; Keltikangas-Järvinen, L. Smoothing facilitates the detection of coupled responses in psychophysiological time series. J. Psychophysiol. 2000, 14, 1–10. [Google Scholar] [CrossRef]
- Kettunen, J.; Ravaja, N. A comparison of different time series techniques to analyze phasic coupling: A case study of cardiac and electrodermal activity. Psychophysiology 2000, 37, 395–408. [Google Scholar] [CrossRef]
- Bizzego, A.; Battisti, A.; Gabrieli, G.; Esposito, G.; Furlanello, C. pyphysio: A physiological signal processing library for data science approaches in physiology. SoftwareX 2019, 10, 100287. [Google Scholar] [CrossRef]
- Bizzego, A.; Mina, M.; Zarbo, C.; Esposito, G.; Furlanello, C. Physiolyze: A galaxy-based web service for Heart Rate Variability analysis with online processing. In Proceedings of the 2014 8th Conference of the European Study Group on Cardiovascular Oscillations (ESGCO), Trento, Italy, 25–28 May 2014; pp. 97–98. [Google Scholar]
- Bizzego, A.; Gabrieli, G.; Azhari, A.; Setoh, P.; Esposito, G. Computational methods for the assessment of empathic synchrony. In Neural Approaches to Dynamics of Signal Exchanges; Esposito, A., Faundez-Zanuy, M., Morabito, F., Pasero, E., Eds.; Smart Innovation Systems and Technologies; Springer: Berlin, Germany, 2019. [Google Scholar]
- Gabrieli, G.; Azhari, A.; Esposito, G. PySiology: A python package for physiological feature extraction. In Neural Approaches to Dynamics of Signal Exchanges; Springer: Berlin, Germany, 2020; pp. 395–402. [Google Scholar]
- Deng, Y.; Chang, L.; Yang, M.; Huo, M.; Zhou, R. Gender differences in emotional response: Inconsistency between experience and expressivity. PLoS ONE 2016, 11, e0158666. [Google Scholar] [CrossRef] [PubMed]
- Butler, E.A. Temporal interpersonal emotion systems: The “TIES” that form relationships. Personal. Soc. Psychol. Rev. 2011, 15, 367–393. [Google Scholar] [CrossRef] [PubMed]
- Reddish, P.; Fischer, R.; Bulbulia, J. Let’s dance together: Synchrony, shared intentionality and cooperation. PLoS ONE 2013, 8, e71182. [Google Scholar] [CrossRef] [PubMed]
- Mobbs, D.; Hagan, C.C.; Dalgleish, T.; Silston, B.; Prévost, C. The ecology of human fear: Survival optimization and the nervous system. Front. Neurosci. 2015, 9, 55. [Google Scholar] [CrossRef] [PubMed]
- Ozturk, Y.; Bizzego, A.; Esposito, G.; Furlanello, C.; Venuti, P. Physiological and self-report responses of parents of children with autism spectrum disorder to children crying. Res. Dev. Disabil. 2018, 73, 31–39. [Google Scholar] [CrossRef]
- Shilling, C.; Mellor, P.A. Durkheim, morality and modernity: Collective effervescence, homo duplex and the sources of moral action. Br. J. Sociol. 1998, 193–209. [Google Scholar] [CrossRef]
- Jong, J.; Whitehouse, H.; Kavanagh, C.; Lane, J. Shared negative experiences lead to identity fusion via personal reflection. PLoS ONE 2015, 10, e0145611. [Google Scholar] [CrossRef]
- Whitehouse, H.; Lanman, J.A. The ties that bind us: Ritual, fusion, and identification. Curr. Anthropol. 2014, 55, 674–695. [Google Scholar] [CrossRef]
- Whitehouse, H.; McQuinn, B.; Buhrmester, M.; Swann, W.B. Brothers in Arms: Libyan revolutionaries bond like family. Proc. Natl. Acad. Sci. USA 2014, 111, 17783–17785. [Google Scholar] [CrossRef]
- Jackson, J.C.; Jong, J.; Bilkey, D.; Whitehouse, H.; Zollmann, S.; McNaughton, C.; Halberstadt, J. Synchrony and physiological arousal increase cohesion and cooperation in large naturalistic groups. Sci. Rep. 2018, 8, 127. [Google Scholar] [CrossRef] [PubMed]
- Timmons, A.C.; Margolin, G.; Saxbe, D.E. Physiological linkage in couples and its implications for individual and interpersonal functioning: A literature review. J. Fam. Psychol. 2015, 29, 720. [Google Scholar] [CrossRef] [PubMed]
- Diamond, L.M.; Hicks, A.M.; Otter-Henderson, K.D. Every time you go away: Changes in affect, behavior, and physiology associated with travel-related separations from romantic partners. J. Personal. Soc. Psychol. 2008, 95, 385. [Google Scholar] [CrossRef] [PubMed]
- Coan, J.A.; Schaefer, H.S.; Davidson, R.J. Lending a hand: Social regulation of the neural response to threat. Psychol. Sci. 2006, 17, 1032–1039. [Google Scholar] [CrossRef]
- Buss, D.M.; Schmitt, D.P. Sexual strategies theory: An evolutionary perspective on human mating. Psychol. Rev. 1993, 100, 204. [Google Scholar] [CrossRef]
- Bleske, A.L.; Buss, D.M. Can men and women be just friends? Pers. Relationsh. 2000, 7, 131–151. [Google Scholar] [CrossRef]
- Kacerguis, M.A.; Adams, G.R. Erikson stage resolution: The relationship between identity and intimacy. J. Youth Adolesc. 1980, 9, 117–126. [Google Scholar] [CrossRef]
- Levenson, R.W.; Gottman, J.M. Physiological and affective predictors of change in relationship satisfaction. J. Personal. Soc. Psychol. 1985, 49, 85. [Google Scholar] [CrossRef]
- Lazaridès, A.; Bélanger, C.; Sabourin, S. Personality as moderator of the relationship between communication and couple stability. Eur. J. Psychol. 2010, 6, 11–31. [Google Scholar] [CrossRef]
- Guerrero, L.K. Nonverbal involvement across interactions with same-sex friends, opposite-sex friends and romantic partners: Consistency or change? J. Soc. Pers. Relationsh. 1997, 14, 31–58. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Emotion | Relationship | Surrogate vs Stimulus | Stimulus vs Copresence | ||
|---|---|---|---|---|---|
| U | p | U | p | ||
| EMBARRASS | Strangers | 17341 | p = 0.866 | 1313 | p = 0.086 |
| Friends | 30042 | = 0.003 | 2576 | p = 0.182 | |
| Lovers | 15920 | < 0.001 | 1892 | p = 0.488 | |
| SAD | Strangers | 12707 | < 0.001 | 1571 | p = 0.408 |
| Friends | 27963 | < 0.001 | 2356 | p = 0.066 | |
| Lovers | 10629 | < 0.001 | 2274 | p = 0.926 | |
| FEAR | Strangers | 14581 | p = 0.046 | 1083 | = 0.009 |
| Friends | 31537 | p = 0.027 | 2663 | p = 0.251 | |
| Lovers | 20119 | p = 0.558 | 1869 | p = 0.453 | |
| CALMNESS | Strangers | 11565 | < 0.001 | 1215 | p = 0.036 |
| Friends | 32648 | p = 0.098 | 2489 | p = 0.126 | |
| Lovers | 16037 | < 0.001 | 1308 | = 0.011 | |
| ROMANCE | Strangers | 11285 | < 0.001 | 1233 | p = 0.043 |
| Friends | 30897 | = 0.011 | 1972 | = 0.005 | |
| Lovers | 16660 | = 0.002 | 1584 | p = 0.111 | |
| PRIDE | Strangers | 14944 | p = 0.094 | 1064 | = 0.007 |
| Friends | 27676 | < 0.001 | 2760 | p = 0.342 | |
| Lovers | 17289 | = 0.011 | 2054 | p = 0.725 | |
| Emot. | Strangers vs Friends | Friends vs Lovers | Strangers vs Lovers | |||
|---|---|---|---|---|---|---|
| U | p | U | p | U | p | |
| EMBARRASS | 187 | p = 0.217 | 218 | p = 0.390 | 170 | p = 0.292 |
| SAD | 190 | p = 0.240 | 195 | p = 0.200 | 183 | p = 0.428 |
| FEAR | 176 | p = 0.144 | 190 | p = 0.168 | 120 | p = 0.025 |
| CALMNESS | 149 | p = 0.041 | 160 | p = 0.045 | 188 | p = 0.483 |
| ROMANCE | 214 | p = 0.460 | 204 | p = 0.267 | 174 | p = 0.332 |
| PRIDE | 169 | p = 0.108 | 170 | p = 0.074 | 116 | p = 0.019 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bizzego, A.; Azhari, A.; Campostrini, N.; Truzzi, A.; Ng, L.Y.; Gabrieli, G.; Bornstein, M.H.; Setoh, P.; Esposito, G. Strangers, Friends, and Lovers Show Different Physiological Synchrony in Different Emotional States. Behav. Sci. 2020, 10, 11. https://doi.org/10.3390/bs10010011
Bizzego A, Azhari A, Campostrini N, Truzzi A, Ng LY, Gabrieli G, Bornstein MH, Setoh P, Esposito G. Strangers, Friends, and Lovers Show Different Physiological Synchrony in Different Emotional States. Behavioral Sciences. 2020; 10(1):11. https://doi.org/10.3390/bs10010011
Chicago/Turabian StyleBizzego, Andrea, Atiqah Azhari, Nicola Campostrini, Anna Truzzi, Li Ying Ng, Giulio Gabrieli, Marc H. Bornstein, Peipei Setoh, and Gianluca Esposito. 2020. "Strangers, Friends, and Lovers Show Different Physiological Synchrony in Different Emotional States" Behavioral Sciences 10, no. 1: 11. https://doi.org/10.3390/bs10010011
APA StyleBizzego, A., Azhari, A., Campostrini, N., Truzzi, A., Ng, L. Y., Gabrieli, G., Bornstein, M. H., Setoh, P., & Esposito, G. (2020). Strangers, Friends, and Lovers Show Different Physiological Synchrony in Different Emotional States. Behavioral Sciences, 10(1), 11. https://doi.org/10.3390/bs10010011