Atmospheric Control of Deep Chlorophyll Maximum Development

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
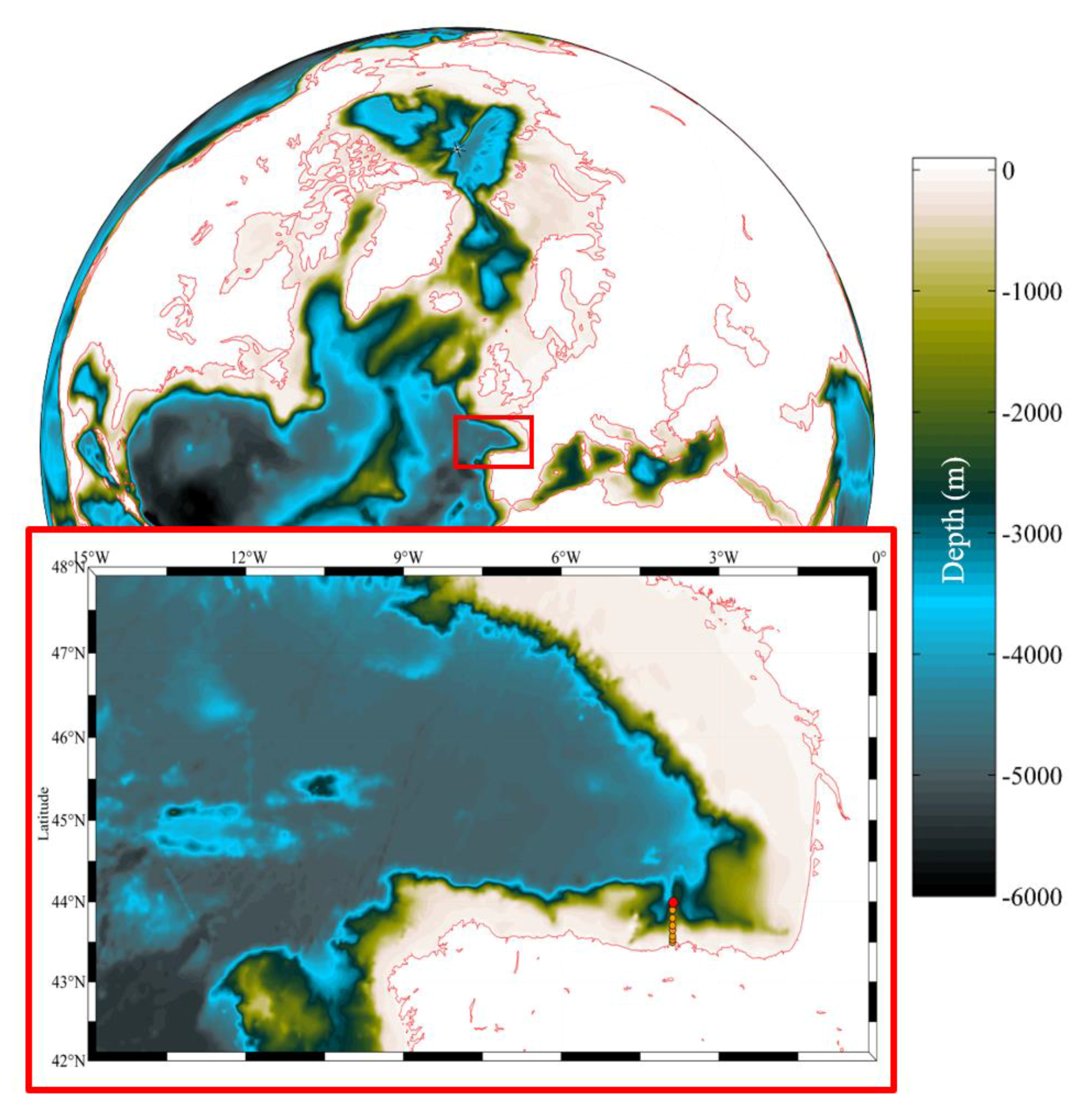
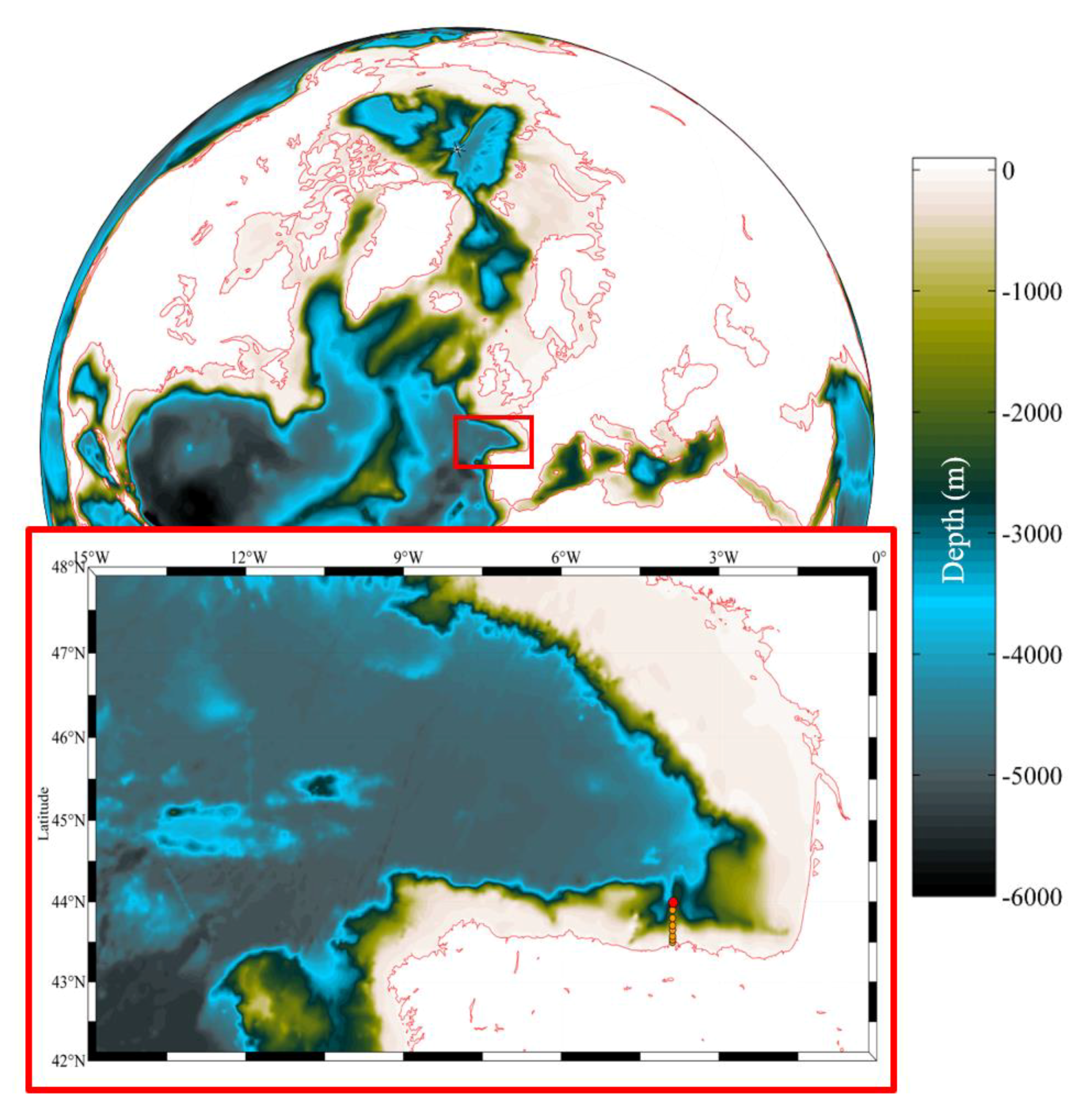
2.1. Study Area and Data Sources
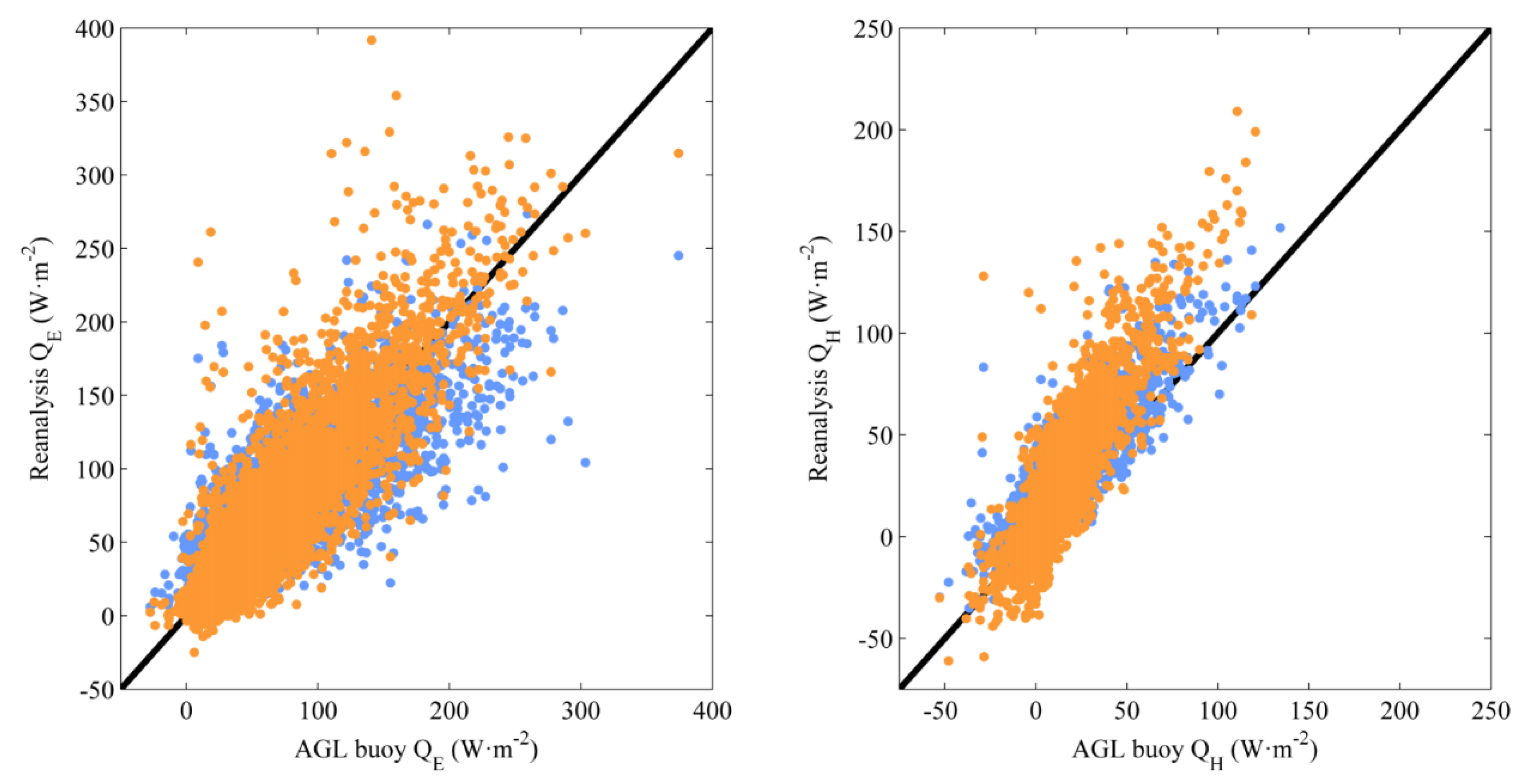
2.2. Changes in Air-Sea Exchanges, Near-Surface Chlorophyll, Depth of DCM, and Mixed-Layer Depth
3. Results and Discussion
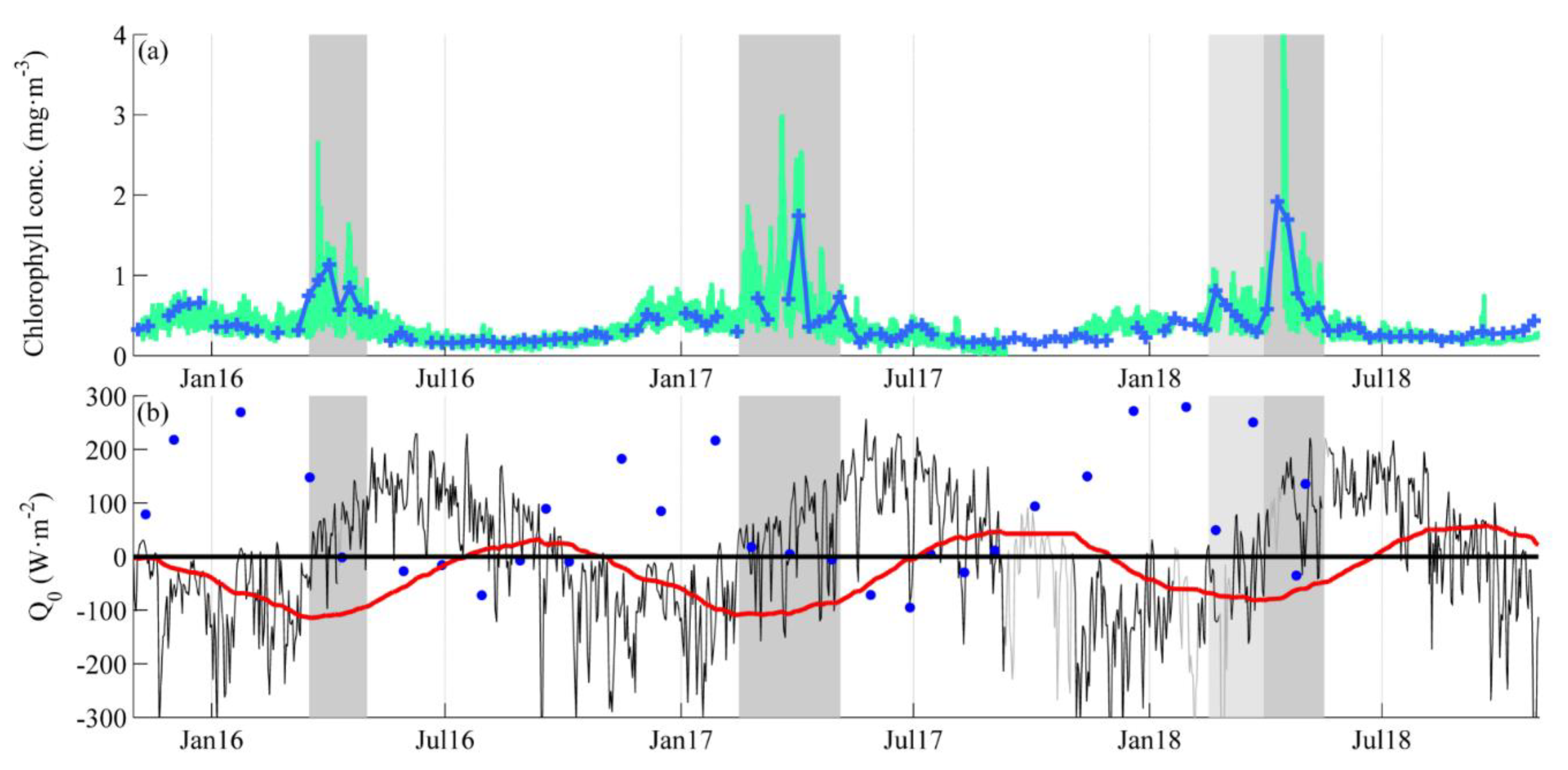
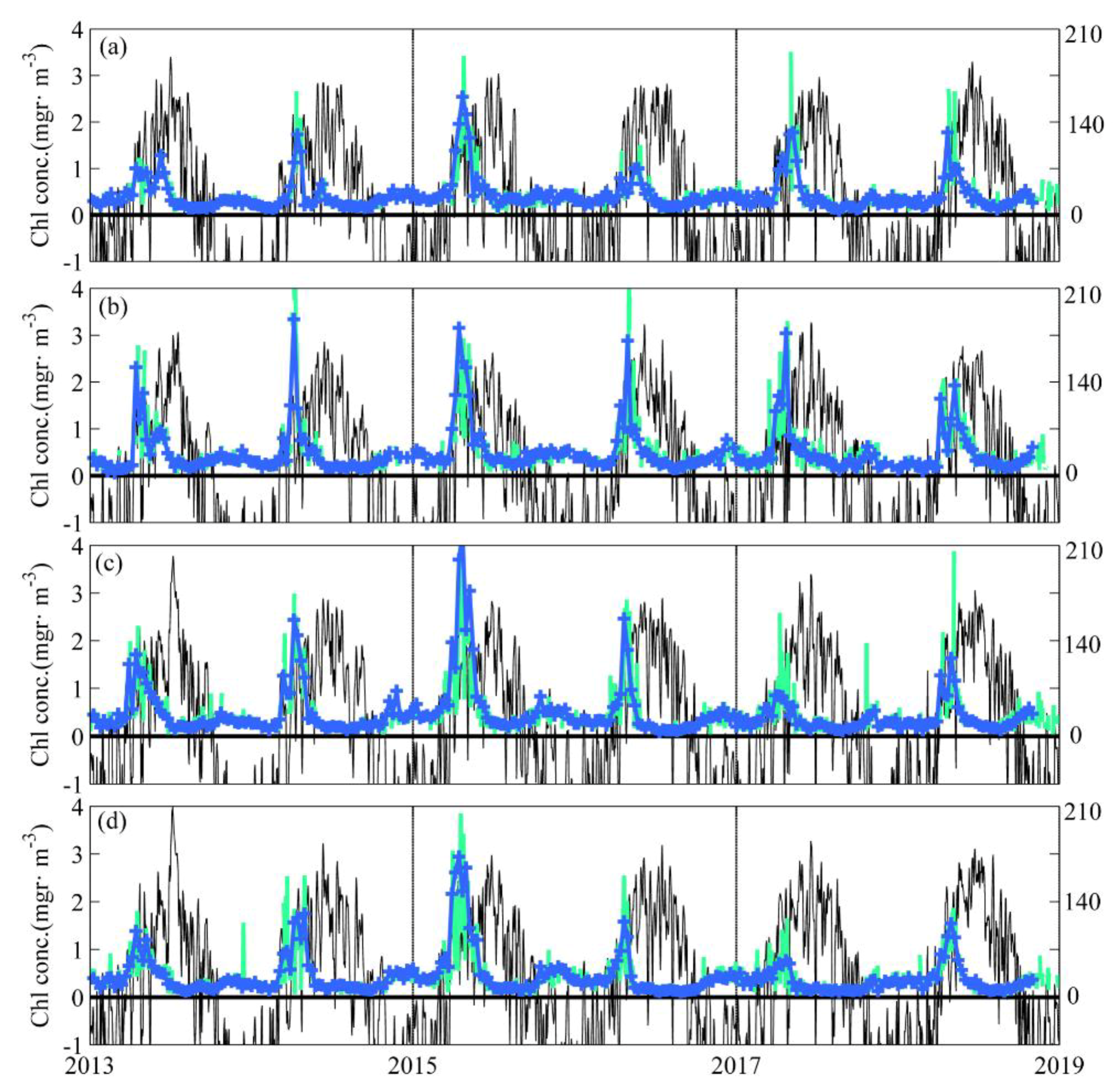
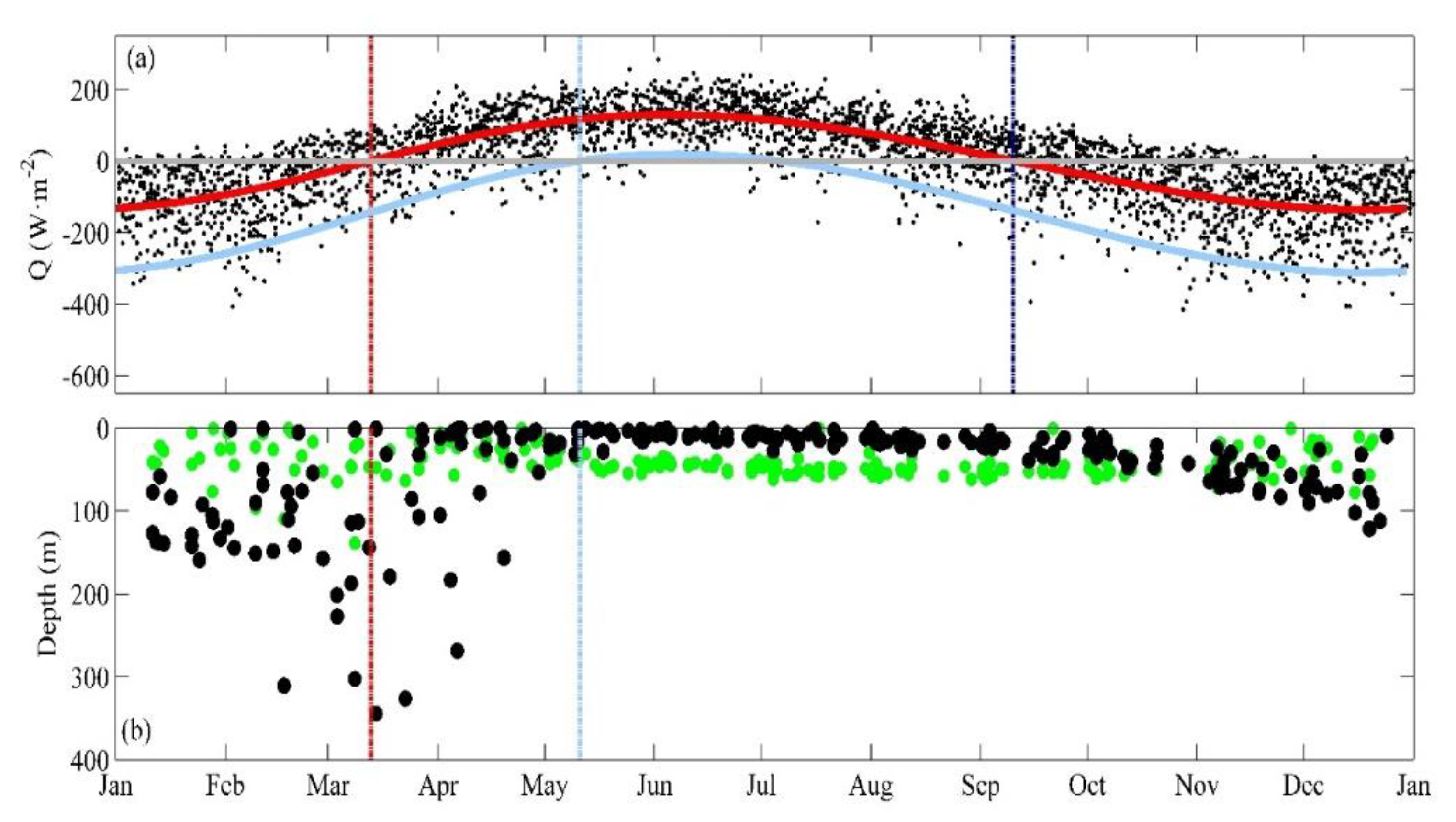
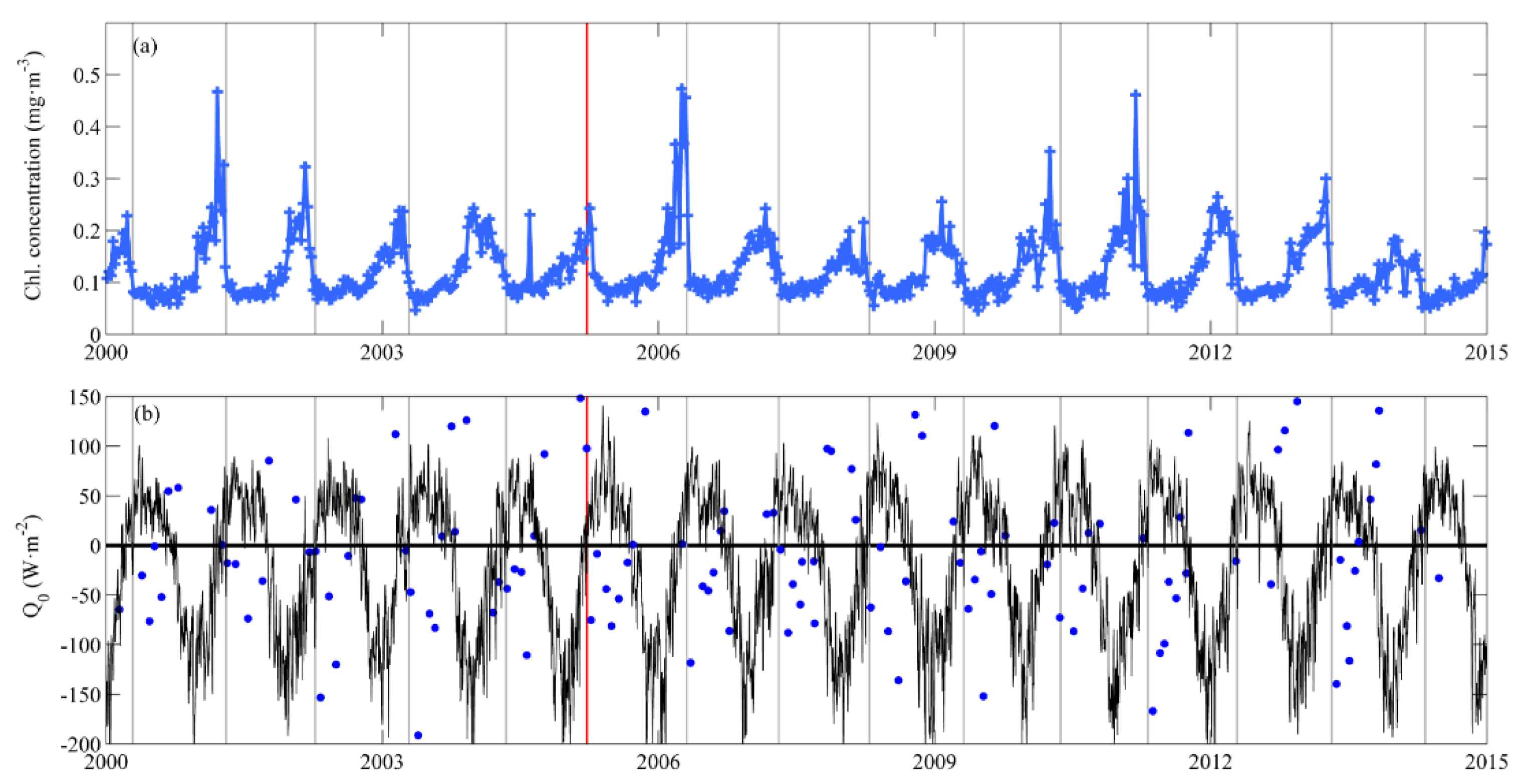
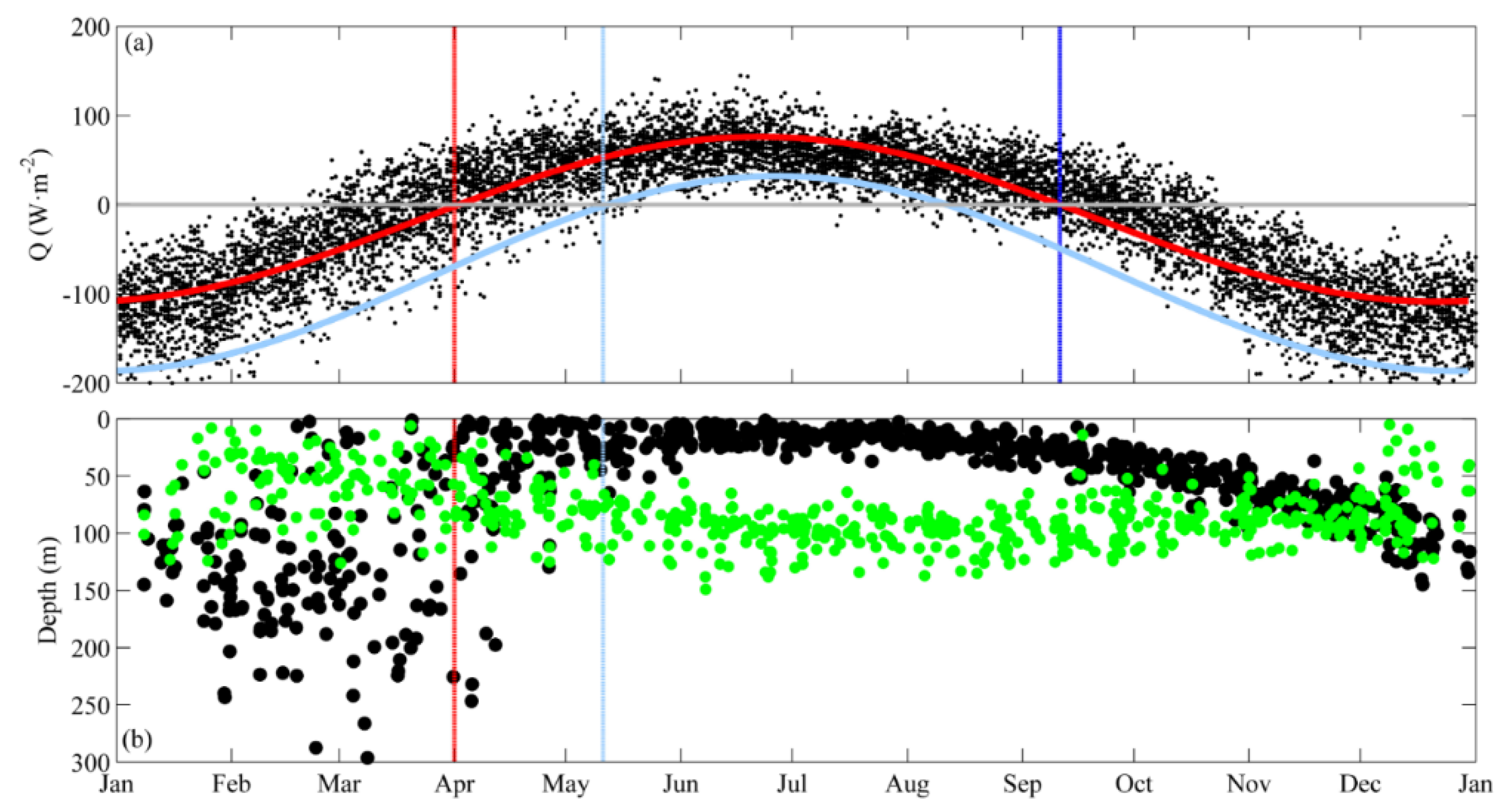
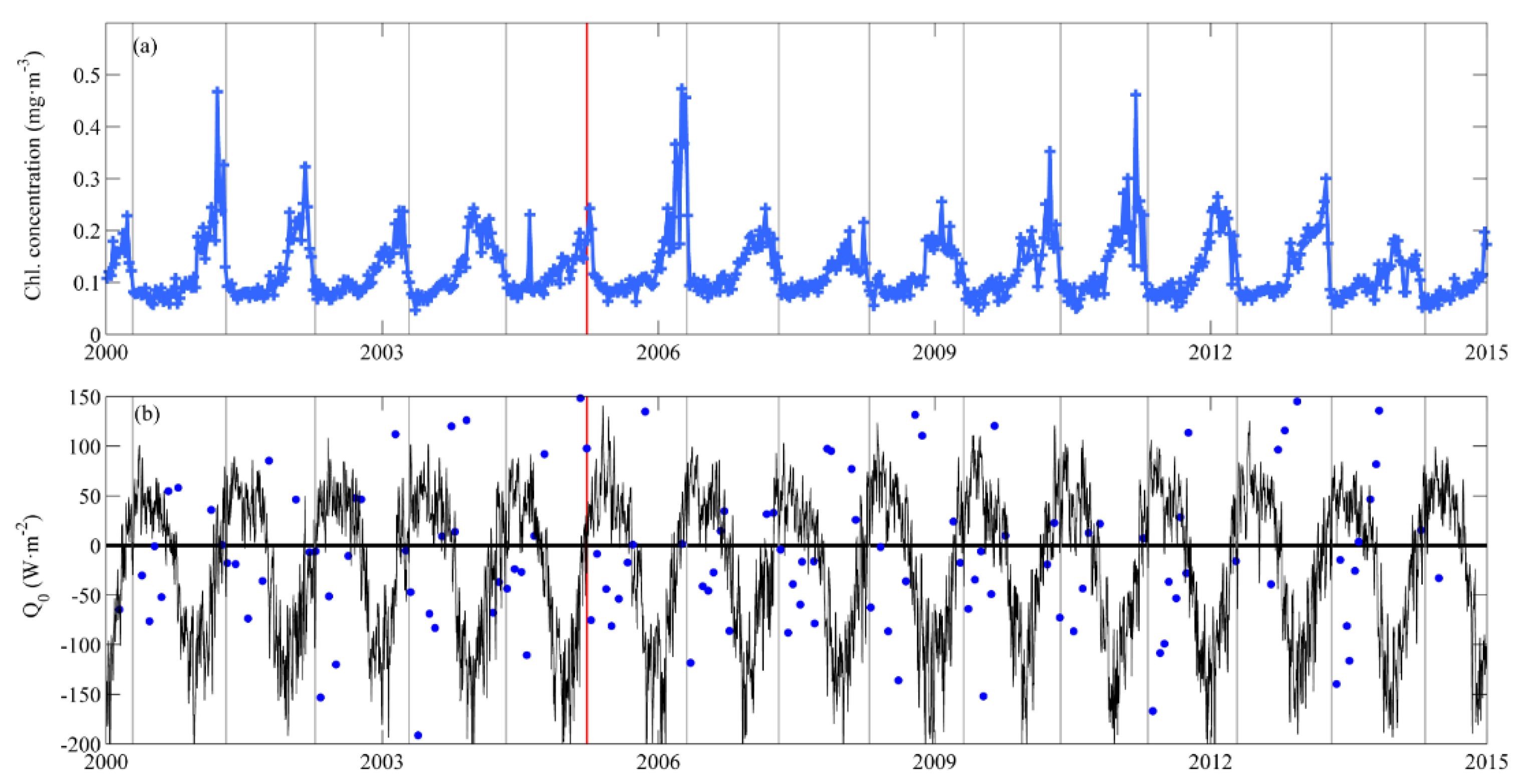
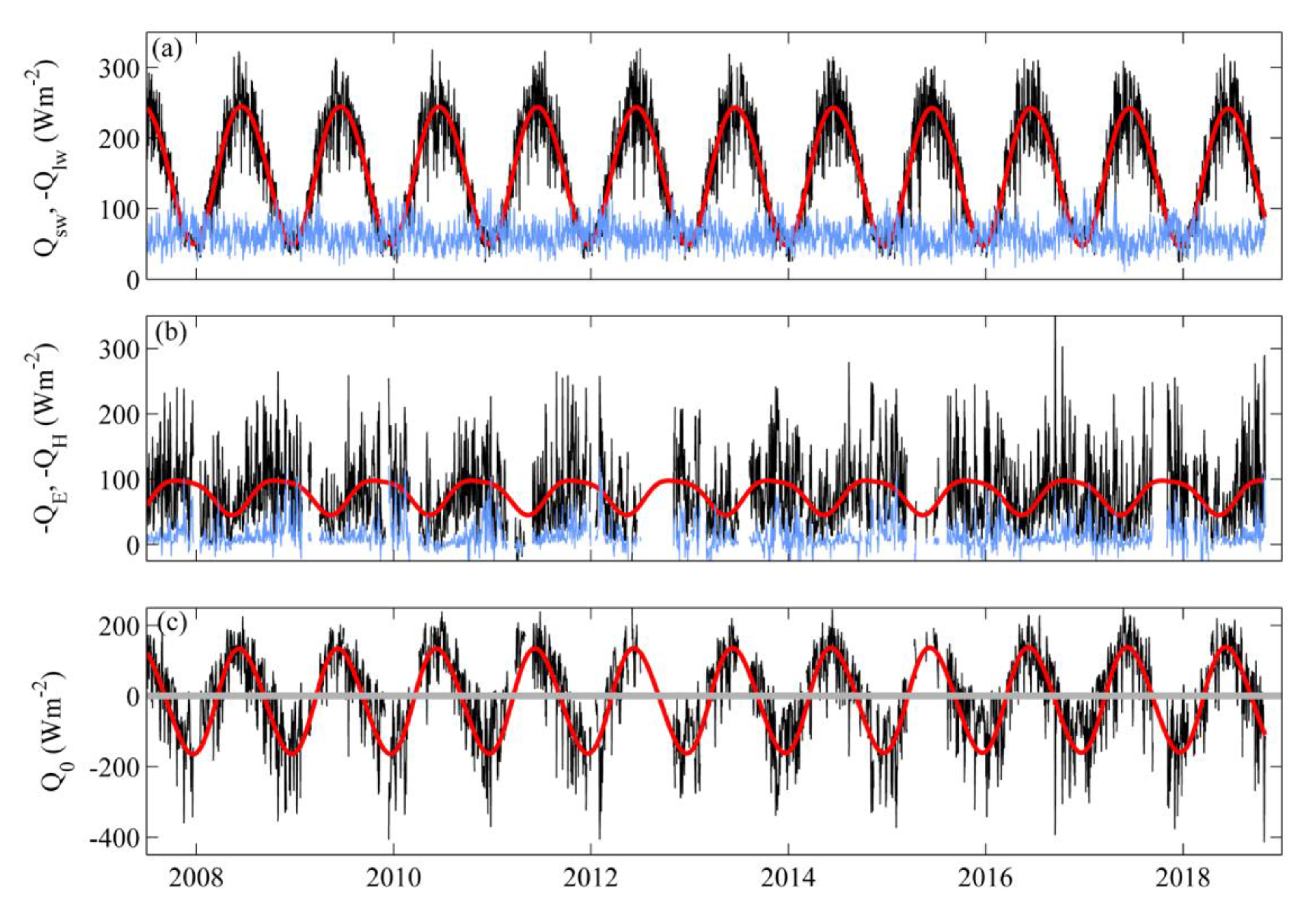
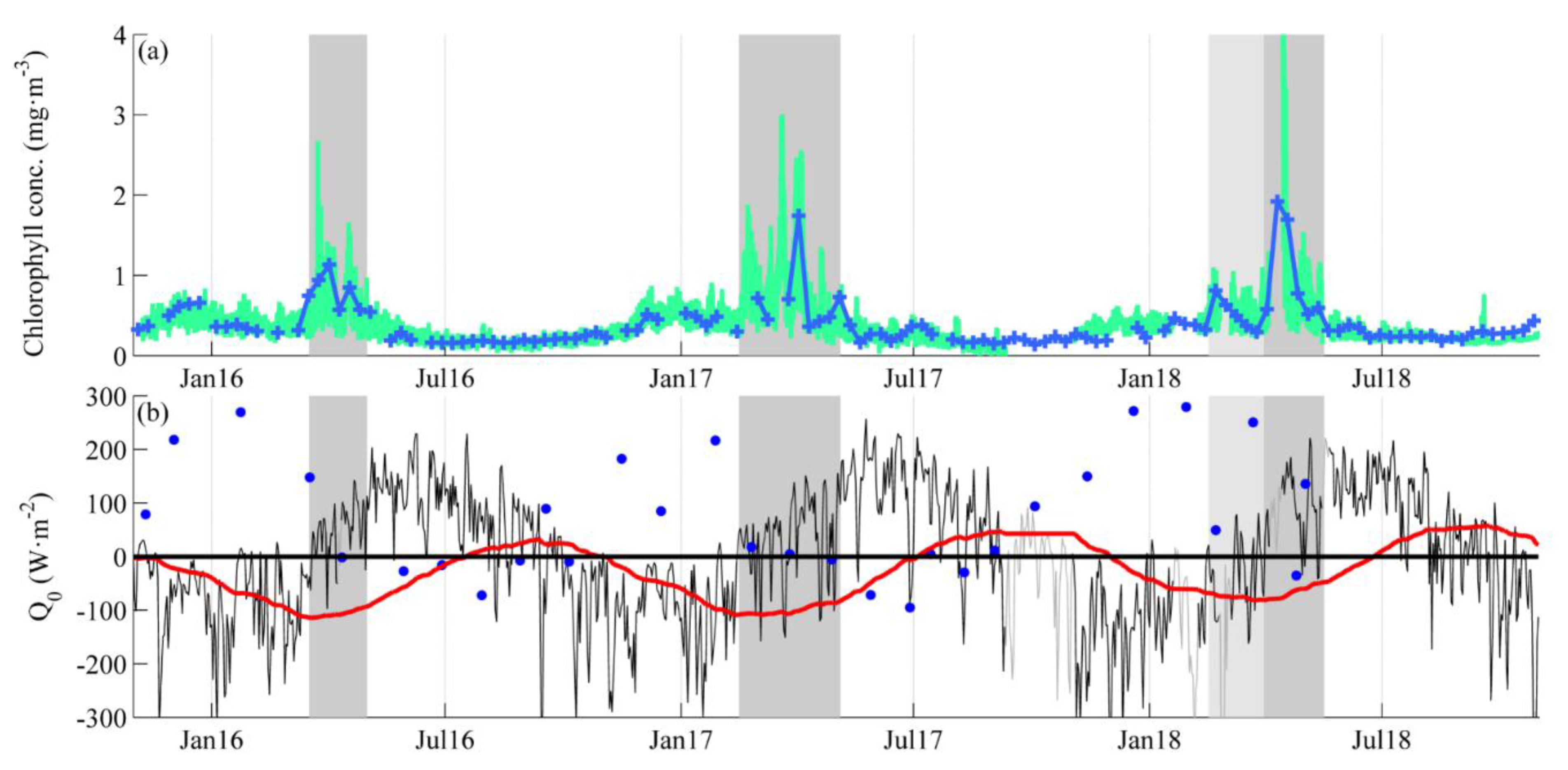
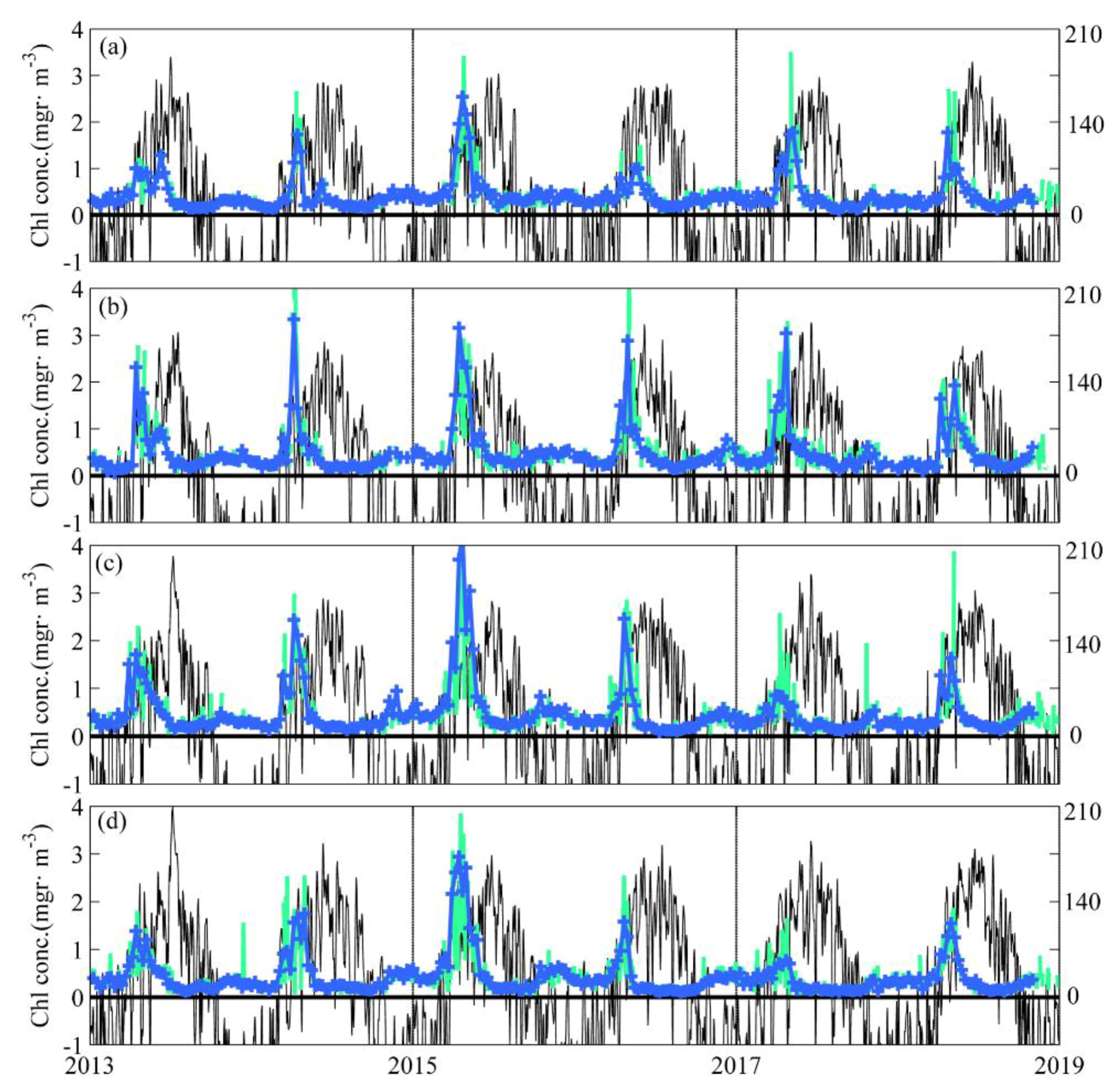
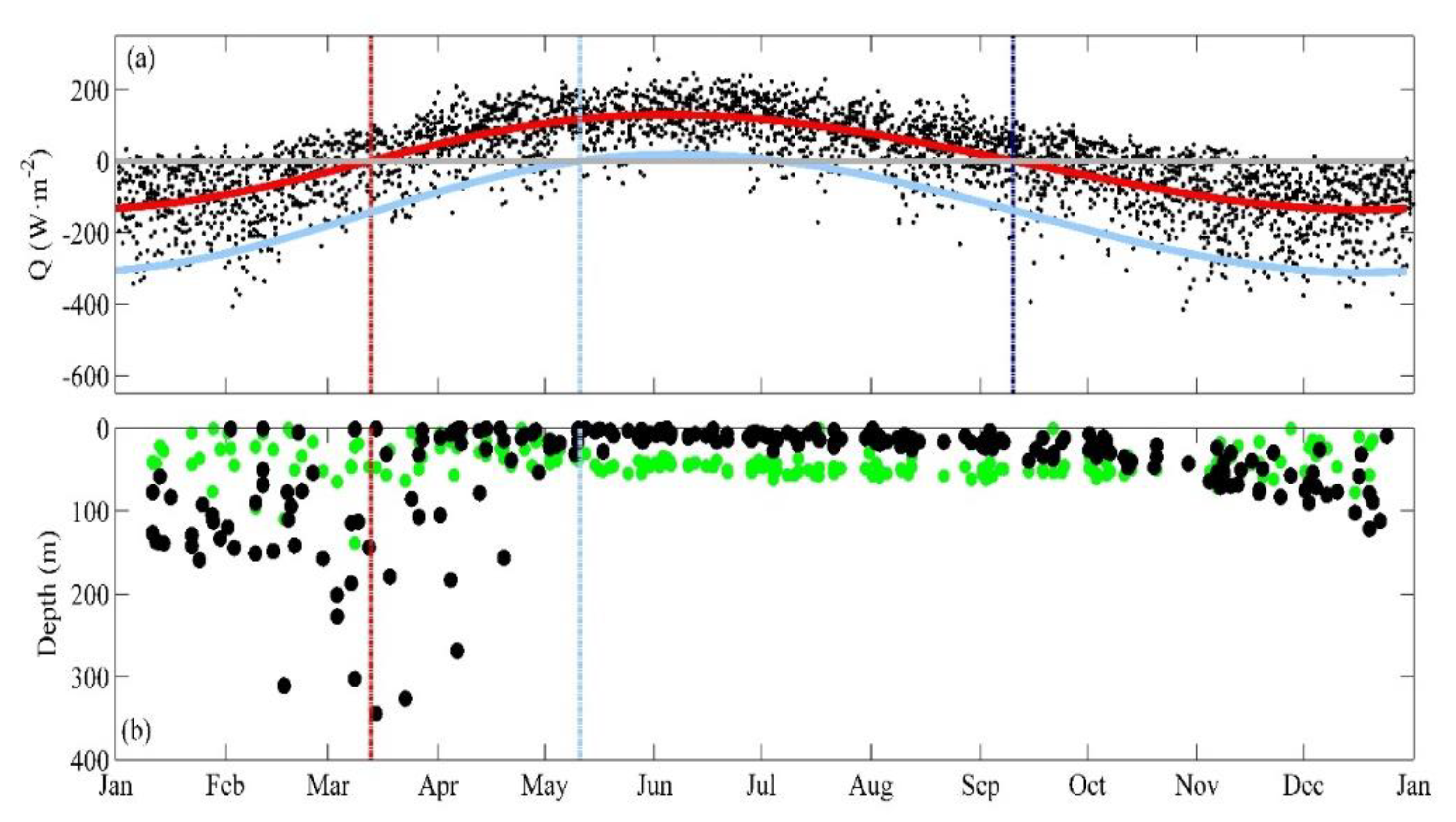
3.1. Near-Surface Phytoplankton Growth Signature and Changes in Air-Sea Exchanges
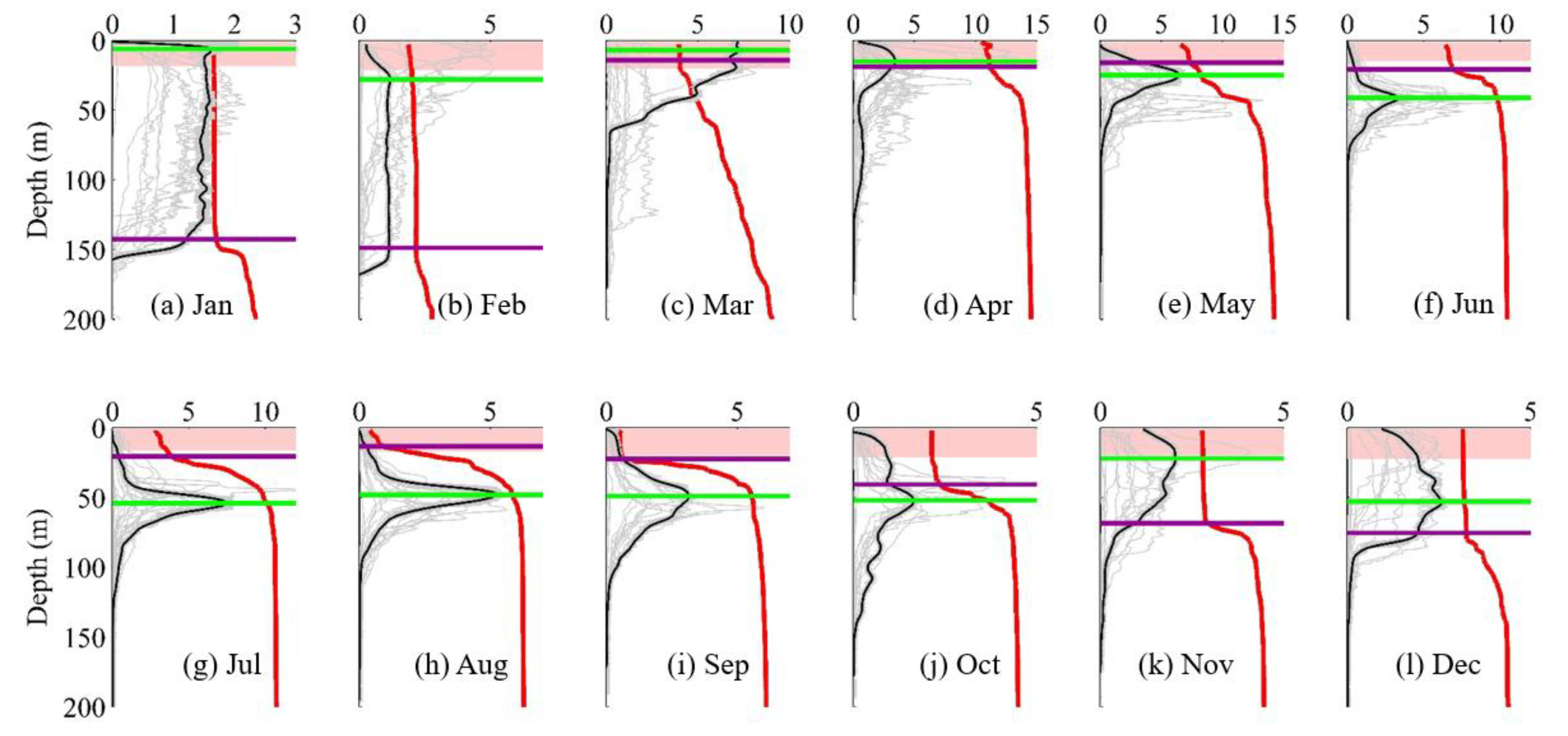
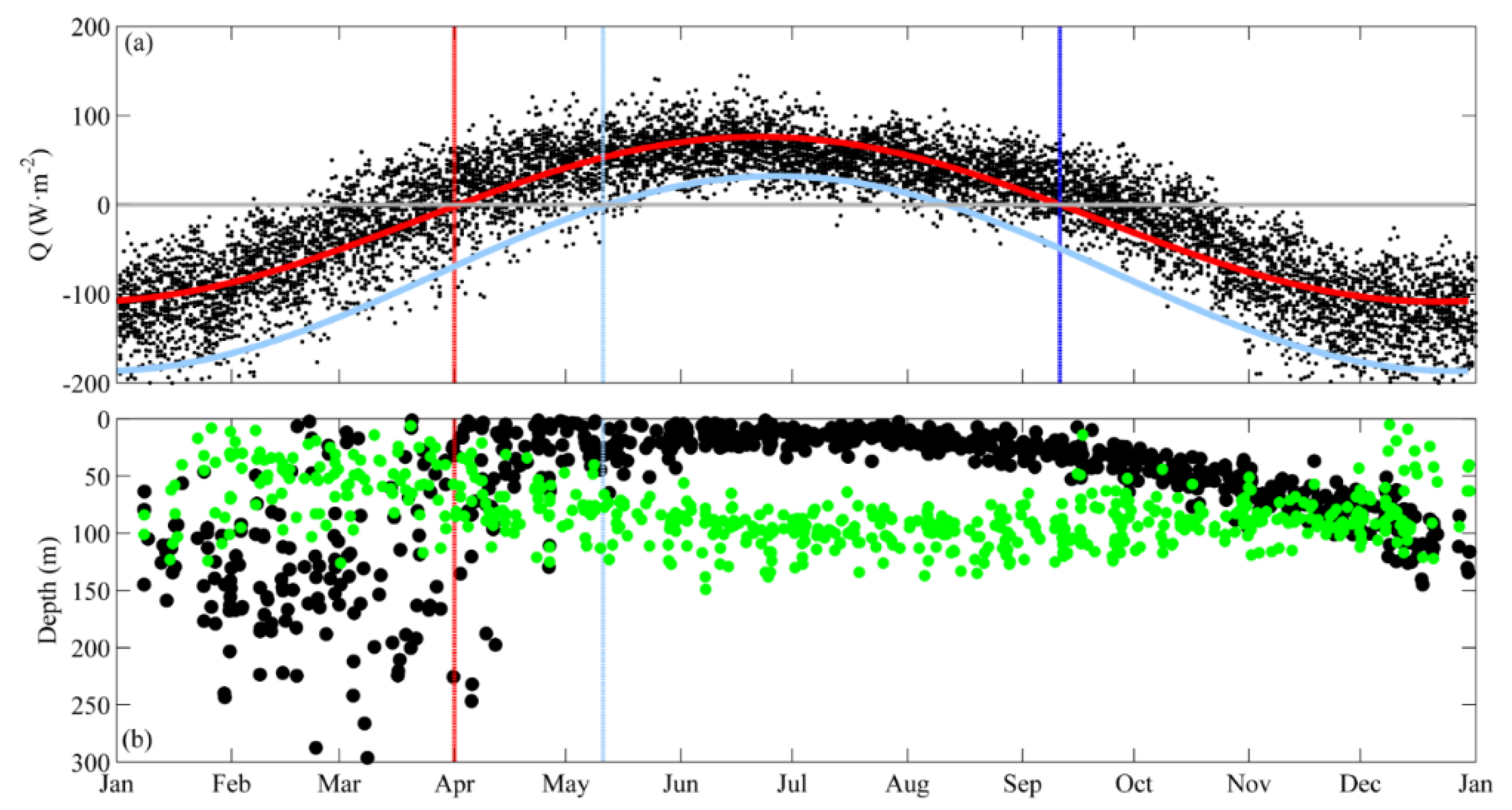
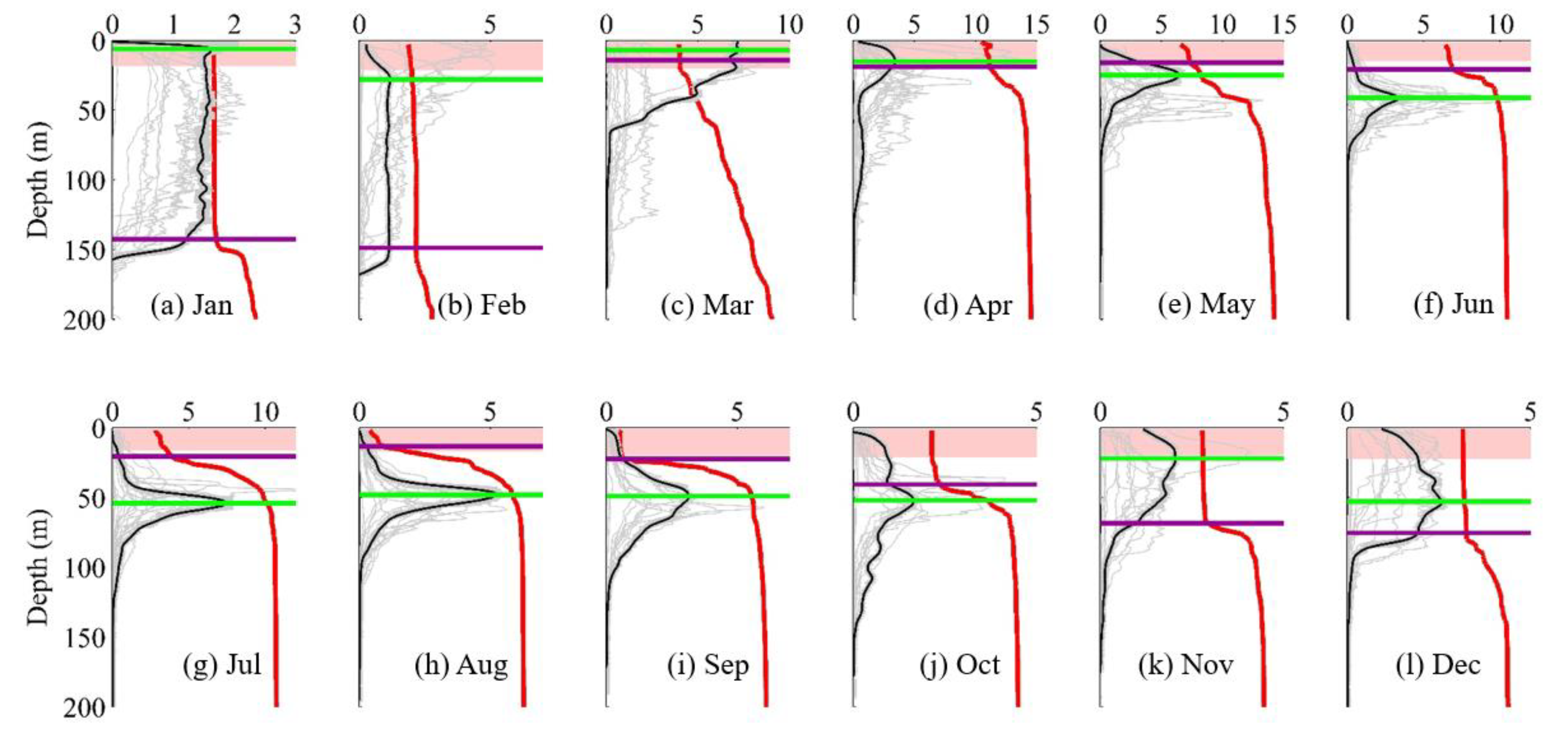
3.2. DCM Development: Change in Chlorophyll and Hydrographic Vertical Structure
3.3. Applicability of Our Results to other Ocean Regions and Further Remarks
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
References
- Sverdrup, H. On conditions for the vernal blooming of phytoplankton. ICES J. Mar. Sci. 1953, 18, 287–295. [Google Scholar] [CrossRef]
- Somavilla, R. Effects of Ocean-Atmosphere Interaction in the Upper Layers of the Bay of Biscay. Ph.D. Thesis, University of Cádiz, Cádiz, Spain, 2010. Available online: https://dialnet.unirioja.es/servlet/tesis?codigo=51752 (accessed on 10 July 2010).
- Taylor, J.R.; Ferrari, R. Shutdown of turbulent convection as a new criterion for the onset of spring phytoplankton blooms. Limnol. Oceanogr. 2011, 56, 2293–2307. [Google Scholar] [CrossRef] [Green Version]
- Behrenfeld, M.J. Abandoning Sverdrup’s Critical Depth Hypothesis on phytoplankton blooms. Ecology 2010, 91, 977–989. [Google Scholar] [CrossRef] [PubMed]
- Latasa, M.; Cabello, A.; Morán, X.; Massana, R.; Scharek, R. Distribution of phytoplankton groups within the deep chlorophyll maximum. Limnol. Oceanogr. 2017, 2, 665–685. [Google Scholar] [CrossRef]
- Herbland, A.; Voituriez, B. Hydrological structure analysis for estimating the primary production in the tropical Atlantic Ocean. J. Mar. Res. 1979, 37, 87–101. [Google Scholar]
- Cullen, J.J. Subsurface chlorophyll maximum layers: Enduring enigma or mystery solved? Annu. Rev. Mar. Sci. 2015, 7, 207–239. [Google Scholar] [CrossRef] [PubMed]
- Waniek, J.; Holliday, N. Large-scale physical controls on phytoplankton growth in the Irminger Sea, Part ii: Model study of the physical and meteorological preconditioning. J. Mar. Syst. 2006, 59, 219–237. [Google Scholar] [CrossRef]
- Behrenfeld, M.; Boss, E. Resurrecting the Ecological Underpinnings of Ocean Plankton Blooms. Mar. Sci. 2014, 6, 167–194. [Google Scholar] [CrossRef] [PubMed]
- González-Pola, C.; Lavín, A.; Vargas-Yánez, M. Intense warming and salinity modification of intermediate water masses in the southeastern corner of the Bay of Biscay for the period 1992–2003. J. Geophys. Res. Oceans 2005, 110, 1–14. [Google Scholar] [CrossRef]
- Somavilla, R.; González-Pola, C.; Rodriguez, C.; Josey, S.A.; Sánchez, R.F.; Lavín, A. Large changes in the hydrographic structure of the Bay of Biscay after the extreme mixing of winter 2005. J. Geophys. Res. 2009, c01001. [Google Scholar] [CrossRef]
- Somavilla, R.; González-Pola, C.; Villarreal, M.; Lavín, A. Mixed layer depth variability in the southern Bay of Biscay. Deepening of winter MLDs concurrent to generalized upper water warming trends? Ocean Dyn. 2011, 61, 1215–1235. [Google Scholar] [CrossRef]
- Somavilla, R.; González-Pola, C.; Schauer, U.; Budéus, G. Mid-2000s North Atlantic shift. Heat budget and circulation changes. Geophys. Res. Lett. 2016, 43, 2059–2068. [Google Scholar] [CrossRef] [PubMed]
- Somavilla, R.; González-Pola, C.; Fernández-Diaz, J. The warmer the ocean surface, the shallower the mixed layer. How much of this is true? J. Geophys. Res. Oceans 2017, 122, 7698–7716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalnay, E.; Kanamitsu, M.; Kistler, R.; Collins, W.; Deaven, D.; Gandin, L.; Iredell, M.; Saha, S.; White, G.; Woollen, J.; et al. The NCEP/NCAR 40-year reanalysis project. Bull. Am. Meteorol. Soc. 1996, 77, 437–471. [Google Scholar] [CrossRef]
- Somavilla, R.; Lavín, A.; Rodriguez, C.; Cano, D. AGL buoy. A high frequency view of processes at the air-sea interface. In Proceedings of the IEEE Oceans 2011-Europe, Santander, Spain, 6–9 June 2011. [Google Scholar] [CrossRef]
- Fairall, C.; Bradley, E.; Rogers, D.; Edson, J.; Young, G. Bulk parameterization of air-sea fluxes for tropical ocean global atmosphere coupled ocean atmosphere response experiment. J. Geophys. Res. 1996, 101, 3747–3764. [Google Scholar] [CrossRef]
- Yu, L.; Jin, X.; Weller, R.A. Multidecade Global Flux Datasets from the Objectively Analyzed Air-Sea Fluxes (OAFlux) Project: Latent and Sensible Heat Fluxes, Ocean Evaporation, and Related Surface Meteorological Variables; OAFlux Project Technical Report: Woods Hole, MA, USA, 2008; OA-2008-01, 64. [Google Scholar]
- Cronin, M.; McPhaden, M. The upper ocean heat balance in the western equatorial pacific warm pool during September–December 1992. J. Geophys. Res. 1997, 102, 8533–8553. [Google Scholar] [CrossRef]
- Somavilla, R.; González-Pola, C.; Lavín, A.; Rodriguez, C. T and S variability in the south-eastern corner of the Bay of Biscay (NE Atlantic). J. Mar. Syst. 2013, 109–110 (Supplement), S105–S120. [Google Scholar] [CrossRef]
- González-Pola, C.; Fernández-Diaz, J.M.; Lavín, A. Vertical structure of the upper ocean from profiles fitted to physically consistent functional forms. Deep Sea Res. Part I 2007, 54, 1985–2004. [Google Scholar] [CrossRef]
- Sauzède, R.; Claustre, H.; Jamet, C.; Uitz, J.; Ras, J.; Mignot, A.; D’Ortenzio, F. Retrieving the vertical distribution of chlorophyll a concentration and phytoplankton community composition from in situ fluorescence profiles: A method based on a neural network with potential for global-scale applications. J. Geophys. Res. Oceans 2015, 120. [Google Scholar] [CrossRef]
- Kantha, L.H.; Clayson, C.A. Small Scale Processes in Geophysical Fluid Flows; International Geophys; Academic Press: Cambridge, MA, USA, 2000; Volume 67. [Google Scholar]
- Alexander, M.A.; Scott, J.D.; Deser, C. Processes that influence sea surface temperature and ocean mixed layer depth variability in a coupled model. J. Geophys. Res. 2000, 105, 16823–16842. [Google Scholar] [CrossRef] [Green Version]
- Cronin, M.F.; Sprintall, J. Wind and buoyancy-forced upper ocean. In Encyclopedia of Ocean Sciences; Steele, J.H., Turekian, K.K., Thorpe, S.A., Eds.; Academic Press Ltd.: Cambridge, MA, USA, 2011; pp. 3219–3227. [Google Scholar]
- Moum, J.; Lien, R.-C.; Perlin, A.; Nash, J.D.; Gregg, M.C.; Wiles, P.J. Sea surface cooling at the Equator by subsurface mixing in tropical instability waves. Nat. Geosci. 2009, 2, 761. [Google Scholar] [CrossRef]
- Moum, J.N.; Perlin, A.; Nash, J.D.; McPhaden, M.J. Seasonal sea surface cooling in the equatorial Pacific cold tongue controlled by ocean mixing. Nature 2013, 500, 64. [Google Scholar] [CrossRef]
- Frajka-Williams, E.; Rhines, P. Physical controls and interannual variability of the Labrador Sea spring phytoplankton bloom in distinct regions. Deep Sea Res. Part I Oceanogr. Res. Pap. 2010, 57, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Marchese, C.; Guardia, L.; Myers, P.; Bélanger, S. Regional differences and inter-annual variability in the timing of surface phytoplankton blooms in the Labrador Sea. Ecol. Indic. 2019, 96, 81–90. [Google Scholar] [CrossRef]
- Erickson, Z.; Thompson, A.; Cassar, N.; Sprintall, J.; Mazloff, M. An advective mechanism for deep chlorophyll maxima formation in southern Drake Passage. Geophys. Res. Lett. 2016, 43, 10846–10855. [Google Scholar] [CrossRef]
- Cullen, J.J. The deep chlorophyll maximum: Comparing vertical profiles of chlorophyll a. Can. J. Fish. Aquat. Sci. 1982, 39, 791–803. [Google Scholar] [CrossRef]
- Mitchell, B.G.; Brody, E.A.; Holm-Hansen, O.; McClain, C.; Bishop, J. Light limitation of phytoplankton biomass and macronutrient utilization in the Southern Ocean. Limnol. Oceanogr. 1991, 36, 1662–1677. [Google Scholar] [CrossRef] [Green Version]

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Somavilla, R.; Rodriguez, C.; Lavín, A.; Viloria, A.; Marcos, E.; Cano, D. Atmospheric Control of Deep Chlorophyll Maximum Development. Geosciences 2019, 9, 178. https://doi.org/10.3390/geosciences9040178
Somavilla R, Rodriguez C, Lavín A, Viloria A, Marcos E, Cano D. Atmospheric Control of Deep Chlorophyll Maximum Development. Geosciences. 2019; 9(4):178. https://doi.org/10.3390/geosciences9040178
Chicago/Turabian StyleSomavilla, Raquel, Carmen Rodriguez, Alicia Lavín, Amaia Viloria, Elena Marcos, and Daniel Cano. 2019. "Atmospheric Control of Deep Chlorophyll Maximum Development" Geosciences 9, no. 4: 178. https://doi.org/10.3390/geosciences9040178
APA StyleSomavilla, R., Rodriguez, C., Lavín, A., Viloria, A., Marcos, E., & Cano, D. (2019). Atmospheric Control of Deep Chlorophyll Maximum Development. Geosciences, 9(4), 178. https://doi.org/10.3390/geosciences9040178