2. Palaeogeography in the North of Britain during the Middle Jurassic
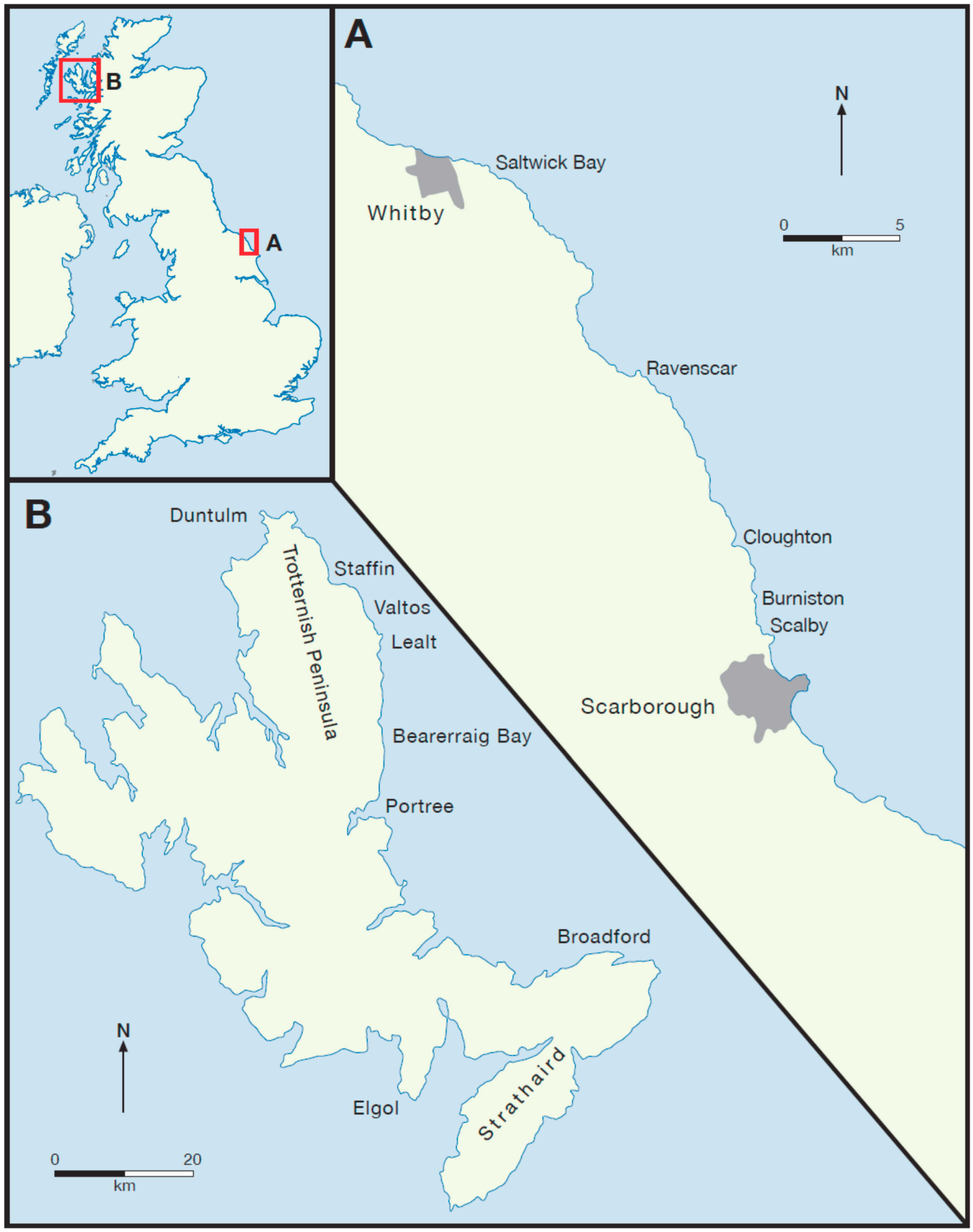
In Yorkshire, rocks of Middle Jurassic age (Aalenian to Bathonian) constitute the Cleveland Basin, while on the Isle of Skye (Aalenian to Callovian) they form part of the Hebrides Basin, a composite basin including the Canna Basin and Coll Basin within the Little Minch Trough south of the Minch Fault, and the Inner Hebrides Basin south of the Skerryvore and Camasunary faults [
10]. During this time, much of Skye was a coastal topographic high known as the Skye High [
11,
12], with a number of transgressive and regressive sequences throughout the Aalenian to Callovian [
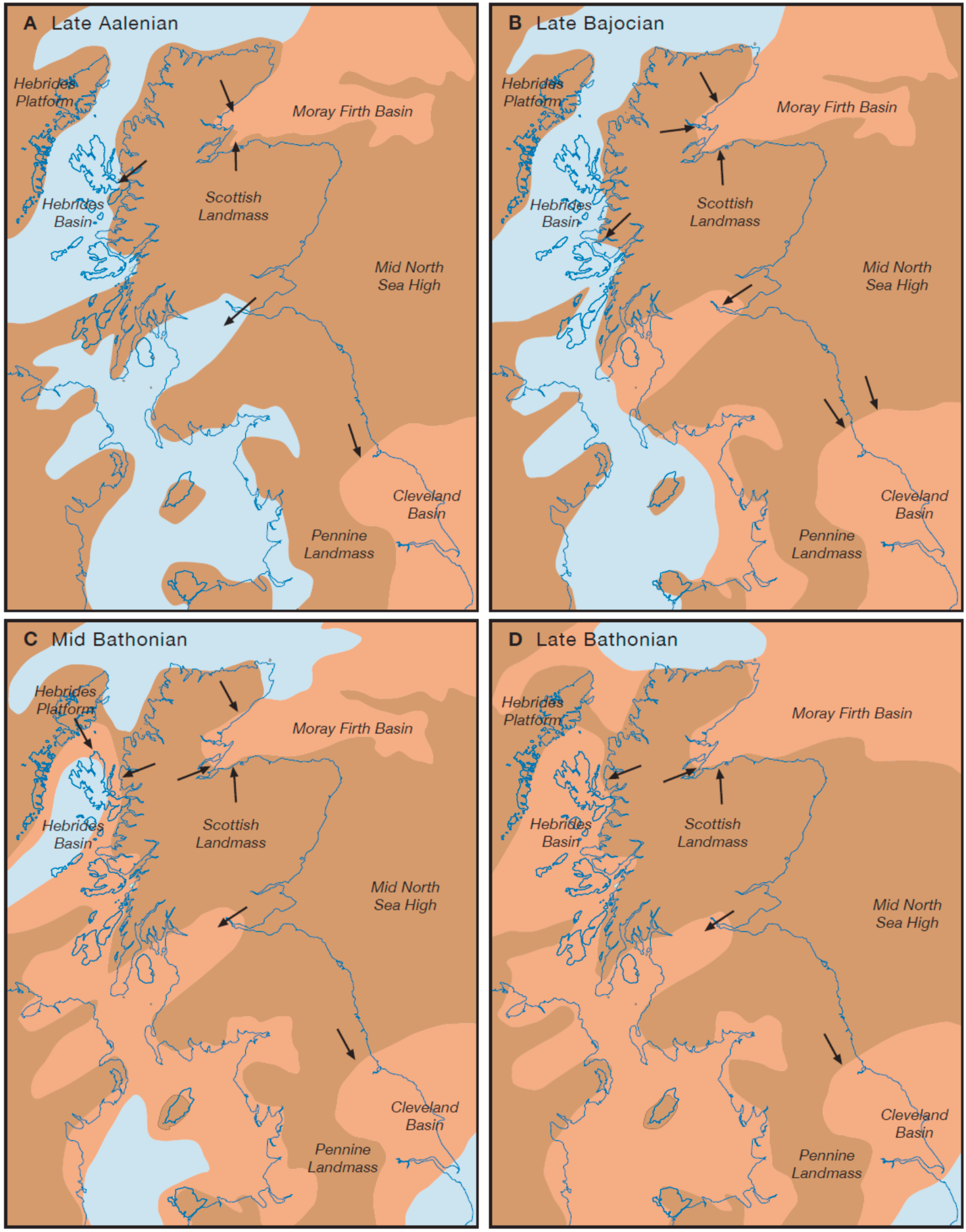
13]. In Early Jurassic times, the Cleveland and Hebrides basins were part of a generally shallow shelf sea, with a large area of land over Scotland made up of the northern Shetland Platform and a connected southern Scottish Landmass (
Figure 2A) [
14]. By early Middle Jurassic times, following the late Aalenian marine transgression, an emergent Mid North Sea High had developed adjacent to the Scottish Landmass, and the Pennine Landmass formed an extension of the Scottish landmass further south. These latter two landmasses provided the source for sediment from rivers draining off into the start of the Cleveland Basin depocentre (
Figure 2B). More or less contemporaneously, the Hebrides Basin developed off to the west of the Scottish Landmass with sediment being derived from this source, but isolated from the Cleveland Basin and possibly with no coastal plain contact with it (
Figure 2B). This scenario was maintained without any major changes throughout the rest of the Middle Jurassic, except that occasionally coastal plains were possibly more extensive and could be traced continuously from one basin to the other (
Figure 2C,D).
These two basins in the Middle Jurassic were at least 400 km apart, separated by landmasses, which, though providing clastic sediment for the basins, were not of sufficient height (nor subjected to such intense climatic conditions) as to provide much coarse-grained sediment to the basins. The general absence of coarse-grained sandstones and siliciclastic clasts in the Middle Jurassic sediments of the Cleveland Basin suggest that they were deposited in an area of low available relief and relatively low flow regimes. The sediments of the Hebrides Basin also lack the coarse-grained sandstones with the first minor lenticular quartz pebble conglomerates, derived from the west, appearing in the Brora Sandstone Member of the Brora Arenaceous Formation (Oxfordian) near Brora, although the late Bathonian Skudiburgh Formation also contains intraformational conglomerates at Port Duntulm, Isle of Skye [
15,
16]. The terrestrial trace fossils found in the two basins indicate that diverse dinosaur communities existed in both.
3. Comparison of Middle Jurassic Lithostratigraphy and Facies of the Cleveland and Hebrides Basins
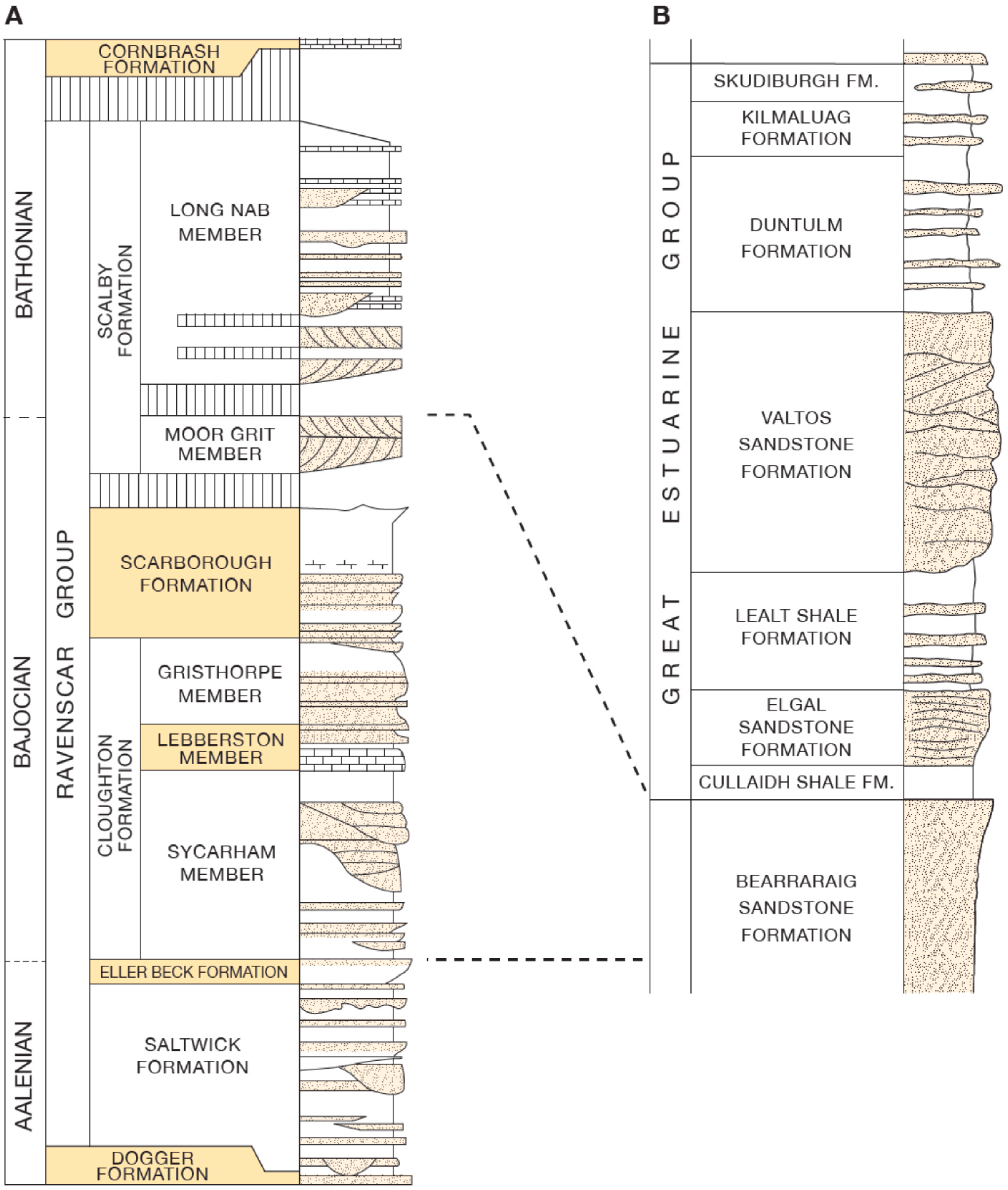
The dinosaur print-bearing units from the Middle Jurassic sequences of the Cleveland Basin range from Aalenian to Bathonian in age, and are assigned to the Ravenscar Group, a unit approximately 220 m thick and comprising five formations (
Figure 3). The sequence is dominantly non-marine, consisting of sandstones and mudrocks, impure coals and ironstones, but interbedded with three marine units which contain impure argillaceous limestones and marine shelly faunas. The Bathonian dinosaur print-bearing strata on the Isle of Skye is approximately 250 m thick and belongs to the Great Estuarine Group (
Figure 3). The Skye sediments are also a complex interplay of coarse and fine-grained siliciclastic sediments lagoonal, fluvial of varying salinities [
13]. Both basins are formed of sediments of the optimal consistency for the preservation of dinosaur footprints [
17,
18].
The rocks of the Ravenscar Group are now generally regarded as being a coastal plain and fluvial complex [
19], represented by paralic fluvial and lacustrine lithofacies (Saltwick, Cloughton and Scalby formations) with three marine intercalations (Eller Beck Formation, a locally developed Lebberston Member within the Cloughton Formation, and Scarborough Formation) [
20,
21]. However, as pointed out by Eschard and others [
22,
23] some sedimentological features may also suggest a deltaic origin for parts of the sequence. The ribbon sheet sandstones of the non-marine units represent crevasse splays and levee deposits, while thicker saucer-shaped sandstone bodies indicate lenticular channel sandstones. The thick mudrocks interbedded within the sequence may have accumulated in flood plains, shallow lakes and marshes, brackish lagoons or abandoned river channels. Locally, peaty mires accumulated, thick enough to form thin coals [
21]. Plant remains such as ferns and horsetails are very common and diverse [
24] and occur as drifted leaves [
25], transported logs and in situ erect stems, while conifer remains were transported into the basin from the hinterland. However, invertebrate body fossils are very rare. Only bivalves (
Unio) have been recorded from the Saltwick and Scalby formations, and bivalve escape structures (
Lockeia) are common. Other invertebrate traces from the non-marine units are locally more abundant, and include
Cochlichnus,
Protovirgularia,
Beaconites (? =
Taenidium) [
26],
Selenichnites [
27,
28,
29,
30],
Kouphichnium [
31,
32],
Diplocraterion and simple burrows.
The Great Estuarine Group on Skye is represented by the Cullaidh, Elgol, Lealt Shale, Valtos Sandstone, Duntulm, Kilmaluag and Skudiburgh formations. The underlying Bearerraig Sandstone (Aalenian–Bajocian) exhibits two major transgressions that are also evident in the Cleveland Basin [
33]. This formation represents a period of marine water cooling by as much as 10 °C perhaps as a result of thermal doming and resulting changes to ocean currents [
34]. The Great Estuarine Group has two main phases of sand deposition separated by muddier sediments and terminating with the fluvial deposits of the Skudiburgh Formation (channel sands, muds and calcrete) [
13,
35]. The sandbodies of the Valtos Sandstone Formation represent lagoonal shorelines and deltas prograded from north to south as they did in the earlier part of the Great Estuarine Group on Skye during the deposition of the Elgol Formation [
13,
36].
The muddier horizons within the Great Estuarine Group on Skye generally represent lagoonal conditions. The Lealt Shale Formation represents a brackish lagoon, whereas the mudstones of the Duntulm Formation are brackish to more normal marine salinities and the Kilmaluag Formation is predominantly freshwater [
13,
35,
36]. The environment of deposition within the Duntulm Formation is complex and has been interpreted as ranging from freshwater carbonate muds and sands, supralittoral carbonate algal marshes, brackish-marine lagoons with an abundance of the fossil oyster
Praeexogyra hebridica [
37], small prograding delta sands and, towards the top of the formation, freshwater muds and sands [
38,
39]. Some of the trackway surfaces in the Duntulm Formation are rippled and do not appear to exhibit desiccation cracks that are commonly seen in the Valtos and Kilmaluag formations and contains abundant
Diplocraterion trace fossils [
39,
40]. At the end of the Great Estuarine Group on Skye, a more off-shore marine environment existed by the Middle Callovian caused by a widespread change in relative sea-level with mudstones in the Trotternish area and marine sandstones in the Strathaird Inner Hebrides Basin [
13].
4. Dinosaur Body Fossils of the Cleveland and Hebrides Basins
Dinosaur body fossils are rare from the Middle Jurassic Ravenscar Group of the Cleveland Basin. The first record of such material was from White Nab, near Scarborough. This was described by Williamson [
41] who assigned one of the bones, a fragment of a vertebra, as possibly ‘
Megalosaurus’; paradoxically the specimens were from the marine Scarborough Formation. Unfortunately, Williamson’s material cannot now be confidently identified [
42]. There were no further records published of dinosaur bones from the Middle Jurassic of Yorkshire until the Sheffield Dinosaur Track Research Group (see below) began work on dinosaur trace fossils from the area. These finds were tabulated by Romano and Whyte [
5] when four new horizons were identified. Seven years later, the number of horizons with skeletal finds had increased to eight [
42], most of these from channel deposits. However, among these, the most significant is an isolated caudal vertebra, from a palaeosol in the Saltwick Formation, of an early sauropodomorph that is at present the earliest stratigraphic occurrence for a British sauropod dinosaur [
43]. Although new finds are continually being reported, it still remains an enigma that they are comparatively rare, and particularly in the complete absence of teeth. This may be due in part to ‘rapid solution by the acidic groundwaters and seasonally high water tables that are made evident by the sphaerosiderite-rich, gley palaeosols’ [
42].
Body fossils are also rare in the Hebridean area, although there are quite a few different groups represented by the bony remains [
44]. The first bone to be found was a partial theropod tibia from the Sinemurian (Early Jurassic) of Strathaird, but every body-fossil found since then has been from the Great Estuarine Group in both Trotternish and Strathaird. Many of the bones found have not been diagnosable beyond dinosaur, but several have been assigned to different groups. The first Middle Jurassic dinosaur body fossil to be described from the Isle of Skye was a limb-bone, possibly the left humerus, of a basal eusauropod [
44,
45,
46,
47] from the Valtos Sandstone Formation. Further remains from this formation included a caudal vertebra [
46] a rib bone and a tooth [
47] all of which belong to a primitive eusauropod. A further sauropod tooth found in the Strathaird section of the Kilmaluag Formation could also belong to a primitive eusauropod or a basal titanosauriform [
48].
Theropod remains are very rare in the Hebrides Basin with only a few teeth fragments and a single caudal vertebra [
49]. The tooth is thought to representative of either a megalosaurid, basal tyrannosauroid, or dromaeosaurid, and that the vertebra may belong to a small-bodied basal coelurosaur [
49]. However, both of these remains lack the diagnostic characteristics that would allow confident assignment to any particular clade and have been left as Theropoda indet [
49]. The only other theropod remains recorded is a tooth fragment from the Fish Bed of the Lealt Shale Formation on the Isle of Eigg [
50].
The only other dinosaur body fossil that has been described from the Middle Jurassic of Skye is a proximal ulna and radius of a thyreophoran dinosaur, perhaps a basal europodan akin to either the ankylosaurs or stegosaurs [
51]. This was found in the older Bajocian Bearerraig Sandstone Formation near Storr Lochs on the Trotternish Peninsula of the Isle of Skye in association with a possible humerus that was subsequently removed by a collector before the local museum was able to retrieve it [
51].
There are many bone fragments, teeth, and more complete bones awaiting preparation and description in the collections of the Hunterian, the National Museums of Scotland and the Staffin Museum. These are currently being studied by the PalAlba group, a consortium of researchers based in Scotland.
5. Brief History of Dinosaur Ichnite Research from the Cleveland and Hebrides Basins
The first record of vertebrate tracks from the Cleveland Basin was in the early twentieth century when Hargreaves [
52] reported that a track found by Mr Rowntree about 18 years earlier from south of Scarborough had been identified by Mr Lamplugh as ‘probably crocodilian’. It is more likely that it was made by a dinosaur, although since the specimen has not been traced it has not proved possible to confirm this [
41].
Brodrick was the first to describe dinosaur tracks from the Cleveland Basin in a series of papers [
53,
54,
55,
56]; all the tracks were of tridactyl forms and were from outcrops around Whitby (Saltwick Formation of Aalenian age, see below). The recording of new finds then declined, following the onset of World War 1 and World War 2 [
42], and only began to show some renaissance by the mid 20th century [
52,
57]. However, it was not until the latter part of the 20th Century that interest was rejuvenated, when Sarjeant [
58] described
Satapliasaurus dsocenidzei Gabouniya [
58], the first named dinosaur ichnite from the Cleveland Basin. Valuable contributions from amateur geologists [
59] followed, and increasing numbers of papers were published on new finds from units of Aalenian to Bajocian age, particularly by the Sheffield Dinosaur Track Research Group [
42] who began the modern phase of ichnological studies [
42,
60,
61,
62,
63,
64,
65,
66]. A distinctive second dinosaur ichnite from the Cleveland Basin was subsequently described [
60] but was not formally named as
Deltapodus brodricki until two years later [
61]. At about this time, further new prints were described from the area and were referred to
Brontopodus-like and
Breviparopus-like [
65], soon followed by the recognition and description of dinosaur swimming tracks (
Characichnos tridactylus) [
62].
The history of dinosaur track research in the Hebrides Basin is in contrast to that in the Cleveland Basin. It was not until 1982 that the first dinosaur footprint (a single track of a purported ornithopod) was found from the Lealt Shale Formation (Bathoninan, see below) at Trotternish, Skye. This was reported in
The Scotsman newspaper [
67] and a full description was subsequently published in 1984 [
68]. Since then, many more footprints at other localities have been found and recorded from other units of Bathonian age [
39,
40,
45,
69,
70,
71,
72].
It is interesting to note that, on both the east Yorkshire coast and on the Isle of Skye, workers had for many years been undertaking research on other aspects of the geology without either noticing the prints, or considering them not significant or important enough to study! The recent years have happily countered this omission, resulting in that both areas are now considered significant on a global scale to help bridge the gap in our knowledge of dinosaurs during the Middle Jurassic.
6. Dinosaur Ichnology of the Ravenscar Group of the Cleveland Basin and Great Estuarine Group of the Hebrides Basin
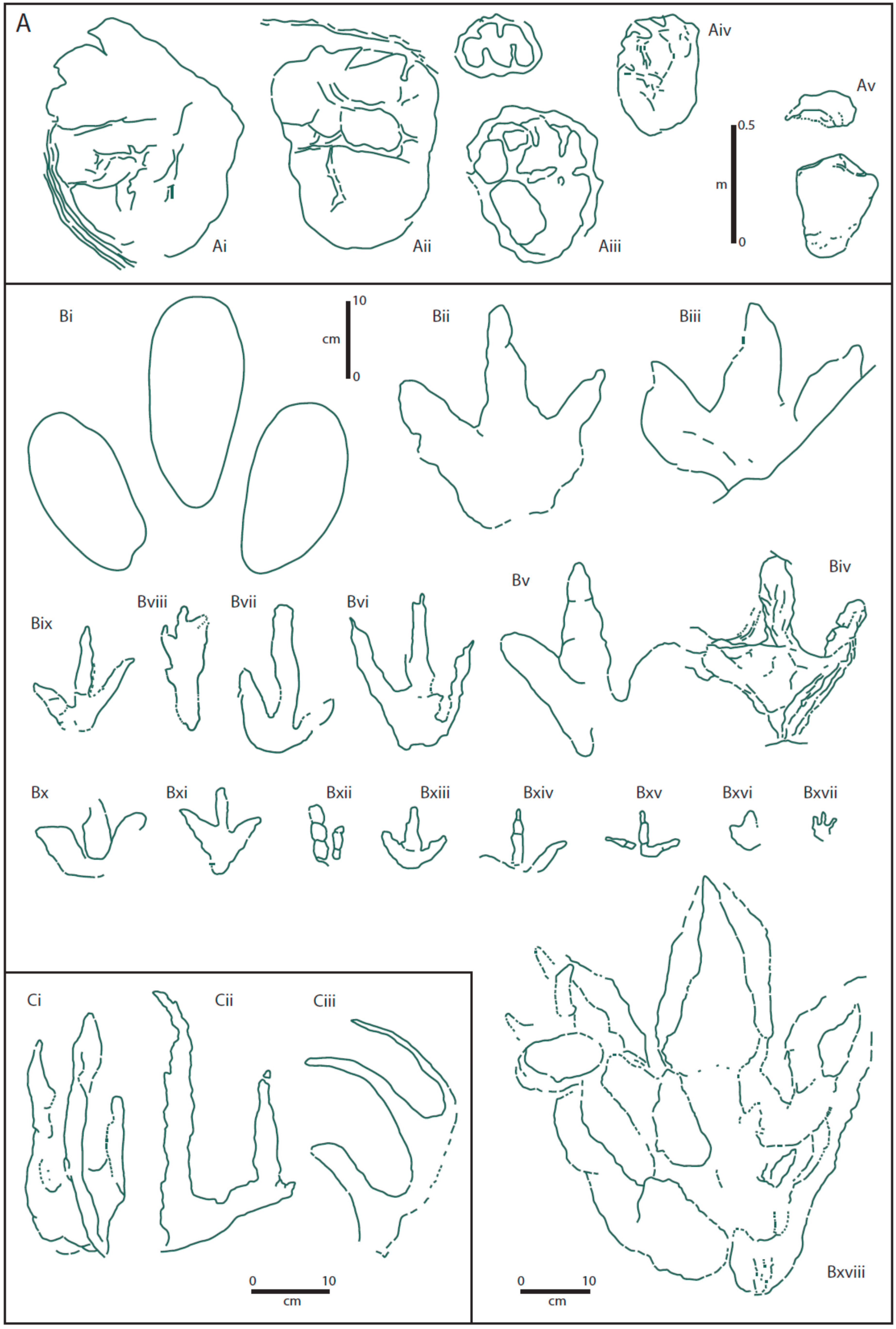
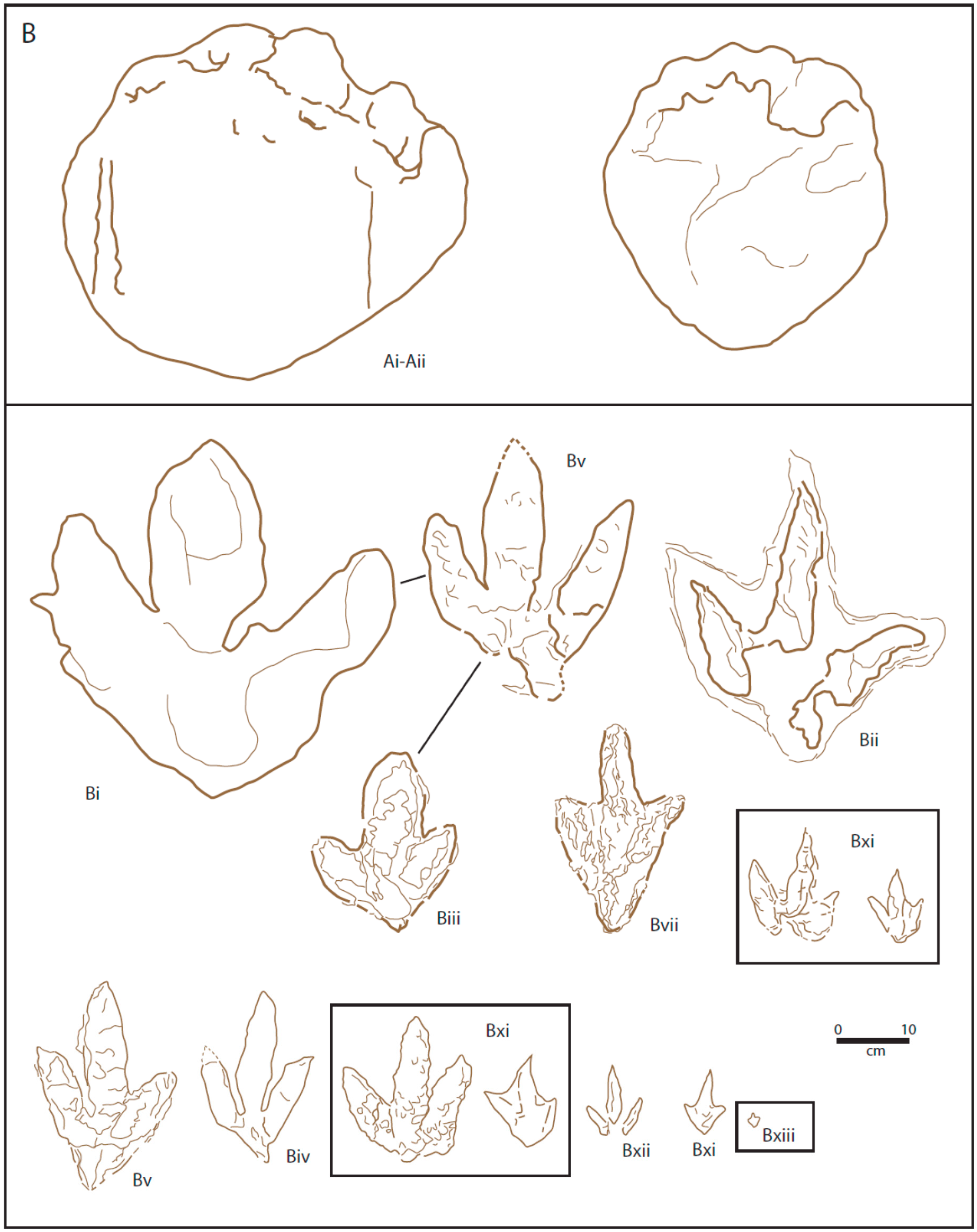
By the early part of the present century, 25 dinosaur print morphotypes had been identified from the Cleveland Basin [
5,
18] (
Figure 4A). At that time, the authors preferred not to refer to the different prints as ichnospecies, nor to regard the brief descriptions as diagnoses, since the general lack of trackways and paucity of a number of the print types prevented any clear conclusions as to whether they represented true ichnospecies or were preservational variants. The 25 morphotypes were divided into three groups: Group A—prints made by quadrupeds, Group B—prints made by bipeds, and Group C—swimming prints [
5,
18].
Group A included five (Ai-v) different morphotypes. Morphotypes Ai and Aii had previously been referred
Brontopodus-like and
Breviparopus-like respectively [
66]. Four years later, because of the general scarcity of manus-pes couples and trackways of prints of these types, these two morphotypes were still only listed as ‘
Brontopodus’ and ‘
Breviparopus’ [
18]. By 2012, a number of additional manus tracks had been recognized, yet none of these showed the ‘somewhat U-shaped’ manus outline of
Brontopodus birdi, the type ichnospecies of
Brontopodus [
73]. The five morphotypes Ai-v are now referred to the following ichnotaxa: Ai—
Brontopodus ichnosp.; Aii, Aiii—
Breviparopus-like ichnosp. [
65]; Aiv—Ichnogen et ichnosp. indet [
64].
The final print morphotype in Group A (Av) was assigned to a new ichnotaxon,
Deltapodus brodricki [
61], after the distinctive ‘delta-shaped’ pes imprint and the name of the person who first described dinosaur prints from the area. This print type has now been recognized across the Jurassic world and has been recorded from Spain, Portugal, Morocco and Utah (USA) [
74], and most recently in the Cretaceous of China [
75].
Group B contained 17 tridactyl prints that had been made by bipeds (
Figure 4). While nearly all are certainly pes prints, Bxvii is most probably a manus print [
5]. Only one (
Satapliasaurus dsocenidzei—Biii) was named at that time [
58], although this identification was questioned by Lockley and Meyer [
8] who suggested that it should perhaps be treated with caution, while one other (Bxii) was assigned to cf. ‘
Grallator’. Since then, other prints in Group B (Bii, Bv, Bxii and more recently, Bxviii) have been tentatively assigned to a named ichnotaxon, such as
Eubrontes,
Megalosauropus or ichnotaxa from the
Anchisauripus-
Grallator series [
76,
77]. The most recently found large print Bviii followed the recording of even larger tridactyl tracks that had been sporadically found from the Scalby Formation [
59,
78]. The problem of whether the remaining prints in Group B may be assigned to distinct ichnospecies is illustrated with respect to print types Bix, Bviii and Bxi. All three of these print types were made by the same animal in an ichnoassemblage that shows prints with a range of morphologies [
8]. Print type Bviii shows imprints of the metatarsal area [
5], which resulted from the animal sinking lower in the sediment and so recording the greatest degree of metatarsal extension. Had a trackway been formed entirely of this type of print (Bviii), it would have been justified in naming the trackway; but an isolated print of this type within an assemblage of prints of varying morphologies does not provide the ichnotaxonomic basis for giving Bviii a different or new name.
Among the most distinctive tridactyl prints in Group B are the small, gracile prints Bxiv and Bxv (
Figure 4), with slender digit prints, showing evidence of phalangeal pads and very wide divarication. Though distinctive, their rarity has not allowed a complete understanding of their morphology, and so they have not been assigned to any ichnotaxon.
Within Group C (swimming group), print type Ci had earlier been described and named as
Characichnos tridactylus [
62]. The two other print types in Group C, Cii and Ciii (
Figure 4), were later reinterpreted as part of a trackway which was produced by the same animal type as that which made
Deltapodus brodricki (see below). These print types were assigned to
Characichnos isp. on the grounds that the limited number of prints in the trackway is insufficient to propose a new ichnospecies [
79].
No attempt has been made to define all the footprint morphotypes in the Hebrides Basin (
Figure 4B), but comparative morphometric analyses have been carried out to differentiate different tridactyl tracks using principal component analyses on non-subjective landmarks [
40,
70,
72]. As a result three different tridactyl tracks were distinguished on this basis [
40,
70,
72]. The first, and most abundant footprint type are the smaller (<20 cm length) footprints from the Valtos Sandstone and Kilmaluag formations (
Table 1. X;
Figure 4B—morphotype Biii, Biv, Bv, Bvii, Bxi, Bxii, Bxiii). The second are the larger footprints from the Valtos Sandstone Formation (
Table 1. Y;
Figure 4B Bv) and also includes the first dinosaur footprint to be found on Skye from the Lealt Shale Formation (
Figure 4B Bi); and the third group of tridactyl tracks are those large tracks found in the Duntulm Formation (
Table 1. Z;
Figure 4B Bii). A potential fourth group could not be analysed as it is represented by a single footprint from level B in the Duntulm Formation at An Corran (
Table 1. W;
Figure 4B Biii).
These footprints can also be looked at in terms of the morphotypes recognized from the Cleveland Basin [
18]. Variations in footprint preservation may be related to differences in the consistency and type of sediments between the Cleveland Basin and the Hebrides Basin. The footprint from the Lealt Shale Formation is about 49 cm long with broad digit impressions similar to morphotype Bi-Bii [
18]. In the Valtos Sandstone Formation, there are three size groups of tridactyl footprints; one based on a single track measuring over 100 cm in length, one large at about 40 cm in length, and another small at up to 28 cm in length [
45]. The first group is similar to the footprint from the Lealt Formation with large rounded digit impressions (Bi-Bii) and the other group are smaller tridactyl footprints ranging in size from 17 to 24 cm in length (Bv) [
69]. Only one small (40 cm long) sauropod footprint is known from the Valtos Formation [
47]. The footprint is possibly closest to the Aiv morphotype [
18], although it is unclear whether the digits curve at the straight anterior margin.
The larger tridactyl footprints at level A from the Duntulm Formation at An Corran appear to contain morphotypes Bii or Bv in terms of shape although they are larger than these morphotypes described from the Cleveland Basin at between 32 and 53 cm in length. From the same formation at level B at An Corran, a single footprint resembles more morphotype Biii and is 24 cm in length [
18,
70]. The large (~70 cm long) sauropod footprints from the Duntulm Formation at Cairidh Ghlumaig are similar to the Aii morphotype described as
Breviparopus-like [
18,
39,
66].
In the Kilmaluag Formation of Lub Score on the western side of the Trotternish Peninsula, the footprints vary greatly in size probably due to the co-existence of adult and juvenile of the same species. The footprints can range from 1.8 to 27 cm in length and their impressions are similar to morphotypes Bv and Bxiii [
72].
To date, no swimming tracks (see Group C morphotypes of the Cleveland Basin,
Figure 4) have been recorded from the Great Estuarine Group of Skye. This is perhaps difficult to explain since they are relatively common throughout the Ravenscar Group [
77] and yet similar facies occur in both basins.
7. Distribution of Print Types with Respect to Facies and Age
Prints within the Cleveland Basin are preserved in a variety of ways and in different facies [
18,
80]. Transmitted and underprints are most common, while surface prints less so [
5]. In addition, the facies in which they occur are extremely variable. Prints are most commonly found at the base of often thin, sheet sandstone beds (‘Burniston footprint bed’ of Hargreaves [
57]; and ‘
Deltapodus sandstone’ of Romano and Whyte [
18]), which are frequently the result of crevasse splays. However, the positive hypichnial prints, which protrude beneath the undersurface of these beds, were commonly not made at the interface between the underlying mudrocks and overlying sandstones. Sectioning through these prints [
18] frequently reveals the true tracking surface higher up (later) in the sequence. This then brings into question whether the animals that lived in the area preferred a sandy instead of muddy substrate on which to move around. Unfortunately, the frequently thick homogeneous mudrock sequences (often over a metre in thickness) will not yield evidence of foot emplacement in the absence of beds with contrasting sediment grain size. Consequently, the well-known ‘footprint beds’ are often the result of underprinting from a higher level.
Despite these apparent anomalies, the distribution of prints within sequences of varying facies indicates that the dinosaurs of the Cleveland Basin frequented the tops and slopes of point bars [
77], river levee deposits and interdistributary areas. In addition, they also probably inhabited the margins of waterholes, as made evident by the occasionally extensive and frequent dinoturbated beds [
18]. It is noteworthy that the relatively common prints attributed to sauropods occur in siliclastic facies, yet have frequently been linked to more marginal marine habitats [
1,
81].
The tridactyl footprints from the lowest unit of the Hebrides Basin (Lealt Shale Formation) are associated with more muddy environments, but may represent undertracks making it difficult to be certain of the sediment on which the animals walked [
68]. In the Valtos Sandstone, Duntulm and Kilmaluag formations, the tridactyl footprints are commonly associated with desiccation cracks that emanate from the digits suggesting that the animals walked on a damp substrate prior to the cracks forming [
38,
40,
69,
70,
72]. Further lithological studies are being carried out on new tracksites, including the sauropod tracksite of the Duntulm Formation, which may help elucidate the life environment of the trackmakers [
39]. The general environment appears to show a series of lagoons with some footprints in the Duntulm and Kilmaluag formations on rippled surfaces indicating that the animals walked on a higher energy sandy lagoon shoreline as well as the muddier low energy environments [
38,
40]. The sauropod prints in the Duntulm Formation were described as being made by animals wading in very shallow lagoonal water [
39].
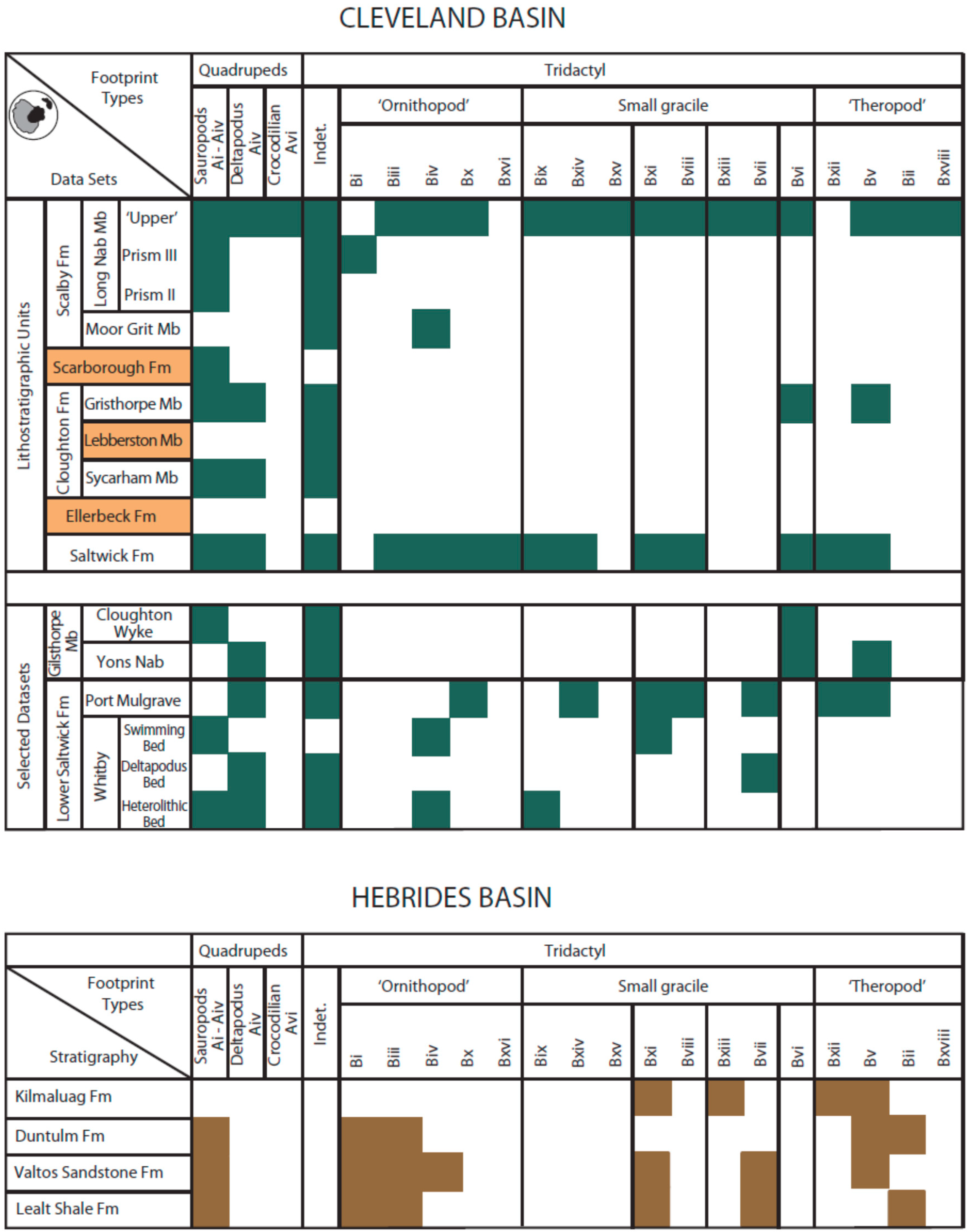
The distribution of prints throughout the stratigraphic range is more difficult to quantify. Their distribution in the Cleveland Basin (
Figure 5) is based on concentrated work in the area over the past 20 years or so, but is influenced by exposure and accessibility, as well as spatial and temporal heterogeneity within the succession [
77]. Thus, the absence of prints from a particular horizon may be due in part to a sampling deficiency or a result of behavioural or environmental control. The former category almost certainly explains the general lack of recorded prints from the Cloughton Formation, as well as the relatively high numbers of prints from the relatively accessible Saltwick Formation and Long Nab Member of the Scalby Formation. The distribution of print types shown as pie charts for the Scalby Formation on the foreshore of Scalby Bay [
77], where prints have been recorded within the top 1–2 m of the unit, show a degree of heterogeneity that may be real rather than a purely preservational bias.
The vertical distribution of prints in the Hebrides Basin (
Figure 5) is also difficult to quantify as there are only a few recorded sites thus far and most tracks are found on loose blocks on the foreshore below high vertical cliffs or eroding out of old landslips. For example, there is only one footprint recorded from the Lealt Shale Formation found near Brothers’ Point thus far [
68] and the stretch of coastline between Brothers’ Point and Dun Dearg is where most of the footprints of the Valtos Sandstone Formation have been found in loose blocks [
47,
72]. The spatial distribution of the footprints in the Valtos Sandstone Formation is difficult to assess as no precise horizon for the dinosaur remains has yet been established although the greatest variety of print types are found in this formation. Two localities are currently known for in situ Duntulm Formation prints: an Corran near Staffin from which the larger tridactyl prints are found; and the sauropod footprints of Cairidh Ghlumaig near Duntulm Castle. Both these localities are described as essentially monospecific although a poorly preserved tridactyl footprint was found at Cairidh Ghlumaig [
39] and an unidentified single small round footprint was found at an Corran [
70]. Only tridactyl footprints were found at Score Bay in the Kilmaluag Formation all of which may have been formed by one trackmaker species as they were indistinguishable based on their morphometrics using landmark analysis [
40]. A few footprints were found in situ at this locality. Precise correlation between localities are also quite difficult on the Isle of Skye as the sediments have been disrupted by multiple injections of Palaeogene igneous intrusions that jump between layers. This coupled with the discontinuity of layers, footprints being found mostly on loose blocks and lack of good continuous exposure make it impossible to be certain as to which precise horizon many of the prints derived from.
8. Animal Makers of Dinosaur Prints from the Cleveland and Hebrides Basins
Although it may be a primary objective to identify the dinosaur responsible for those making the prints, in reality, this is perhaps the most difficult, and often unsolvable part of the study of vertebrate palaeoichnology. Indeed, ‘tracks have a relatively low taxonomic resolution and are usually not attributable below the family level’ [
82]. Unlike similar studies in invertebrate palaeoichnology, where a number of examples are known where the maker is preserved intimately associated with the prints (
Kouphichnium and limulid [
83],
Rusophycus and trilobite [
84],
Lockeia and bivalve [
85]), such instances have never, as far as the authors are aware, been documented in the dinosaur ichnological record (other than the possible protoceratopsid with tracks from Mongolia [
86]). However, by studying well-preserved elite footprints [
87], together with unambiguous knowledge of dinosaur foot morphology, it is theoretically possible to confidently suggest possible print makers. That this should then be taken further and assigned a name to the footprint that connects it to its presumed maker is not a practice we would advocate. For example, prints originally described and named
Iguanodonichnus frenkii and
Neosauropus lagosteiriensis have been subsequently reinterpreted as being of sauropod and bipedal dinosaur origin, respectively [
4,
88,
89,
90].
However, there are some print types from the Cleveland Basin from which it is quite feasible to suggest their makers (at least to family level) with considerable confidence. Among these are the large prints (up to 1 m or more across) subspherical to oval in outline and with generally small, and sometimes curved up to five digit impressions. This group of prints, represented by Ai-Aiv in Group A (
Figure 3) belong to sauropod makers, and the markedly different forms of these in the Cleveland Basin suggest that at least two and possibly three different sauropod types inhabited Yorkshire during the Middle Jurassic. Print type Ai (
Brontopodus ichnosp., and Aii and Aiii (
Breviparopus-like ichnosp.) (
Figure 4) have been taken to indicate the presence of brachiosaur sauropods for the former [
73,
89], and a non-brachiosaurid [
89] such as
Cetiosaurus for the latter [
77].
The final print in Group A (
Figure 4, Av),
Deltapodus brodricki, has had a more controversial history in terms of its likely maker. Originally, it was tentatively thought to have been made by a sauropod [
60,
61] but was subsequently reinterpreted as possibly having a stegosaur maker [
62].
Apart from the above sauropod and stegosaur suggestions, it is extremely difficult to assign specific makers to individual print types. However, a provisional classification into ‘Ornithopoda’ or ‘Theropoda’ may be attempted for the print types in Group B. This was attempted by Whyte and others [
77], where five ‘ornithopods’ (Bi, Biii, Biv, Bx, Bxvi) and four ‘theropods’ (Bii, Bv, Bxii, BBxviii) were recognized. In addition, eight further prints (Bvi-ix, Bxi, Bxiii-xv) were classified as ‘small gracile’ prints. The two slender gracile prints from the Cleveland Basin (
Figure 4, Bxiv, Bxv) are particularly distinctive. Their wide divarication (105° and 145°, respectively [
18] is more reminiscent of birds, and indeed Currie [
91] maintained that the ‘divarification between digits II and IV in even the smallest dinosaurs never exceeds 100° on an average per trackway’. This apparent anomaly suggests either that birds were around as early as the Middle Jurassic, or (more likely) that some small dinosaurs (small adults or juveniles) did indeed exhibit such large divarication in the digits of their pes. The two print types Bxiv and Bxv (
Figure 4) resemble some traces assigned to
Trisauropodiscus, but the Cleveland Basin specimens lack the hallux imprint and, in the case of Bxv, show a wider divarication of digits II and IV [
92].
In the final group of morphotypes, Group C (
Figure 4), it is normally not possible to suggest the dinosaur makers, since these prints are purely the result of a behavioural (swimming) pattern that does not necessarily reflect the morphology of the foot, except in the extreme case of recording the number of digits (or at least those that made contact with the substrate), the divarification of the digits and whether the digit ended in claws or rounded terminations. However, two examples from the Cleveland Basin have been published where a swimming trace has been connected to walking prints that reflect the shape of the maker’s foot. One of these, the least certain, was described by Whyte and Romano [
63] when describing a dinosaur ichnocoenosis that showed walking, running and swimming prints. In this work, they suggested that the animal that made a walking trackway (Trackway C) was also responsible for the swimming Trackway B (
Figure 6 [
18,
63]), represented by Ci in
Figure 4. The second example is of a swimming track made by the same (type of) animal that made associated tracks of
Deltapodus brodricki. Since
D. brodricki is considered to have been made by a stegosaur (see above [
62]), the described swimming trackway (track Cii in
Figure 4, and also Ciii) that showed a number of features that connected it to the walking tracks provided suggestive evidence that stegosaurs could swim [
79].
The small prints (<20 cm length) from the Valtos Sandstone and Kilmaluag formations of the Hebrides Basin have been provisionally assigned to small theropod dinosaurs. The larger footprints from the Valtos Sandstone Formation are interpreted as having been made by a large ornithopod (
Table 1. PCA group Z), while the third group of large tridactyl tracks from the Duntulm Formation are regarded as large theropod tracks (
Table 1. PCA group Y) [
68,
71,
72]. The large track from the Valtos Sandstone Formation may be a large form of (
Table 1. PCA group Y), but the preservation was not good enough to provide reliable data. The sauropod prints recently described from the Duntulm Formation were assigned to a non-neosauropod maker [
39].
9. Palaeobiogeographical Aspects of the Dinosaur Communities Based on both Body- and Ichno-Fossil Evidence
It is proposed that similarities between the dinosaur ichnofaunas of the Cleveland and Hebrides basins may indicate that interchange of faunas took place between the two areas during the Jurassic. Whether this took place by chance or was the result of intentional migrations as a result of climatic pressures or searching for new food on during seasonal changes is not possible to speculate. However, certainly, the distances involved would not be excessive, since modern day wildebeest and zebra migrate over 3000 km each year in search of rain-ripened grass. Caribou migrate about 700 km from their wintering grounds to their calving grounds [
93] and can accumulate up to more than 5000 km in a year [
94] for the round journey.
From the diverse ichnofaunas of the Cleveland and Hebrides basins during the Middle Jurassic, it is apparent that both areas supported rich and varied dinosaur communities within this brief period of time (c. 10 Ma for the Cleveland Basin, and less for the Hebrides Basin). The former has evidence of brachiosaur dinosaurs and non-brachiosaur ‘
Cetiosaurus–type’ dinosaurs, from the Aalenian and Bathonian (and by inference from the Bajocian as well). These giant quadrupeds, which are represented by at least two and possibly three genera, invaded the Cleveland area during the earliest Middle Jurassic and were a dominant part of the community until persistent marine conditions returned in the Callovian. In the Hebrides Basin, sauropod remains are first recorded in the Valtos Sandstone Formation (Bathonian) where a footprint, (as well as bones and a tooth) have been found of a primitive eusauropod [
45,
47]. Further footprints and trackways of a primitive, non-neosauropod species have been found in the Duntulm Formation from the Trotternish Peninsula and represent the most northerly Middle Jurassic sauropod tracksite currently known [
39]. The last occurrence of sauropod remains being from the Kilmaluag Formation in Strathaird, southern Skye where a peg-like tooth was found that could belong to either a basal eusauropod or a basal titanosauriform [
48]. Sauropod prints have not yet been recorded from this unit. It is possible that these represent three different sauropod types on the Isle of Skye in the Middle Jurassic, and are thus comparable in diversity to that of the Cleveland Basin. Their presence first in the Cleveland Basin may suggest that they later spread to the Hebrides Basin. Whether this took place over the ‘highland’ of the Scottish and Pennine landmasses separating the two depocentres, or by way of the marginal alluvial plains and deltas (
Figure 2) is not possible to say. Certainly, the ‘highlands’ do not appear to have been a major physical barrier, since the general lack of much coarse-grained sediment at least in the Cleveland Basin indicates a rather subdued topography (see
Section 2). The abundance of plant remains [
24], both as in situ erect stems (and rootlet beds) and isolated drifted leaves and trunks [
5], indicates a rich source of plant food, both on the alluvial plain and in the hinterland [
95]. Thus, migrations from the Cleveland to Hebrides area could have taken place either around the margins of the land masses or over their tops. It has been suggested that the long legs of sauropods aided long distance movement [
1]. If this was the case, then the journey from the Cleveland Basin to the Hebrides Basin would appear to be quite achievable.
An apparent anomaly is the relatively common occurrence of possible stegosaur prints (
Deltapodus) from the Cleveland Basin, and their apparent absence from the currently described track record of the Hebrides Basin.
Deltapodus tracks are first recorded from the Aalenian of the Cleveland Basin, and subsequently occur in Upper Jurassic and Lower Cretaceous on a world-wide scale (see
Section 5 above). Whether they occur in the Hebrides Basin and have not yet been recognized, or if indeed they are not present in the latter is not possible to determine at this time. Despite the lack of footprints of stegosaurs from the Hebrides Basin, there is the proximal ulna and radius of a thyreophoran that may be an early stegosaur or ankylosaur [
51] as well as other undescribed bones that may belong to this group also. However, it still remains difficult to account for the absence of
Deltapodus tracks in the Hebrides Basin. In addition, even if such tracks are eventually identified, they may have been rare because they have yet to be found in the same sites where so many sauropod and tridactyl dinosaur (theropod and ornithopod) tracks have been found. Even if stegosaurs (the suggested
Deltapodus maker) were not physically suited to making excursions inland, away from the more flat-lying alluvial plains, they presumably could have explored the routes around the margins of the landmasses which were probably nearly continuous from Mid-Late Bathonian times (
Figure 2C,D). The presence of river systems along such routes should not have been an obstacle to stegosaur movement since it has been shown recently that stegosaurs were probably capable of swimming [
79].
Among the tridactyl tracks, the largest theropod tracks (Bxviii) from the Cleveland Basin are rare, with only two described examples [
76,
77]. Slightly smaller tracks made by theropods are represented by morphotypes Bii, Bv and Bxii (
Figure 4) [
77]. Two size ranges of prints made by ornithopods have also been identified; those of medium size (Bi, Biii, Biv, Bx, Bxvi), and those of small (<20 cm) gracile prints (Bvi-viii, xi, xiii-xv, ix) [
77]. In the Hebrides Basin, close comparisons may be made with some of the Cleveland Basin morphotypes (
Table 1), and broadly similar groups are represented, although on the whole diversity is less. This suggests that the food chains in the two areas are comparable (see
Section 9) and may have arisen from either the interchange of faunas from the two basins, or the (continuous or intermittent) influx of faunas from one basin to the other. In this scenario, we suggest that, since the Yorkshire Basin was established and colonized first, it was possible that initial migrations at first were from the Cleveland Basin to the Hebrides Basin.
While the above discussions are centered on the possibility of faunal exchange between the Cleveland and Hebrides basins during the Middle Jurassic, it cannot be discounted that animals also made their way into the Hebrides Basin from the ‘west’, from the landmass of Laurasia. In addition, of course, it still leaves unresolved the problem where the animals came from initially to colonise the slightly older Cleveland Basin. Most of the basins around Scotland at that time (until the Callovian) had shallow lagoonal and river deposits rather than open marine with deep channels, so it is equally possible that the dinosaurs came from North America across to the UK area via Greenland. The Bathonian dinosaur footprints from Wyoming are also indistinguishable from the footprints of the Kilmaluag Formation on Skye based on comparisons between landmark data [
72]. Sediment appears to have been derived from a southeasterly source during the deposition of the Great Estuarine Group in the Hebrides Basin, although it seems mostly to have been locally derived from the Scottish mainland [
96].
10. Structure of the Middle Jurassic Terrestrial Dinosaur Ecosystems from the Cleveland and Hebrides Basins as Deduced from Their Prints—A Tentative Food Chain
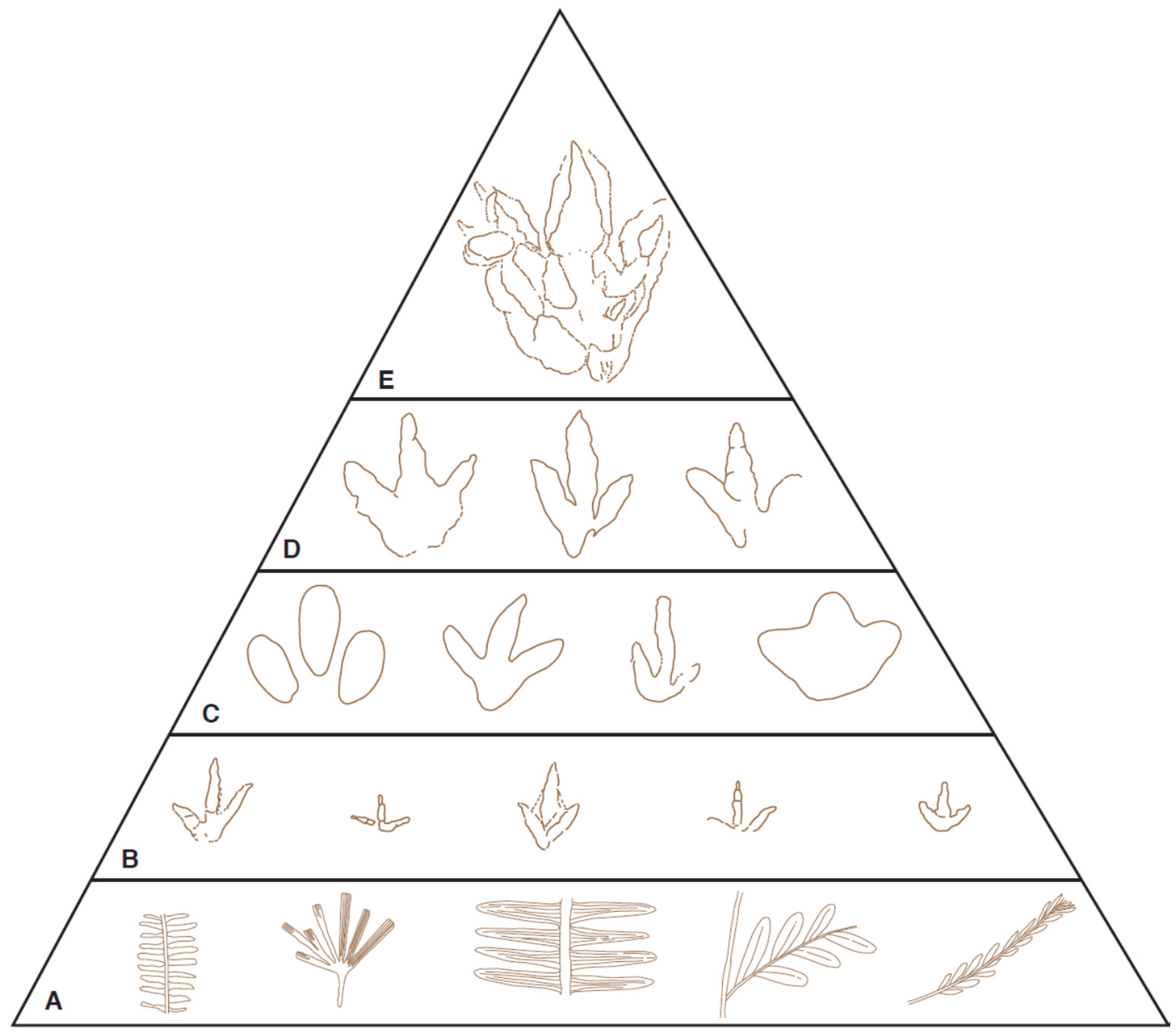
Finally, we attempt to portray visually the structure of the dinosaur ecosystems of the two basins during the Middle Jurassic. This is based on the print types and their relative abundance within the community. A visual representation has previously been attempted for part of a beach section within the Scalby Formation of the Cleveland Basin [
77], where the abundances of three groups (large tridactyl, small tridactyl and sauropod) were based on the relative numbers of their prints. While we categorically accept that this approach is not based on a firm statistical basis, and is heavily dependent on collector’s bias and exposure, we feel that a visual portrayal has some benefit in trying to reconstruct the past community based on our current level of understanding (and with the recognition that such visual portrayals can be easily modified with new discoveries). We here propose a broadly similar visual approach, based on the prints recorded within the sequences, except that the diagram is in the form of a food chain that shows how food energy moves from one organism to another (where one animal eats another animal that is lower in the food chain) in a given environment. At each level of the food chain, print examples are known from both the Cleveland and Hebrides basins.
The diagram (
Figure 6) starts at the base of the food chain with a selected sample of the diverse flora that existed at the time. Over 250 plant species have been recorded from the Yorkshire Basin, representing species that lived on the alluvial plain or in the adjacent hinterland. These belong to conifers, ferns, bryophytes, lycopsids, sphenophytes, pteridosperms, Caytoniales, Cycadales, Ginkgoales and Bennettitales [
24,
95,
97,
98], while allochthonous floras have been recorded from Skye [
99]. The next layer in the food chain may be represented by the small gracile forms, both herbivorous ornithischians (not illustrated in
Figure 6) feeding on this flora and carnivorous theropods. The latter would be feeding on small prey that includes the diverse invertebrate fauna (not represented on the diagram) that was likely present at that time, such as arthropods, bivalves, annelids, as well as fish, pond tortoises [
48], small ornithischians, and perhaps cadavers that were the result of natural death or the remains of a kill from the larger theropods. Medium to large size ornithischians (including ornithopods and stegosaurs) would occupy the next level of the food chain. This group of animals, probably existing in herds, would generally not be troubled by small carnivores, and could browse on the higher vegetation of leaves and young shoots not available to their smaller relatives. The final two layers in the food chain were the domain of the large carnivores. Whether these animals were primarily hunters, scavengers or both may remain open to question, but their position at the top of the food chain is without doubt. It is interesting to note that the largest theropod prints found are very rare. Only two clear examples are known from the Cleveland Basin, identified as morphotype Bxviii [
76,
77], and were tentatively assigned to a
Megalosaurus maker [
76]. In the Hebrides Basin, theropod tracks over 40 cm in length are known from the Duntulm Formation, and a recently found tridactyl print from the Valtos Sandstone Formation is over 1 m in length (although it is unclear if this was made by a theropod or an ornithopod). The scarcity of top predator, as evidenced by their meager print record, organisms at the top of the food chain is well shown in the dinosaur print record of the Cleveland Basin where the two prints of these large carnivores contrasts with nearly 40 of a small bipedal gracile form such as Bix [
77].
The two remaining groups not incorporated into the food chain are those represented by the morphotypes in Group A (
Figure 4); the sauropods (Ai-iv) and stegosaurs (Av). Sauropod food requirements of these giant herbivores ‘must have been impressive’ [
84], but they presumably found sufficient food in the abundant and diverse plant life that colonized the levees of the river systems in the alluvial plains. The food chain of these animals is much reduced compared with that described above. Although it is likely that carnivores repeatedly hunted small sauropods, it is equally likely that their success was limited when confronted by the enormous body size of an adult sauropod [
96]. Whether there was competition for food between the brachiosaur dinosaurs and non-brachiosaur ‘
Cetiosaurus-type’ dinosaurs in the Cleveland Basin [
5,
65], or the eusauropods and Cetiosaurid in the Hebrides Basin [
39,
45,
47] is not possible to tell. However, from the print evidence of these giant animals in the Cleveland Basin, they coexisted from the Aalenian to Bathonian (
Figure 5).
Whatever the actual food requirements were for these morphologically diverse Middle Jurassic herbivores, the range of plant food available both on the flood plains and in the hinterland was exceedingly diverse as can be deduced from studies of plant remains and spore-pollen assemblages [
24,
95,
97,
98,
100].
A similarly restricted food chain must have existed for the stegosaurs. As with the sauropods, stegosaurs were herbivorous animals [
97], yet despite their considerable bulk, they were significantly smaller than the sauropods and did not possess an elongated neck nor long legs (particularly the anterior pair). Hence, they are generally considered to be low browsers [
95,
100], even if they were capable, as has been suggested, to rear up on their hind legs using the tail as a tripod to feed on leaves higher up in the trees [
101]. Large theropods may have preyed on stegosaurs, but the rows of dorsal plates and the spiny termination to their tail may have proved an effective deterrent.
11. Problems still to Be Resolved
Despite our knowledge of the dinosaur tracks and their probable makers within the Cleveland and Hebrides basins, there are still many outstanding problems to be resolved. We have virtually no evidence of social behaviour, such as herding and post-hatching care, from the footprints, but this is almost certainly the result of the lack of large bedding planes in the mainly coastal exposures. We also lack any evidence of eggs and nesting sites from either of the two basins., and only a single recording of tridactyl tracks of small/juvenile bipedal forms from the Cleveland Basin for one of the morphotypes (
Figure 4, Bix) [
77]. There are, however, juvenile and hatchling tracks in the Hebrides Basin in the Kilmaluag Formation [
40,
72] and also size variations in the footprints in the Valtos Sandstone Formation [
69,
71,
72]. The footprints on a single slab of the Kilmaluag Formation with the two track sizes may indicate that there was post-hatching care in theropods as all the tracks are pointing in the same direction [
40]. The slab, however, only shows about 24 footprints within the 100 cm × 175 cm area. The only non-sauropod quadruped maker for which juvenile prints have been recorded are for morphotype Av (
Figure 4) [
77] from the Cleveland Basin. Until now, no small/juvenile tracks have been recorded for sauropods from either the Cleveland area (
Figure 4, morphotypes Ai-Aiv) or Isle of Skye. It is tempting to explain the absence of the latter by proposing that nests were made in the drier upland areas adjacent to the two basins, and so that is where small (baby and juvenile) dinosaur tracks would have been made, before making their way as young adults to the coastal and alluvial plains. However, it is also relevant to appreciate that the general absence of very small tracks is not a preservational phenomenon, since delicate tracks made by small non-dinosaurian vertebrates and invertebrates are common in the fine-grained sedimentary rocks of the two basins [
8,
70]. More work is needed on these two classic areas before we can begin to unravel the full story.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}