Terraced Iron Formations: Biogeochemical Processes Contributing to Microbial Biomineralization and Microfossil Preservation

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
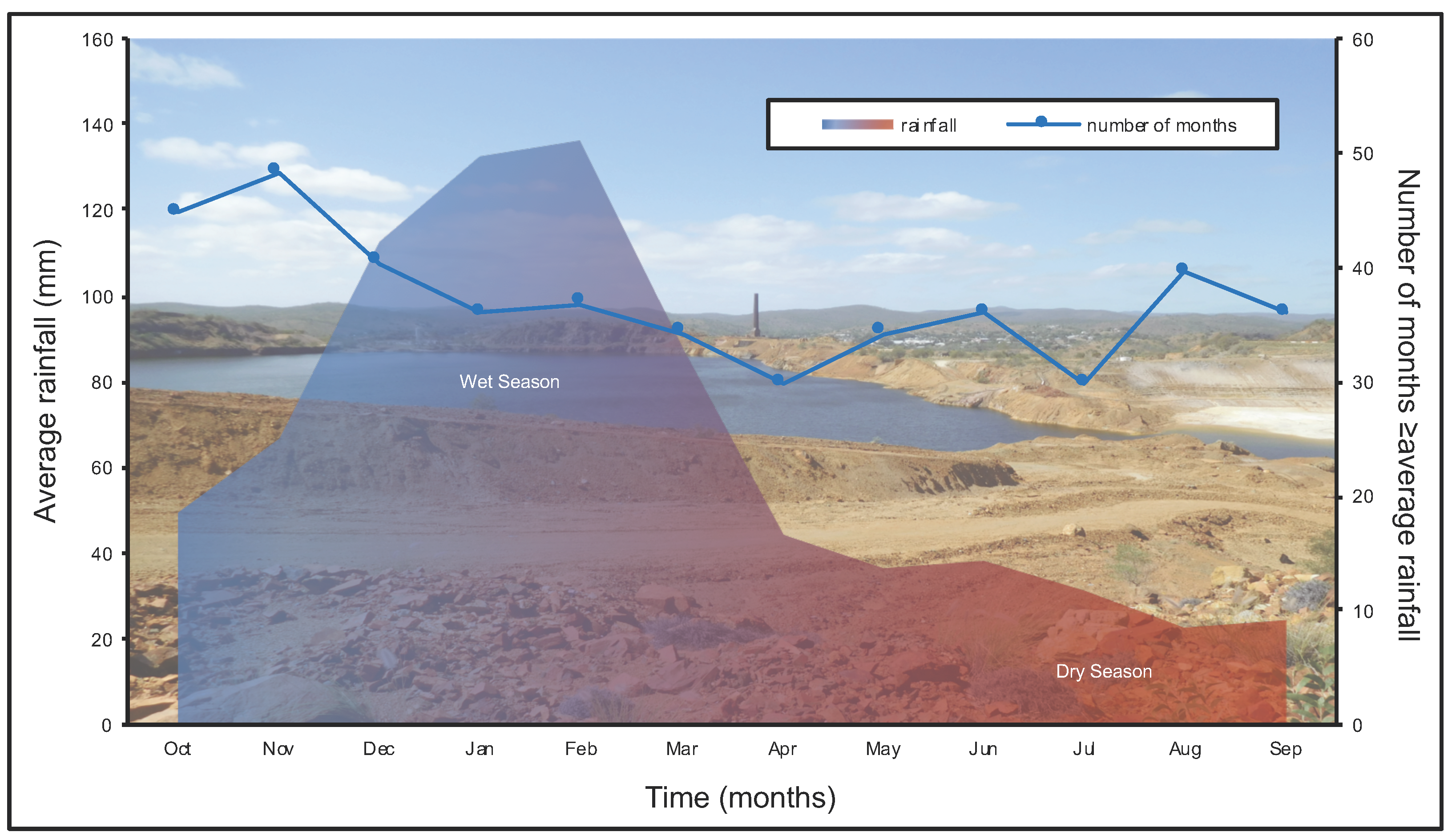
2.1. Field Site and Sampling Acquisition
2.2. Microbe and Mineral Characterization
2.2.1. Molecular Analysis
2.2.2. Iron-oxidizing Bacterial Enrichment
2.2.3. Electron Microscopy–Energy Dispersive Spectroscopy
2.3. TIF Chemistry, Mineralogy, and Structure
2.3.1. Inductively Coupled Plasma–Mass Spectroscopy
2.3.2. X-ray Diffraction
2.3.3. X-Ray Florescence Microscopy
2.3.4. Solid Phase Scanning Electron Microscopy–Energy Dispersive Spectroscopy
3. Results
3.1. Microbial Diversity
3.2. Iron-Oxidizing Bacterial Enrichment
3.3. Structure and Chemistry of Terrace Iron Formations
4. Discussion
4.1. Interpreting Biogeochemical Processes Contributing to TIF Development
4.2. Estimating the Kinetics of TIF Development
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- España, J.S.; Pamo, E.L.; Pastor, E.S.; Andrés, J.R.; Rubí, J.A.M. The natural attenuation of two acidic effluents in Tharsis and La Zarza-Perrunal mines (Iberian Pyrite Belt, Huelva, Spain). Environ. Geol. 2005, 49, 253–266. [Google Scholar] [CrossRef]
- Sánchez España, J.; Santofimia Pastor, E.; López Pamo, E. Iron terraces in acid mine drainage systems: A discussion about the organic and inorganic factors involved in their formation through observations from the Tintillo acidic river (Riotinto mine, Huelva, Spain). Geosphere 2007, 3, 133–151. [Google Scholar] [CrossRef]
- Brown, J.F.; Jones, D.S.; Mills, D.B.; Macalady, J.L.; Burgos, W.D. Application of a depositional facies model to an acid mine drainage site. Appl. Environ. Microbiol. 2011, 77, 545–554. [Google Scholar] [CrossRef] [PubMed]
- Sánchez España, J.; López Pamo, E.; Santofimia, E.; Aduvire, O.; Reyes, J.; Barettino, D. Acid mine drainage in the Iberian Pyrite Belt (Odiel river watershed, Huelva, SW Spain): Geochemistry, mineralogy and environmental implications. Appl. Geochem. 2005, 20, 1320–1356. [Google Scholar] [CrossRef]
- Jones, D.S.; Kohl, C.; Grettenberger, C.; Larson, L.N.; Burgos, W.D.; Macaladya, J.L. Geochemical niches of iron-oxidizing acidophiles in acidic coal mine drainage. Appl. Environ. Microbiol. 2015, 81, 1242–1250. [Google Scholar] [CrossRef]
- Jones, R.A.; Koval, S.F.; Nesbitt, H.W. Surface alteration of arsenopyrite (FeAsS) by Thiobacillus ferrooxidans. Geochim. Cosmochim. Acta 2003, 67, 955–965. [Google Scholar] [CrossRef]
- Bigham, J.M.; Schwertmann, U.; Carlson, L.; Murad, E. A poorly crystallized oxyhydroxysulfate of iron formed by bacterial oxidation of Fe(II) in acid mine waters. Geochim. Cosmochim. Acta 1990, 54, 2743–2758. [Google Scholar] [CrossRef]
- Bigham, J.M.; Schwertmann, U.; Traina, S.J.; Winland, R.L.; Wolf, M. Schwertmannite and the chemical modeling of iron in acid sulfate waters. Geochim. Cosmochim. Acta 1996, 60, 2111–2121. [Google Scholar] [CrossRef]
- Nordstrom, D.K. Aqueous pyrite oxidation and the consequent formation of secondary iron minerals. In Acid Sulfate Weathering; Kittrick, J.A., Fanning, D.S., Hosner, L.R., Eds.; Soil Science Society of America: Madison, WI, USA, 1982; pp. 37–56. [Google Scholar]
- Nordstrom, D.K.; Southam, G. Geomicrobiology of sulphide mineral oxidation. Rev. Mineral. 1997, 35, 362–390. [Google Scholar]
- Sasaki, K.; Sakimoto, T.; Endo, M.; Konno, H. FE-SEM study of microbially formed jarosite by Acidithiobacillus Ferrooxidans. Mater. Trans. 2006, 47, 1155–1162. [Google Scholar] [CrossRef]
- Sánchez-España, J.; Yusta, I.; Gray, J.; Burgos, W.D. Geochemistry of dissolved aluminum at low pH: Extent and significance of Al–Fe(III) coprecipitation below pH 4.0. Geochim. Cosmochim. Acta 2016, 175, 128–149. [Google Scholar] [CrossRef]
- Brake, S.S.; Hasiotis, S.T.; Dannelly, H.K. Diatoms in Acid Mine Drainage and Their Role in the Formation of Iron-Rich Stromatolites. Geomicrobiol. J. 2004, 21, 331–340. [Google Scholar] [CrossRef]
- Fernández-Remolar, D.C.; Knoll, A.H. Fossilization potential of iron-bearing minerals in acidic environments of Rio Tinto, Spain: Implications for Mars exploration. Icarus 2008, 194, 72–85. [Google Scholar] [CrossRef]
- Sasaki, K.; Tsunekawa, M.; Konno, H. Characterisation of argentojarosite formed from biologically oxidised Fe3+ ions. Can. Mineral. 1995, 33, 1311–1319. [Google Scholar]
- Groat, L.A.; Jambor, J.L.; Pemberton, B.C. The crystal structure of argentojarosite. Can. Miner. 2003, 41, 921–928. [Google Scholar] [CrossRef]
- Shuster, J.; Reith, F.; Izawa, M.; Flemming, R.; Banerjee, N.; Southam, G. Biogeochemical Cycling of Silver in Acidic, Weathering Environments. Minerals 2017, 7, 218. [Google Scholar] [CrossRef]
- Shuster, J.; Bolin, T.; MacLean, L.C.W.; Southam, G. The effect of iron-oxidising bacteria on the stability of gold (I) thiosulfate complex. Chem. Geol. 2014, 376, 52–60. [Google Scholar] [CrossRef]
- Ferris, F.; Hallbeck, L.; Kennedy, C.; Pedersen, K. Geochemistry of acidic Rio Tinto headwaters and role of bacteria in solid phase metal partitioning. Chem. Geol. 2004, 212, 291–300. [Google Scholar] [CrossRef]
- Levett, A.; Gagen, E.; Shuster, J.; Rintoul, L.; Tobin, M.; Vongsvivut, J.; Bambery, K.; Vasconcelos, P.; Southam, G. Evidence of biogeochemical processes in iron duricrust formation. J. S. Am. Earth Sci. 2016, 71, 131–142. [Google Scholar] [CrossRef]
- Preston, L.J.; Shuster, J.; Fernandez-Remolar, D.; Banerjee, N.R.; Osinski, G.R.; Southam, G. The preservation and degradation of filamentous bacteria and biomolecules within iron oxide deposits at Rio Tinto, Spain. Geobiology 2011, 9, 233–249. [Google Scholar] [CrossRef]
- Chi Fru, E.; Ivarsson, M.; Kilias, S.P.; Bengtson, S.; Belivanova, V.; Marone, F.; Fortin, D.; Broman, C.; Stampanoni, M. Fossilized iron bacteria reveal a pathway to the biological origin of banded iron formation. Nat. Commun. 2013, 4, 2050. [Google Scholar] [CrossRef]
- Edraki, M.; Golding, S.D.; Baublys, K.A.; Lawrence, M.G. Hydrochemistry, mineralogy and sulfur isotope geochemistry of acid mine drainage at the Mt. Morgan mine environment, Queensland, Australia. Appl. Geochem. 2005, 20, 789–805. [Google Scholar] [CrossRef]
- Lane, D.J. 16S:23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Osborn, A.M.; Moore, E.R.B.; Timmis, K.N. An evaluation of terminal-restriction fragment length polymorphism (T-RFLP) analysis for the study of microbial community structure and dynamics. Environ. Microbiol. 2000, 2, 39–50. [Google Scholar] [CrossRef] [PubMed]
- Reith, F.; Fairbrother, L.; Nolze, G.; Wilhelmi, O.; Clode, P.L.; Gregg, A.; Parsons, J.E.; Wakelin, S.A.; Pring, A.; Hough, R.; et al. Nanoparticle factories: Biofilms hold the key to gold dispersion and nugget formation. Geology 2010, 38, 843–846. [Google Scholar] [CrossRef]
- Bissett, A.; Fitzgerald, A.; Court, L.; Meintjes, T.; Mele, P.M.; Reith, F.; Dennis, P.G.; Breed, M.F.; Brown, B.; Brown, M.V.; et al. Introducing BASE: The Biomes of Australian Soil Environments soil microbial diversity database. Gigascience 2016, 5, 21. [Google Scholar] [CrossRef]
- Lane, D.J.; Pace, B.; Olsen, G.J.; Stahl, D.A.; Sogin, M.L.; Pace, N.R. Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc. Natl. Acad. Sci. USA 1985, 82, 6955–6959. [Google Scholar] [CrossRef]
- Silverman, M.P.; Lundgren, D.G. Studies on the chemoautotrophic iron bacterium Ferrobacillus Ferrooxidans. J. Bacteriol. 1959, 77, 642–647. [Google Scholar]
- Sánchez-España, J.; Yusta, I.; López, G.A. Schwertmannite to jarosite conversion in the water column of an acidic mine pit lake. Mineral. Mag. 2012, 76, 2659–2682. [Google Scholar] [CrossRef]
- Paterson, D.; de Jonge, M.D.; Howard, D.L.; Lewis, W.; McKinlay, J.; Starritt, A.; Kusel, M.; Ryan, C.G.; Kirkham, R.; Moorhead, G.; et al. The X-ray Fluorescence Microscopy Beamline at the Australian Synchrotron. In Proceedings of the AIP Conference, Chicago, IL, USA, 15–20 Aug 2010; pp. 219–222. [Google Scholar]
- CSIRO. GeoPIXE Software for SXRF Imaging. 2011. Available online: http://nmp.csiro.au/GeoPIXE.html (accessed on 7 October 2017).
- Li, K.; Etschmann, B.; Rae, N.; Reith, F.; Ryan, C.; Kirkham, R.; Howard, D.; Rosa, D.; Zammit, C.; Pring, A.; et al. Ore petrography using megapixel X-ray imaging: Rapid insights into element distribution and mobilization in complex Pt and U-Ge-Cu ores. Econ. Geol. 2016, 111, 487–501. [Google Scholar] [CrossRef]
- Bureau of Meteorology (Australian Government). Available online: www.bom.gov.au (accessed on 12 June 2018).
- Ehrlich, H.L. Bacterial oxidation of arsenopyrite and enargite. Econ. Geol. 1964, 59, 1306–1312. [Google Scholar] [CrossRef]
- Ehrlich, H.L. Microbes and metals. Appl. Microbiol. Biotechnol. 1997, 48, 687–692. [Google Scholar] [CrossRef]
- Gagen, E.J.; Levett, A.; Shuster, J.; Fortin, D.; Vasconcelos, P.M.; Southam, G. Microbial Diversity in Actively Forming Iron Oxides from Weathered Banded Iron Formation Systems. Microbes Environ. 2018. [Google Scholar] [CrossRef] [PubMed]
- Mazuelos, A.; Moreno, J.M.; Carranza, F.; Palomino, C.; Torres, A.; Villalobo, E. Biotic factor does not limit operational pH in packed-bed bioreactor for ferrous iron biooxidation. J. Ind. Microbiol. Biotechnol. 2012, 39, 1851–1858. [Google Scholar] [CrossRef] [PubMed]
- Ueoka, N.; Kouzuma, A.; Watanabe, K. Missing Iron-Oxidizing Acidophiles Highly Sensitive to Organic Compounds. Microbes Environ. 2016, 31, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Rowe, O.F.; Sanchez-Espana, J.; Hallberg, K.B.; Johnson, D.B. Microbial communities and geochemical dynamics in an extremely acidic, metal-rich stream at an abandoned sulfide mine (Huelva, Spain) underpinned by two functional primary production systems. Environ. Microbiol. 2007, 9, 1761–1771. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Archilla, A.I.; Marin, I.; Amils, R. Microbial Community Composition and Ecology of an Acidic Aquatic Environment: The Tinto River, Spain. Microb. Ecol. 2001, 41, 20–35. [Google Scholar] [PubMed]
- Johnson, D.B.; Hallberg, K.B. The microbiology of acidic mine waters. Res. Microbiol. 2003, 154, 466–473. [Google Scholar] [CrossRef]
- Schippers, A.; Sand, W. Bacterial leaching of metal sulfides proceeds by two indirect mechanisms via thiosulfate or via polysulfides and sulfur. Appl. Environ. Microbiol. 1999, 65, 319–321. [Google Scholar]
- Bigham, J.M.; Jones, F.S.; Özkaya, B.; Sahinkaya, E.; Puhakka, J.A.; Tuovinen, O.H. Characterization of jarosites produced by chemical synthesis over a temperature gradient from 2 to 40 °C. Int. J. Miner. Process. 2010, 94, 121–128. [Google Scholar] [CrossRef]
- Capaldo-Kimball, F.; Barbour, S. Involvement of recombination gemes in growth and viability of Eschichia coli K-12. J. Bacteriol. 1971, 106, 204–212. [Google Scholar]
- Gasparon, M.; Smedley, A.; Costagliola, P.; Benvenuti, M. Acid mine drainage at mount Morgan, Queensland (Australia): Experimental simulation and geochemical modelling of buffering reactions. In Proceedings of the IMWA Symposium 2007: Water in Mining Environments, Cagliari, Italy, 27–31 May 2007; pp. 433–436. [Google Scholar]
- Westall, F. The nature of fossil bacteria: A guide to the search for extraterrestrial life. J. Geophys. Res. Planet 1999, 104, 16437–16451. [Google Scholar] [CrossRef]
- McCutcheon, J.; Southam, G. Advanced biofilm staining techniques for TEM and SEM in geomicrobiology: Implications for visualizing EPS architecture, mineral nucleation, and microfossil generation. Chem. Geol. 2018, 498, 115–127. [Google Scholar] [CrossRef]
- Chan, C.S.; McAllister, S.M.; Leavitt, A.H.; Glazer, B.T.; Krepski, S.T.; Emerson, D. The Architecture of Iron Microbial Mats Reflects the Adaptation of Chemolithotrophic Iron Oxidation in Freshwater and Marine Environments. Front. Microbiol. 2016, 7, 796. [Google Scholar] [CrossRef] [PubMed]
- Shuster, J.; Lengke, M.F.; Márquez-Zavalía, M.F.; Southam, G. Floating gold grains and nanophase particles produced from the biogeochemical weathering of a gold-bearing ore. Econ. Geol. 2016, 111, 1485–1494. [Google Scholar] [CrossRef]
- Levett, A.; Gagen, E.J.; Diao, H.; Guagliardo, P.; Rintoul, L.; Paz, A.; Vasconcelos, P.M.; Southam, G. The role of aluminium in the preservation of microbial biosignatures. Geosci. Front. 2018. [Google Scholar] [CrossRef]
- Olson, G.J. Rate of pyrite bioleaching by Thiobacillus ferrooxidans: Results of an interlaboratory comparison. Appl. Environ. Microbiol. 1991, 57, 642–644. [Google Scholar] [PubMed]
- Shuster, J.; Marsden, S.; Maclean, L.; Ball, J.; Bolin, T.; Southam, G. The immobilization of gold from gold (III) chloride by a halophilic sulfate-reducing bacterial consortium. Geol. Soc. Lond. Spec. Pub. 2013, 393, 249–263. [Google Scholar] [CrossRef]
- Shuster, J.; Southam, G. The in-vitro “growth” of gold grains. Geology 2014, 43, 79–82. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element (Detection Limit) | Average (Standard Deviation) | |
|---|---|---|
| B | (5.0 × 10−2) | <5.0 × 10−2 |
| Na | (2.0 × 10−1) | 7.92 × 102 (± 3.97 × 102) |
| Mg | (1.0 × 10−1) | 4.92 × 103 (± 9.17 × 101) |
| Al | (5.0 × 10−2) | 3.32 × 104 (± 7.37 × 102) |
| P | (2.0 × 10−1) | 8.94 × 102 (± 6.84 × 101) |
| S | (2.0 × 10−1) | 7.46 × 104 (± 4.08 × 103) |
| K | (2.0 × 10−1) | 1.13 × 102 (± 3.12 × 101) |
| Ca | (1.0 × 10−1) | 3.14 × 104 (± 8.14 × 102) |
| Cr | (5.0 × 10−2) | 6.47 × 101 (± 9.77 × 10−1) |
| Mn | (5.0 × 10−2) | 3.27 × 102 (± 5.08 × 101) |
| Fe | (1.0 × 10−1) | 2.78 × 105 (± 3.56 × 104) |
| Co | (5.0 × 10−2) | <5.0 × 10−2 |
| Ni | (5.0 × 10−2) | <5.0 × 10−2 |
| Cu | (5.0 × 10−2) | 2.28 × 101 (± 2.11 × 100) |
| Zn | (5.0 × 10−2) | 9.57 × 101 (± 7.91 × 10−1) |
| As | (5.0 × 10−2) | 1.29 × 101 (± 5.11 × 10−1) |
| Se | (5.0 × 10−2) | <5.0 × 10−2 |
| Mo | (5.0 × 10−2) | <5.0 × 10−2 |
| Ag | (2.0 × 10−1) | 6.00 × 10−2 |
| Cd | (5.0 × 10−2) | <5.0 × 10−2 |
| Sb | (1.0 × 10−1) | <1.0 × 10−1 |
| Au | (2.0 × 10−1) | 3.00 × 10−1 |
| Pb | (1.0 × 10−1) | 3.40 × 100 (± 3.00 × 10−1) |
| Fe Oxidation Rate (ppm L−1 year−1) A | Fe3+ as Precipitate (ppm L−1 year−1) B | Fe3+ per m2 (ppm year−1) C | Time (years) D |
|---|---|---|---|
| 9.72 × 105 | 1.42 × 103 | 1.05 × 106 | 0.25 |
| 1.07 × 105 | 1.56 × 102 | 1.16 × 105 | 2.28 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shuster, J.; Rea, M.A.; Etschmann, B.; Brugger, J.; Reith, F. Terraced Iron Formations: Biogeochemical Processes Contributing to Microbial Biomineralization and Microfossil Preservation. Geosciences 2018, 8, 480. https://doi.org/10.3390/geosciences8120480
Shuster J, Rea MA, Etschmann B, Brugger J, Reith F. Terraced Iron Formations: Biogeochemical Processes Contributing to Microbial Biomineralization and Microfossil Preservation. Geosciences. 2018; 8(12):480. https://doi.org/10.3390/geosciences8120480
Chicago/Turabian StyleShuster, Jeremiah, Maria Angelica Rea, Barbara Etschmann, Joël Brugger, and Frank Reith. 2018. "Terraced Iron Formations: Biogeochemical Processes Contributing to Microbial Biomineralization and Microfossil Preservation" Geosciences 8, no. 12: 480. https://doi.org/10.3390/geosciences8120480
APA StyleShuster, J., Rea, M. A., Etschmann, B., Brugger, J., & Reith, F. (2018). Terraced Iron Formations: Biogeochemical Processes Contributing to Microbial Biomineralization and Microfossil Preservation. Geosciences, 8(12), 480. https://doi.org/10.3390/geosciences8120480