Algal Bloom Exacerbates Hydrogen Sulfide and Methylmercury Contamination in the Emblematic High-Altitude Lake Titicaca

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
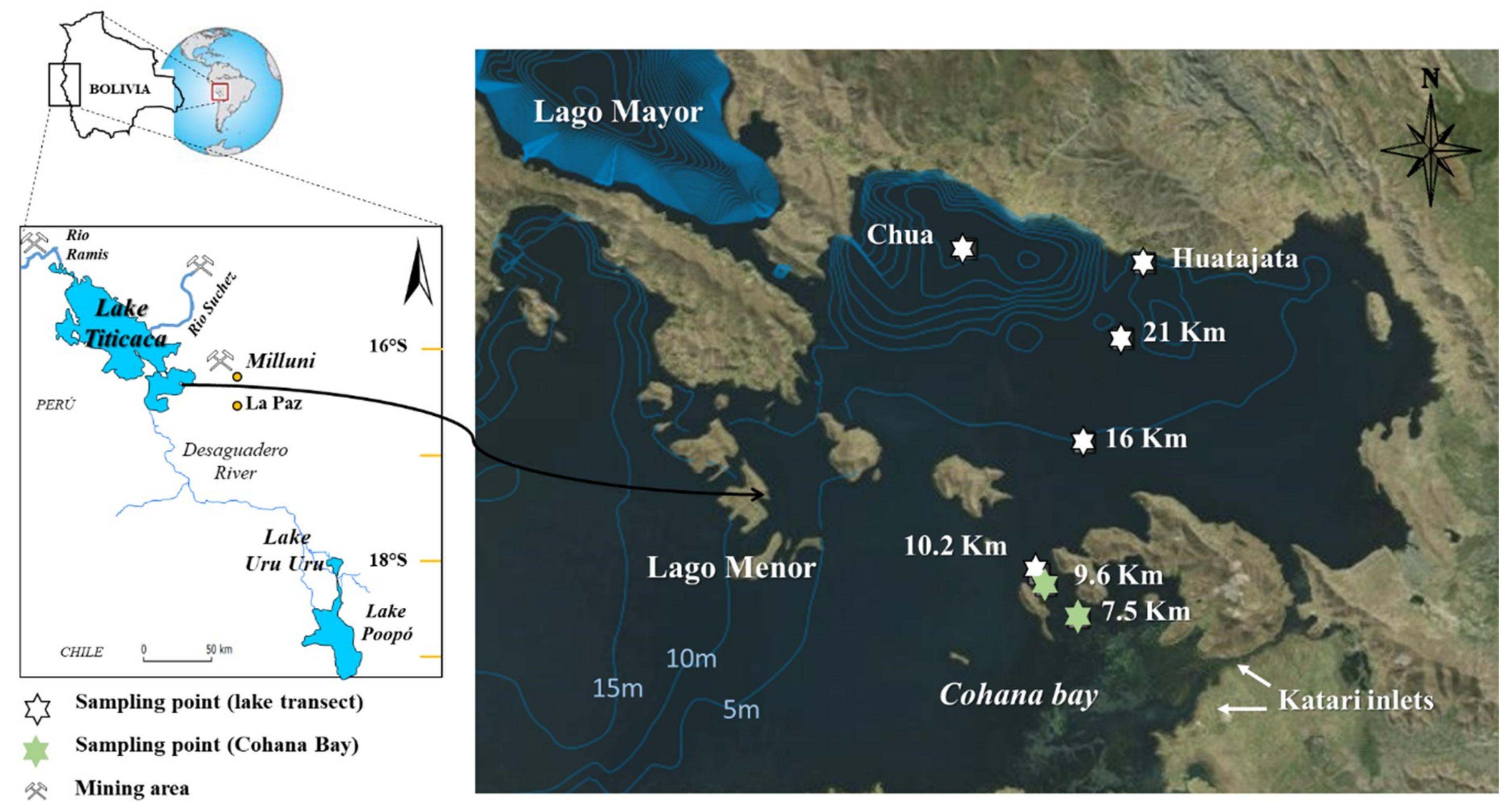
2.1. Study Area and Sampling
2.2. Hydrogen Sulfide
2.3. Mercury and Methylmercury
2.4. Dissolved Organic Carbon and Algae Identification
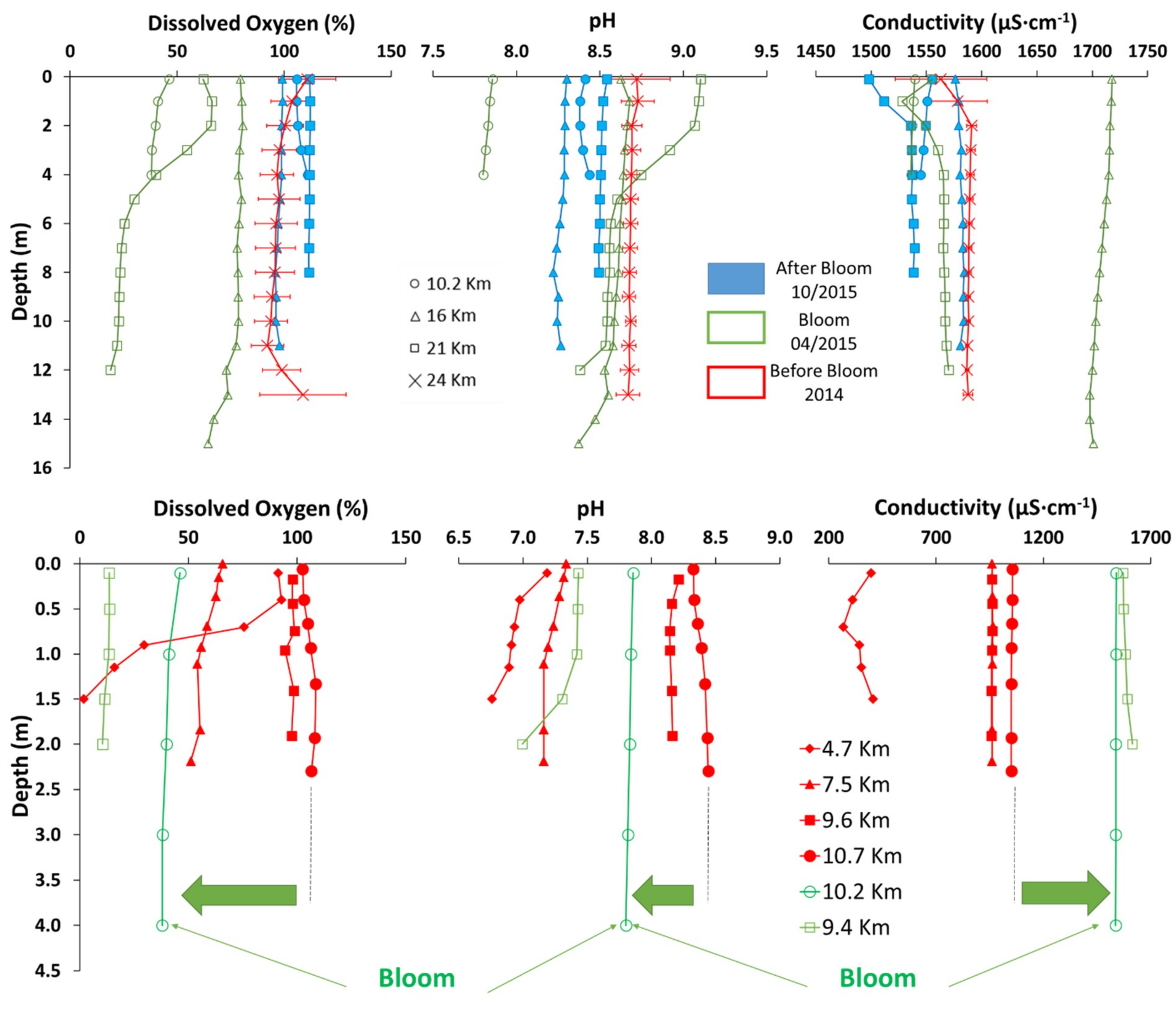
2.5. Water Column Characterization
2.6. Incubation Experiments
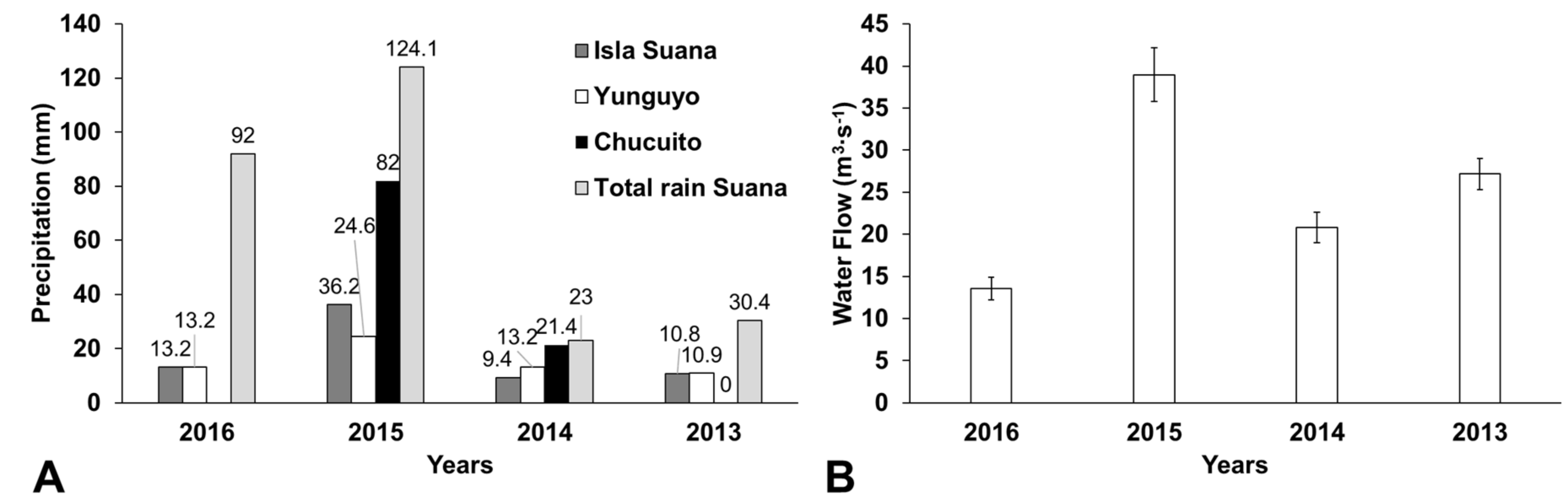
2.7. Climatological Data
2.8. Statistical Analysis
3. Results and Discussion
3.1. Algae Responsible for the Bloom and Consequences on Water Quality
3.2. The Trigger Mechanism of the Bloom and Its Impact on Water Column Quality
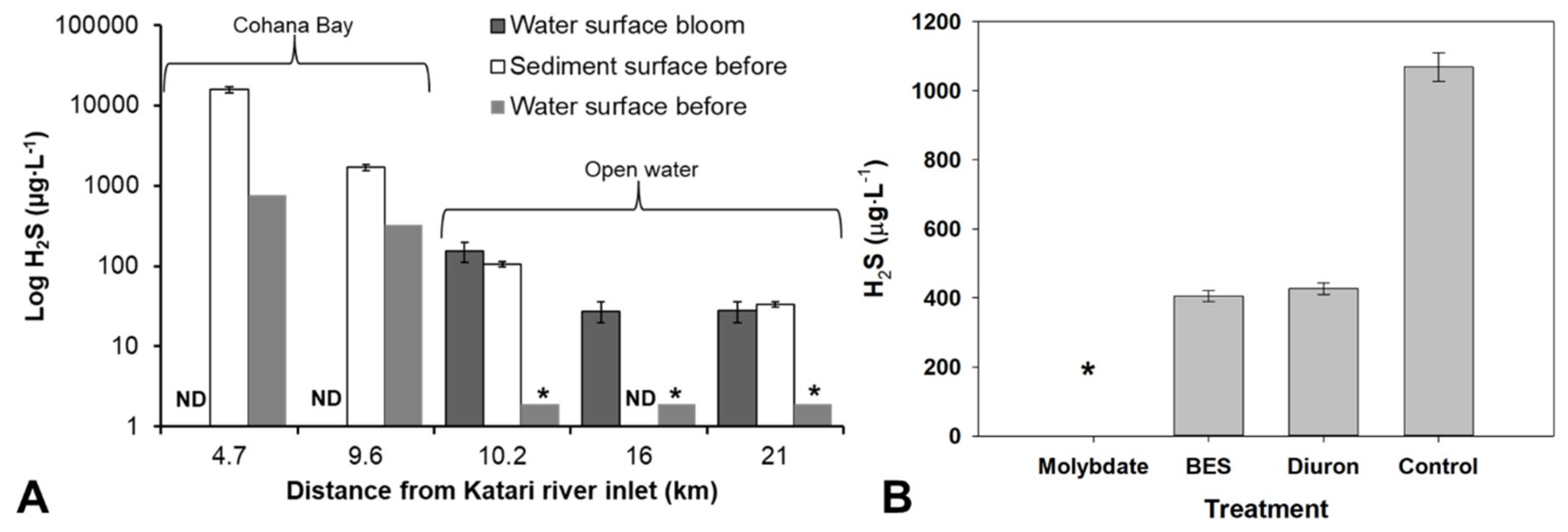
3.3. Algal Bloom Exacerbate the Production of H2S by Sulfate-Reducing Bacteria
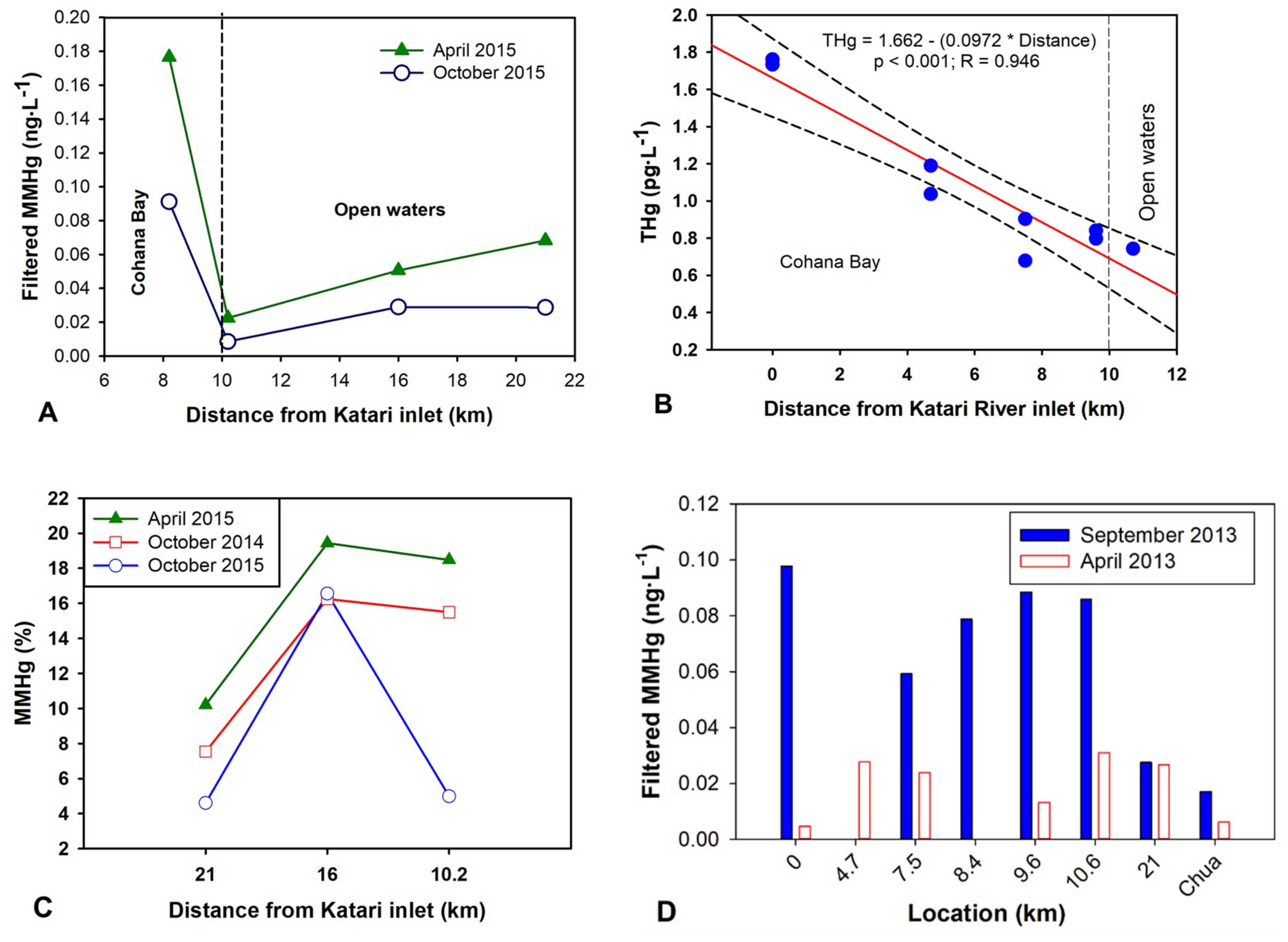
3.4. Enhanced Methylmercury Concentration in Surface Water during the Bloom
4. Conclusions and Perspectives
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, V.H. Eutrophication of freshwater and coastal marine ecosystems: A global problem. Environ. Sci. Pollut. Res. 2003, 10, 126–139. [Google Scholar] [CrossRef]
- Fey, S.B.; Siepielski, A.M.; Nusslé, S.; Cervantes-Yoshida, K.; Hwan, J.L.; Huber, E.R.; Fey, M.J.; Catenazzi, A.; Carlson, S.M. Recent shifts in the occurrence, cause, and magnitude of animal mass mortality events. Proc. Natl. Acad. Sci. USA 2015, 112, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Paerl, H.W.; Huisman, J. Blooms Like It Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Havens, K.E. Cyanobacteria blooms: Effects on aquatic ecosystems. In Cyanobacterial Harmful Algal Blooms: State of the Science and Research Needs; Hudnell, H.K., Ed.; Springer: New York, NY, USA, 2008; pp. 733–747. [Google Scholar]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful algal blooms and eutrophication: Nutrient sources, composition, and consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Pickhardt, P.C.; Folt, C.L.; Chen, C.Y.; Klaue, B.; Blum, J.D. Algal blooms reduce the uptake of toxic methylmercury in freshwater food webs. Proc. Natl. Acad. Sci. USA 2002, 99, 4419–4423. [Google Scholar] [CrossRef] [PubMed]
- Walters, D.M.; Raikow, D.F.; Hammerschmidt, C.R.; Mehling, M.G.; Kovach, A.; Oris, J.T. Methylmercury Bioaccumulation in Stream Food Webs Declines with Increasing Primary Production. Environ. Sci. Technol. 2015, 49, 7762–7769. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Parra, P.; Sierra-Alvarez, R.; Field, J.A. Algae as an electron donor promoting sulfate reduction for the bioremediation of acid rock drainage. J. Hazard. Mater. 2016, 317, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Russell, R.A.; Holden, P.J.; Wilde, K.L.; Neilan, B.A. Demonstration of the use of Scenedesmus and Carteria biomass to drive bacterial sulfate reduction by Desulfovibrio alcoholovorans isolated from an artificial wetland. Hydrometallurgy 2003, 71, 227–234. [Google Scholar] [CrossRef]
- Parks, J.M.; Johs, A.; Podar, M.; Bridou, R.; Hurt, R.A.; Smith, S.D.; Tomanicek, S.J.; Qian, Y.; Brown, S.D.; Brandt, C.C.; et al. The Genetic Basis for Bacterial Mercury Methylation. Science 2013, 339, 1332–1335. [Google Scholar] [CrossRef] [PubMed]
- King, J.K.; Kostka, J.E.; Frischer, M.E.; Saunders, F.M. Sulfate-reducing bacteria methylate mercury at variable rates in pure culture and in marine sediments. Appl. Environ. Microbiol. 2000, 66, 2430–2437. [Google Scholar] [CrossRef] [PubMed]
- Watras, C.J.; Back, R.C.; Halvorsen, S.; Hudson, R.J.M.; Morrison, K.A.; Wente, S.P. Bioaccumulation of mercury in pelagic freshwater food webs. Sci. Total Environ. 1998, 219, 183–208. [Google Scholar] [CrossRef]
- Mason, R.; Reinfelder, J.; Morel, F. Bioaccumulation of mercury and methylmercury. Water Air Soil Pollut. 1995, 80, 915–921. [Google Scholar] [CrossRef]
- Watanabe, C.; Satoh, H. Evolution of our understanding of methylmercury as a health threat. Environ. Health Perspect. 1996, 104, 367–379. [Google Scholar] [PubMed]
- Yokoo, E.M.; Valente, J.G.; Grattan, L.; Schmidt, S.L.; Platt, I.; Silbergeld, E.K. Low level methylmercury exposure affects neuropsychological function in adults. Environ. Health 2003, 2, 8. [Google Scholar] [CrossRef] [PubMed]
- Gilmour, C.C.; Podar, M.; Bullock, A.L.; Graham, A.M.; Brown, S.D.; Somenahally, A.C.; Johs, A.; Hurt, R.A.; Bailey, K.L.; Elias, D.A. Mercury Methylation by Novel Microorganisms from New Environments. Environ. Sci. Technol. 2013, 47, 11810–11820. [Google Scholar] [CrossRef] [PubMed]
- Compeau, G.C.; Bartha, R. Sulfate-reducing bacteria: Principal methylators of mercury in anoxic estuarine sediment. Appl. Environ. Microbiol. 1985, 50, 498–502. [Google Scholar] [PubMed]
- Hellal, J.; Guédron, S.; Huguet, L.; Schafer, J.; Laperche, V.; Joulian, C.; Lanceleur, L.; Burnol, A.; Ghestem, J.P.; Garrido, F.; et al. Mercury mobilization and speciation linked to bacterial iron oxide and sulfate reduction: A column study to mimic reactive transfer in an anoxic aquifer. J. Contam. Hydrol. 2015, 180, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Fleming, E.J.; Mack, E.E.; Green, P.G.; Nelson, D.C. Mercury Methylation from Unexpected Sources: Molybdate-Inhibited Freshwater Sediments and an Iron-Reducing Bacterium. Appl. Environ. Microbiol. 2006, 72, 457–464. [Google Scholar] [CrossRef] [PubMed]
- Hamelin, S.P.; Amyot, M.; Barkay, T.; Wang, Y.; Planas, D. Methanogens: Principal Methylators of Mercury in Lake Periphyton. Environ. Sci. Technol. 2011, 45, 7693–7700. [Google Scholar] [CrossRef] [PubMed]
- Achá, D.; Pabón, C.A.; Hintelmann, H. Mercury methylation and hydrogen sulfide production among unexpected strains isolated from periphyton of two macrophytes of the Amazon. FEMS Microbiol. Ecol. 2012, 80, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Eckley, C.; Watras, C.J.; Hintelmann, H.; Morrison, K.; Kent, A.D.; Regnell, O. Mercury methylation in the hypolimnetic waters of lakes with and without connection to wetlands in northern Wisconsin. Can. J. Fish. Aquat. Sci. 2005, 62, 400–411. [Google Scholar] [CrossRef]
- Watras, C.J.; Bloom, N.S.; Claas, S.A.; Morrison, K.A.; Gilmour, C.C.; Craig, S.R. Methylmercury production in the anoxic hypolimnion of a Dimictic Seepage Lake. Water Air Soil Pollut. 1995, 80, 735–745. [Google Scholar] [CrossRef]
- Compeau, G.C.; Bartha, R. Effect of Salinity on Mercury-Methylating Activity of Sulfate-Reducing Bacteria in Estuarine Sediments. Appl. Environ. Microbiol. 1987, 53, 261–265. [Google Scholar] [PubMed]
- Gilmour, C.C.; Henry, E.A.; Mitchell, R. Sulfate Stimulation of Mercury Methylation In Freshwater Sediments. Environ. Sci. Technol. 1992, 26, 2281–2287. [Google Scholar] [CrossRef]
- Guimarães, J.R.D.; Roulet, M.; Lucotte, M.; Mergler, D. Mercury methylation along a lake-forest transect in the Tapajós river floodplain, Brazilian Amazon: Seasonal and vertical variations. Sci. Total Environ. 2000, 261, 91–98. [Google Scholar] [CrossRef]
- Desrosiers, M.; Planas, D.; Mucci, A. Mercury methylation in the epilithon of boreal shield aquatic ecosystems. Environ. Sci. Technol. 2006, 40, 1540–1546. [Google Scholar] [CrossRef] [PubMed]
- Guédron, S.; Grimaldi, M.; Grimaldi, C.; Cossa, D.; Tisserand, D.; Charlet, L. Amazonian former gold mined soils as a source of methylmercury: Evidence from a small scale watershed in French Guiana. Water Res. 2011, 45, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Mauro, J.B.; Guimaraes, J.R.; Hintelmann, H.; Watras, C.J.; Haack, E.A.; Coelho-Souza, S.A. Mercury methylation in macrophytes, periphyton, and water—Comparative studies with stable and radio-mercury additions. Anal. Bioanal. Chem. 2002, 374, 983–989. [Google Scholar] [PubMed]
- Acha, D.; Iniguez, V.; Roulet, M.; Guimaraes, J.R.; Luna, R.; Alanoca, L.; Sanchez, S. Sulfate-reducing bacteria in floating macrophyte rhizospheres from an Amazonian floodplain lake in Bolivia and their association with Hg methylation. Appl. Environ. Microbiol. 2005, 71, 7531–7535. [Google Scholar] [CrossRef] [PubMed]
- Molina, C.I.; Gibon, F.-M.; Duprey, J.-L.; Dominguez, E.; Guimaraes, J.R.D.; Roulet, M. Transfer of mercury and methylmercury along macroinvertebrate food chains in a floodplain lake of the Beni River, Bolivian Amazonia. Sci. Total Environ. 2010, 408, 3382–3391. [Google Scholar] [CrossRef] [PubMed]
- Hsu-Kim, H.; Eckley, C.S.; Achá, D.; Feng, X.; Gilmour, C.C.; Jonsson, S.; Mitchell, C.P.J. Challenges and opportunities for managing aquatic mercury pollution in altered landscapes. Ambio 2018, 47, 141–169. [Google Scholar] [CrossRef] [PubMed]
- Abbott, M.B.; Wolfe, B.B.; Wolfe, A.P.; Seltzer, G.O.; Aravena, R.; Mark, B.G.; Polissar, P.J.; Rodbell, D.T.; Rowe, H.D.; Vuille, M. Holocene paleohydrology and glacial history of the central Andes using multiproxy lake sediment studies. Palaeogeogr. Palaeoclim. Palaeoecol. 2003, 194, 123–138. [Google Scholar] [CrossRef]
- Bradley, R.S.; Vuille, M.; Diaz, H.F.; Vergara, W. Threats to Water Supplies in the Tropical Andes. Science 2006, 312, 1755–1756. [Google Scholar] [CrossRef] [PubMed]
- Vuille, M.; Francou, B.; Wagnon, P.; Juen, I.; Kaser, G.; Mark, B.G.; Bradley, R.S. Climate change and tropical Andean glaciers: Past, present and future. Earth Sci. Rev. 2008, 89, 79–96. [Google Scholar] [CrossRef]
- Perry, L.B.; Seimon, A.; Andrade-Flores, M.F.; Endries, J.L.; Yuter, S.E.; Velarde, F.; Arias, S.; Bonshoms, M.; Burton, E.J.; Winkelmann, I.R.; et al. Characteristics of Precipitating Storms in Glacierized Tropical Andean Cordilleras of Peru and Bolivia. Ann. Am. Assoc. Geogr. 2017, 107, 309–322. [Google Scholar] [CrossRef]
- Delclaux, F.; Coudrain, A.; Condom, T. Evaporation estimation on Lake Titicaca: A synthesis review and modelling. Hydrol. Process. 2007, 21, 1664–1677. [Google Scholar] [CrossRef]
- Weide, D.M.; Fritz, S.C.; Hastorf, C.A.; Bruno, M.C.; Baker, P.A.; Guédron, S.; Salenbien, W. A ~6000 yr diatom record of mid- to late Holocene fluctuations in the level of Lago Wiñaymarca, Lake Titicaca (Peru/Bolivia). Quat. Res. 2017, 88, 179–192. [Google Scholar] [CrossRef]
- Fritz, S.C.; Baker, P.A.; Tapia, P.; Spanbauer, T.; Westover, K. Evolution of the Lake Titicaca basin and its diatom flora over the last ~370,000 years. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 317–318, 93–103. [Google Scholar] [CrossRef]
- Cross, S.L.; Baker, P.A.; Seltzer, G.O.; Fritz, S.C.; Dunbar, R.B. Late Quaternary climate and hydrology of tropical South America inferred from an isotopic and chemical model of Lake Titicaca, Bolivia and Peru. Quat. Res. 2001, 56, 1–9. [Google Scholar] [CrossRef]
- Fritz, S.C.; Metcalfe, S.E.; Dean, W. Holocene climate patterns in the Americas inferred from paleolimnological records. In Interhemispheric Climate Linkages; Elsevier: San Diego, CA, USA, 2001; pp. 241–263. [Google Scholar]
- Baker, P.A.; Fritz, S.C.; Garland, J.; Ekdahl, E. Holocene hydrologic variation at Lake Titicaca, Bolivia/Peru, and its relationship to North Atlantic climate variation. J. Quat. Sci. Publ. Quat. Res. Assoc. 2005, 20, 655–662. [Google Scholar] [CrossRef]
- Francou, B.; Vuille, M.; Wagnon, P.; Mendoza, J.; Sicart, J.-E. Tropical climate change recorded by a glacier in the central Andes during the last decades of the twentieth century: Chacaltaya, Bolivia, 16°S. J. Geophys. Res. Atmos. 2003, 108, 4154. [Google Scholar] [CrossRef]
- Heidinger, H.; Carvalho, L.; Jones, C.; Posadas, A.; Quiroz, R. A new assessment in total and extreme rainfall trends over central and southern Peruvian Andes during 1965–2010. Int. J. Climatol. 2018, 38, e998–e1015. [Google Scholar] [CrossRef]
- Vera, C.; Silvestri, G.; Liebmann, B.; González, P. Climate change scenarios for seasonal precipitation in South America from IPCC-AR4 models. Geophy. Res. Lett. 2006, 33, L13707. [Google Scholar] [CrossRef]
- Fischer, E.M.; Knutti, R. Anthropogenic contribution to global occurrence of heavy-precipitation and high-temperature extremes. Nature Clim. Chang. 2015, 5, 560–564. [Google Scholar] [CrossRef]
- Paerl, H.W.; Paul, V.J. Climate change: Links to global expansion of harmful cyanobacteria. Water Res. 2012, 46, 1349–1363. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, P.G.; Wilby, R.L.; Battarbee, R.W.; Kernan, M.; Wade, A.J. A review of the potential impacts of climate change on surface water quality. Hydrol. Sci. J. 2009, 54, 101–123. [Google Scholar] [CrossRef]
- O’Donnell, D.R.; Wilburn, P.; Silow, E.A.; Yampolsky, L.Y.; Litchman, E. Nitrogen and phosphorus colimitation of phytoplankton in Lake Baikal: Insights from a spatial survey and nutrient enrichment experiments. Limnol. Oceanogr. 2017, 62, 1383–1392. [Google Scholar] [CrossRef]
- Heisler, J.; Glibert, P.; Burkholder, J.; Anderson, D.; Cochlan, W.; Dennison, W.; Gobler, C.; Dortch, Q.; Heil, C.; Humphries, E.; et al. Eutrophication and Harmful Algal Blooms: A Scientific Consensus. Harmful Algae 2008, 8, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Dale, B.; Edwards, M.; Reid, P.C. Climate Change and Harmful Algal Blooms. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Springer: Heidelberg, Germany, 2006; pp. 367–378. [Google Scholar]
- Freitas, R.; Vieira, H.H.; de Moraes, G.P.; de Melo, M.L.; Vieira, A.A.H.; Sarmento, H. Productivity and rainfall drive bacterial metabolism in tropical cascading reservoirs. Hydrobiologia 2018, 809, 233–246. [Google Scholar] [CrossRef]
- Booth, S.; Zeller, D. Mercury, Food Webs, and Marine Mammals: Implications of Diet and Climate Change for Human Health. Environ. Health Perspect. 2005, 113, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Ronchail, J.; Espinosa, J.C.; Labat, D.; Callede, J.; Lavado, W. Evolution of the Titicaca Lake level during the 20th century. In Línea Base de Conocimientos Sobre Los Recursos Hidrológicos e Hidrobiológicos en el Sistema TDPS con Enfoque en la Cuenca del Lago Titicaca; Aguirre, M., Pouilly, M., Lazzaro, X., Point, D., Eds.; UICN-IRD: La Paz, Bolivia, 2014; pp. 1–13. [Google Scholar]
- El Lago Titicaca: Síntesis del Conocimiento Limnológico Actual; Dejoux, C., Iltis, A., Eds.; ORSTOM—HISBOL: La Paz, Bolivia, 1991; p. 584. [Google Scholar]
- Archundia, D.; Duwig, C.; Spadini, L.; Uzu, G.; Guédron, S.; Morel, M.C.; Cortez, R.; Ramos Ramos, O.; Chincheros, J.; Martins, J.M.F. How Uncontrolled Urban Expansion Increases the Contamination of the Titicaca Lake Basin (El Alto, La Paz, Bolivia). Water Air Soil Pollut. 2017, 228, 44. [Google Scholar] [CrossRef]
- Guédron, S.; Point, D.; Acha, D.; Bouchet, S.; Baya, P.A.; Tessier, E.; Monperrus, M.; Molina, C.I.; Groleau, A.; Chauvaud, L.; et al. Mercury contamination level and speciation inventory in Lakes Titicaca & Uru-Uru (Bolivia): Current status and future trends. Environ. Pollut. 2017, 231, 262–270. [Google Scholar]
- Truesdale, G.A.; Downing, A.L.; Lowden, G.F. The solubility of oxygen in pure water and sea-water. J. Appl. Chem. 1955, 5, 53–62. [Google Scholar] [CrossRef]
- Villafañe, V.E.; Andrade, M.; Lairana, V.; Zaratti, F.; Helbling, E.W. Inhibition of phytoplankton photosynthesis by solar ultraviolet radiation: Studies in Lake Titicaca, Bolivia. Freshw. Biol. 1999, 42, 215–224. [Google Scholar] [CrossRef]
- Helbling, E.W.; Villafañe, V.; Buma, A.; Andrade, M.; Zaratti, F. DNA damage and photosynthetic inhibition induced by solar ultraviolet radiation in tropical phytoplankton (Lake Titicaca, Bolivia). Eur. J. Phycol. 2001, 36, 157–166. [Google Scholar] [CrossRef]
- Molina, J.; Satge, F.; Pillco, R. Los recursos hídricos del TDPS. In Línea Base de Conocimientos Sobre Los Recursos Hidrológicos en el Sistema TDPS. Con Enfoque en la Cuenca del Lago Titicaca; Puilly, M., Lazzaro, X., Point, D., Aguirre, M., Eds.; IRD—UIC: Quito, Ecuador, 2014; pp. 15–39. [Google Scholar]
- Reese, B.; Finneran, D.; Mills, H.; Zhu, M.-X.; Morse, J. Examination and Refinement of the Determination of Aqueous Hydrogen Sulfide by the Methylene Blue Method. Aquat. Geochem. 2011, 17, 567–582. [Google Scholar] [CrossRef]
- Small, J.; Hintelmann, H. Methylene blue derivatization then LC–MS analysis for measurement of trace levels of sulfide in aquatic samples. Anal. Bioanal. Chem. 2007, 387, 2881–2886. [Google Scholar] [CrossRef] [PubMed]
- Small, J.M.; Hintelmann, H. Sulfide and mercury species profiles in two Ontario boreal shield lakes. Chemosphere 2014, 111, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Monperrus, M.; Rodriguez Gonzalez, P.; Amouroux, D.; Garcia Alonso, J.I.; Donard, O.F. Evaluating the potential and limitations of double-spiking species-specific isotope dilution analysis for the accurate quantification of mercury species in different environmental matrices. Anal. Bioanal. Chem. 2008, 390, 655–666. [Google Scholar] [CrossRef] [PubMed]
- Lund, J.W.G.; Kipling, C.; Le Cren, E.D. The inverted microscope method of estimating algal numbers and the statistical basis of estimations by counting. Hydrobiologia 1958, 11, 143–170. [Google Scholar] [CrossRef]
- Bellinger, E.G.; Sigee, D.C. Freshwater Algae: Identification and Use as Bioindicators; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Wehr, J.D.; Sheath, R.G.; Kociolek, J.P. Freshwater Algae of North America: Ecology and Classification; Elsevier: Boston, MA, USA, 2015. [Google Scholar]
- Beamud, S.G.; León, J.G.; Kruk, C.; Pedrozo, F.; Diaz, M. Using trait-based approaches to study phytoplankton seasonal succession in a subtropical reservoir in arid central western Argentina. Environ. Monit. Assess. 2015, 187, 271. [Google Scholar] [CrossRef] [PubMed]
- Al-Homaidan, A.A.; Arif, I.A. Ecology and bloom-forming algae of a semi-permanent rain-fed pool at Al-Kharj, Saudi Arabia. J. Arid Environ. 1998, 38, 15–25. [Google Scholar] [CrossRef]
- Rushforth, S.R.; St. Clair, L.L.; Grimes, J.A.; Whiting, M.C. Phytoplankton of Utah Lake. Great Basin Nat. Mem. 1981, 5, 85–100. [Google Scholar]
- Janta, K.; Pekkoh, J.; Tongsiri, S.; Pumas, C.; Peerapornpisal, Y. Selection of some native microalgal strains for possibility of bio-oil production in Thailand. Chiang Mai J. Sci. 2013, 40, 593–602. [Google Scholar]
- Bouvy, M.; Molica, R.; De Oliveira, S.; Marinho, M.; Beker, B. Dynamics of a toxic cyanobacterial bloom (Cylindrospermopsis raciborskii) in a shallow reservoir in the semi-arid region of northeast Brazil. Aquat. Microb. Ecol. 1999, 20, 285–297. [Google Scholar] [CrossRef]
- Menezes, M.; Bicudo, C.E.d.M. Flagellate green algae from four water bodies in the state of Rio de Janeiro, Southeast Brazil. Hoehnea 2008, 35, 435–468. [Google Scholar] [CrossRef]
- Collins, P.A.; Williner, V. Feeding of Acetes paraguayensis (Nobili) (Decapoda: Sergestidae) from the Parana River, Argentina. Hydrobiologia 2003, 493, 1–6. [Google Scholar] [CrossRef]
- Lin, L.; He, J.; Zhang, F.; Cao, S.; Zhang, C. Algal bloom in a melt pond on Canada Basin pack ice. Polar Rec. 2016, 52, 114–117. [Google Scholar] [CrossRef]
- Perez, W. Dos Toneladas de Ranas, Reces y Aves Mueren en el Titicaca. Available online: http://www.la-razon.com/sociedad/toneladas-ranas-peces-mueren-Titicaca_0_2259374132.html (accessed on 24 April 2015).
- Peck, L.S.; Chapelle, G. Reduced oxygen at high altitude limits maximum size. Proc. R. Soc. B Biol. Sci. 2003, 270, S166–S167. [Google Scholar] [CrossRef] [PubMed]
- Michalak, A.M.; Anderson, E.J.; Beletsky, D.; Boland, S.; Bosch, N.S.; Bridgeman, T.B.; Chaffin, J.D.; Cho, K.; Confesor, R.; Daloğlu, I.; et al. Record-setting algal bloom in Lake Erie caused by agricultural and meteorological trends consistent with expected future conditions. Proc. Natl. Acad. Sci. USA 2013, 110, 6448–6452. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.; Turner, M.; Hesslein, R. Acidification and alkalinization of lakes by experimental addition of nitrogen compounds. Biogeochemistry 1985, 1, 117–133. [Google Scholar] [CrossRef]
- Verspagen, J.M.H.; Van de Waal, D.B.; Finke, J.F.; Visser, P.M.; Van Donk, E.; Huisman, J. Rising CO2 Levels Will Intensify Phytoplankton Blooms in Eutrophic and Hypertrophic Lakes. PLoS ONE 2014, 9, e104325. [Google Scholar] [CrossRef] [PubMed]
- Ibelings, B.W.; Maberly, S.C. Photoinhibition and the availability of inorganic carbon restrict photosynthesis by surface blooms of cyanobacteria. Limnol. Oceanogr. 1998, 43, 408–419. [Google Scholar] [CrossRef]
- Balmer, M.B.; Downing, J.A. Carbon dioxide concentrations in eutrophic lakes: Undersaturation implies atmospheric uptake. Inland Waters 2011, 1, 125–132. [Google Scholar] [CrossRef]
- Sobczyłski, T.; Joniak, T. The Variability and Stability of Water Chemistry in a Deep Temperate Lake: Results of Long-Term Study of Eutrophication. Pol. J. Environ. Stud. 2013, 22, 227–237. [Google Scholar]
- Søndergaard, M.; Jensen, J.P.; Jeppesen, E. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia 2003, 506, 135–145. [Google Scholar] [CrossRef]
- Jensen, H.S.; Andersen, F.O. Importance of temperature, nitrate, and pH for phosphate release from aerobic sediments of four shallow, eutrophic lakes. Limnol. Oceanogr. 1992, 37, 577–589. [Google Scholar] [CrossRef]
- Søndergaard, M.; Kristensen, P.; Jeppesen, E. Phosphorus release from resuspended sediment in the shallow and wind-exposed Lake Arresø, Denmark. Hydrobiologia 1992, 228, 91–99. [Google Scholar] [CrossRef]
- Effler, S.W.; Hassett, J.P.; Auer, M.T.; Johnson, N. Depletion of epilimnetic oxygen and accumulation of hydrogen sulfide in the hypolimnion of Onondaga Lake, NY, USA. Water Air Soil Pollut. 1988, 39, 59–74. [Google Scholar] [CrossRef]
- Reese, B.K.; Anderson, M.A.; Amrhein, C. Hydrogen sulfide production and volatilization in a polymictic eutrophic saline lake, Salton Sea, California. Sci. Total Environ. 2008, 406, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Luther, G.W.; Findlay, A.J.; MacDonald, D.J.; Owings, S.M.; Hanson, T.E.; Beinart, R.A.; Girguis, P.R. Thermodynamics and Kinetics of Sulfide Oxidation by Oxygen: A Look at Inorganically Controlled Reactions and Biologically Mediated Processes in the Environment. Front. Microbiol. 2011, 2, 62. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, M.; Jorgensen, B.B. Microsensor Measurements of Sulfate Reduction and Sulfide Oxidation in Compact Microbial Communities of Aerobic Biofilms. Appl. Environ. Microbiol. 1992, 58, 1164–1174. [Google Scholar] [PubMed]
- Myrbo, A.; Swain, E.B.; Johnson, N.W.; Engstrom, D.R.; Pastor, J.; Dewey, B.; Monson, P.; Brenner, J.; Dykhuizen Shore, M.; Peters, E.B. Increase in Nutrients, Mercury, and Methylmercury as a Consequence of Elevated Sulfate Reduction to Sulfide in Experimental Wetland Mesocosms. J. Geophys. Res. Biogeosci. 2017, 122, 2769–2785. [Google Scholar] [CrossRef]
- Morse, J.W.; Millero, F.J.; Cornwell, J.C.; Rickard, D. The chemistry of the hydrogen sulfide and iron sulfide systems in natural waters. Earth-Sci. Rev. 1987, 24, 1–42. [Google Scholar] [CrossRef]
- Canfield, D.E.; Stewart, F.J.; Thamdrup, B.; De Brabandere, L.; Dalsgaard, T.; Delong, E.F.; Revsbech, N.P.; Ulloa, O. A cryptic sulfur cycle in oxygen-minimum-zone waters off the Chilean coast. Science 2010, 330, 1375–1378. [Google Scholar] [CrossRef] [PubMed]
- WHO. Guidelines for Drinking-Water Quality: Fourth Edition Incorporating the First Addendum; Licence: CC BY-NC-SA 3.0 IGO; World Health Organization: Geneva, Switzerland, 2017; p. 631. [Google Scholar]
- US-EPA. In Quality Criteria for Water; EPA 550/5–86/001; Agency, E.P., Ed.; Environmental Protection Agency: Cincinnati, OH, USA, 1986. [Google Scholar]
- Bagarinao, T. Sulfide as an environmental factor and toxicant: Tolerance and adaptations in aquatic organisms. Aquat. Toxicol. 1992, 24, 21–62. [Google Scholar] [CrossRef]
- Lamers, L.P.; Govers, L.L.; Janssen, I.C.; Geurts, J.J.; Van der Welle, M.E.; Van Katwijk, M.M.; Van der Heide, T.; Roelofs, J.G.; Smolders, A.J. Sulfide as a soil phytotoxin—A review. Front. Plant Sci. 2013, 4, 268. [Google Scholar] [CrossRef] [PubMed]
- Knezovich, J.P.; Steichen, D.J.; Jelinski, J.A.; Anderson, S.L. Sulfide Tolerance of Four Marine Species Used to Evaluate Sediment and Pore-Water Toxicity. Bull. Environ. Contam. Toxic. 1996, 57, 450–457. [Google Scholar] [CrossRef]
- Kinsman-Costello, L.E.; O’Brien, J.M.; Hamilton, S.K. Natural stressors in uncontaminated sediments of shallow freshwaters: The prevalence of sulfide, ammonia, and reduced iron. Environ. Toxic. Chem. 2015, 34, 467–479. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Chapman, P.M. Biological implications of sulfide in sediment—A review focusing on sediment toxicity. Environ. Toxic. Chem. 1999, 18, 2526–2532. [Google Scholar] [CrossRef]
- Myrbo, A.; Swain, E.R.; Engstrom, D.; Coleman Wasik, J.; Brenner, J.; Dykhuizen Shore, M.B.; Peters, E.; Blaha, G. Sulfide Generated by Sulfate Reduction is a Primary Controller of the Occurrence of Wild Rice (Zizania palustris) in Shallow Aquatic Ecosystems. Sulfide Occur. Wild Rice 2017, 122, 2736–2753. [Google Scholar]
- Sundberg-Jones, S.E.; Hassan, S.M. Macrophyte Sorption and Bioconcentration of Elements in a Pilot Constructed Wetland for Flue Gas Desulfurization Wastewater Treatment. Water Air Soil Pollut. 2007, 183, 187–200. [Google Scholar] [CrossRef]
- Lanza, W.G.; Achá, D.; Point, D.; Masbou, J.; Alanoca, L.; Amouroux, D.; Lazzaro, X. Association of a Specific Algal Group with Methylmercury Accumulation in Periphyton of a Tropical High-Altitude Andean Lake. Arch. Environ. Contam. Toxic. 2017, 72, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bouchet, S.; Goñi-Urriza, M.; Monperrus, M.; Guyoneaud, R.; Fernandez, P.; Heredia, C.; Tessier, E.; Gassie, C.; Point, D.; Guédron, S.; et al. Linking Microbial Activities and Low-Molecular-Weight Thiols to Hg Methylation in Biofilms and Periphyton from High-Altitude Tropical Lakes in the Bolivian Altiplano. Environ. Sci. Technol. 2018, 52, 9758–9767. [Google Scholar] [CrossRef] [PubMed]
- Verburg, P.; Hickey, C.W.; Phillips, N. Mercury biomagnification in three geothermally-influenced lakes differing in chemistry and algal biomass. Sci. Total Environ. 2014, 493, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Pickhardt, P.C.; Folt, C.L.; Chen, C.Y.; Klaue, B.; Blum, J.D. Impacts of zooplankton composition and algal enrichment on the accumulation of mercury in an experimental freshwater food web. Sci. Total Environ. 2005, 339, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Soerensen, A.L.; Schartup, A.T.; Gustafsson, E.; Gustafsson, B.G.; Undeman, E.M.; Björn, E. Eutrophication increases phytoplankton methylmercury concentrations in a coastal sea—A Baltic Sea case study. Environ. Sci. Technol. 2016, 50, 11787–11796. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yin, Y.; Li, Y.; Cai, Y.; Liu, J. Critical role of natural organic matter in photodegradation of methylmercury in water: Molecular weight and interactive effects with other environmental factors. Sci. Total Environ. 2017, 578, 535–541. [Google Scholar] [CrossRef] [PubMed]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Achá, D.; Guédron, S.; Amouroux, D.; Point, D.; Lazzaro, X.; Fernandez, P.E.; Sarret, G. Algal Bloom Exacerbates Hydrogen Sulfide and Methylmercury Contamination in the Emblematic High-Altitude Lake Titicaca. Geosciences 2018, 8, 438. https://doi.org/10.3390/geosciences8120438
Achá D, Guédron S, Amouroux D, Point D, Lazzaro X, Fernandez PE, Sarret G. Algal Bloom Exacerbates Hydrogen Sulfide and Methylmercury Contamination in the Emblematic High-Altitude Lake Titicaca. Geosciences. 2018; 8(12):438. https://doi.org/10.3390/geosciences8120438
Chicago/Turabian StyleAchá, Darío, Stephane Guédron, David Amouroux, David Point, Xavier Lazzaro, Pablo Edgar Fernandez, and Géraldine Sarret. 2018. "Algal Bloom Exacerbates Hydrogen Sulfide and Methylmercury Contamination in the Emblematic High-Altitude Lake Titicaca" Geosciences 8, no. 12: 438. https://doi.org/10.3390/geosciences8120438
APA StyleAchá, D., Guédron, S., Amouroux, D., Point, D., Lazzaro, X., Fernandez, P. E., & Sarret, G. (2018). Algal Bloom Exacerbates Hydrogen Sulfide and Methylmercury Contamination in the Emblematic High-Altitude Lake Titicaca. Geosciences, 8(12), 438. https://doi.org/10.3390/geosciences8120438