Abstract
Mercury (Hg) remains hazardous in aquatic environments, because of its toxicity and high biomagnification in food webs. In phytoplankton and macrophytes, Hg compounds at high concentration have been reported to affect the growth, photosynthesis, and nutrient metabolism, as well as to induce oxidative stress and damage. Here, we reviewed the recent knowledge gained on cellular toxicity of inorganic and methyl Hg (IHg; MeHg) in aquatic primary producers at more relevant environmental concentrations, with a particular focus on omics data. In addition, we compared a case study conducted with transcriptomic on the green microalga Chlamydomonas reinhardtii and the macrophyte Elodea nuttallii. At lower concentrations, IHg and MeHg influenced similar gene categories, including energy metabolism, cell structure, and nutrition. In addition, genes involved in the cell motility in the microalgae, and in hormone metabolism in the macrophyte were regulated. At equivalent intracellular concentration, MeHg regulated more genes than IHg supporting a higher molecular impact of the former. At the organism level in C. reinhardtii, MeHg increased reactive oxygen species, while both IHg and MeHg increased photosynthesis efficiency, whereas in E. nuttallii MeHg induced anti-oxidant responses and IHg reduced chlorophyll content. Data showed differences, according to species and characteristics of life cycle, in responses at the gene and cellular levels, but evidenced a higher molecular impact of MeHg than IHg and different cellular toxicity pathways in aquatic primary producers.
1. Introduction
Mercury (Hg) contamination of freshwaters arises from the deposition of atmospheric Hg (dry settling or rainfall), polluted soils runoffs, industrial effluents, gold-mining and reemission from historical contaminated systems ([1,2]). It subsequently settles in the sediment of lakes, rivers or bays where it is transformed into methylmercury (MeHg), absorbed by primary producers, transferred to primary and secondary consumers, and accumulates especially in long-lived predatory species, e.g., shark and swordfish. In surface waters, Hg exists as various forms: elemental Hg0 (highly volatile) and divalent Hg (HgII), present in both inorganic Hg (HgII, IHg) and MeHg (CH3Hg+) forms. Hg enters aquatic environment mainly as IHg, which consequently is the predominant form in freshwaters. In the water column, the dissolved gaseous Hg0 accounts for less than 30% of total Hg (THg = IHg + MeHg + Hg0), while MeHg is the scarcest form, generally representing less than 1% of THg, but up to 30% in some conditions (e.g., [3,4]). Typical environmental concentrations measured in rivers and lakes range between 0.003 and 30 nM THg ([5]). Both IHg and MeHg are mainly bound to dissolved organic matter (DOM) ([1,6,7]) in particular to thiol (RSH) groups. For the fraction of Hg not bound with DOM, water chemistry (e.g., pH and chloride concentration) controls its distribution among different dissolved chemical species and complexes ([8,9]). In freshwaters, predominant species in the dissolved portion of Hg are hydroxo- (e.g., Hg(OH)+, CH3HgOH) and chloro- (e.g., HgCl2, HgCl+, CH3HgCl) complexes ([8]). It has been shown that IHg and MeHg show distinct affinities to DOM and ions in water ([1,6,7]). Nevertheless, currently the measurement and the modelling of speciation of Hg in natural waters and linking Hg speciation with uptake in biota both remain challenging. Studies have shown that IHg and MeHg both bioaccumulate in aquatic organisms ([4,10,11]). As a soft-acid, Hg exhibits a strong affinity to –SH functional groups of essential biomolecules (e.g., enzymes), displaces essential ions from such groups, or modify their conformation making them non-functional and impacting key metabolism such as energy metabolism or growth, as well as increases the generation of reactive oxygen species (ROS) resulting in oxidative stress when cell defenses are overwhelmed ([12]). For this reason, Hg and its compounds are listed as priority hazardous substances ([13]).
Ecotoxicology studies effects of pollutants on ecosystems at different levels of biological organization. One of the core missions of ecotoxicology is to understand the mechanisms by which contaminants perturb normal biological performance linking responses at molecular and cellular levels to the whole organism, population and community level effects. Indeed, the effects at high hierarchical levels are always preceded by early changes at lower level of organization ([14]). In recent years, the development of high-throughput omics has revolutionized ecotoxicology. Molecular biology approaches give impetus for new understanding of the underlying processes and mechanisms determining Hg species toxicity, by allowing to evidence early molecular and cellular toxicity at environmental concentrations. The present review paper focusses on the Hg effects on primary producers. Primary producers sustain ecosystems by biomass production and play major ecological roles in freshwater ecosystems as a source of food for high trophic chain levels, and in nutrient cycling. Primary producers comprise a vast diversity of photosynthetic organisms, of which phytoplankton and macrophytes, represent the major groups in aquatic environment ([15,16]). These two groups include cyanobacteria, planktonic algae (Chlorophytes and diatoms), large algae, bryophytes and vascular plants including both aquatic spermatophytes (flowering plants), as well as pteridophytes (ferns). In a wide range of littoral ecosystems (i.e., rivers, marshes, ponds and lakes), macrophytes represent the predominant group of organisms, in terms of biomass and primary production, while phytoplankton, accounting for less than 1% of photosynthetic biomass, sustains the largest ecosystem on the Earth contributing to about half of the primary production on our planet ([15,16]). Both phytoplankton and macrophytes are highly complementary organisms for ecotoxicology studies: the former is typically found in the water column, while rooted macrophytes occupy the benthic environment and this difference in the habitat creates substantial differences with respect to the amount and exposure length of toxicants, including Hg ([6,17]). In recent years, several publications reviewed the uptake and effect of Hg in phototroph ([18]), phytoplankton ([19]) and macrophytes ([20]). Here, our aim was thus to review comprehensively the recent studies on Hg bioaccumulation (Section 2) and Hg effects on different levels of biological organization of primary producers (Section 3), with a particular focus on recent omics data. Subsequently, we used a case study to compare omics data in microalgae and a macrophyte (Section 4). In particular, the role of omics data in identifying cellular toxicity pathways at different levels of biological organization and cross-species was discussed.
2. Bioaccumulation of Hg in Aquatic Primary Producers
Bioaccumulation of IHg and MeHg certainly determines Hg impact in primary producers. In freshwaters, concentrations of Hg can be very low (fM). However, macrophytes and algae have shown high bioaccumulation potential ([4,21,22,23]). In field-collected algae and macrophytes, concentrations usually range between 0.01–0.2 µg·g−1 THg dry weight (dw) ([5]). In a contaminated site (Olt river, Romania), concentration of THg in plankton reached 0.7 µg·g−1dw THg ([4]). In the same site, Potamogeton pectinatus and Elodea nuttallii showed concentrations up to 2 µg·g−1dw THg ([22]). This difference in uptake might be due to different life forms (floating vs. benthic) or/and different uptake capacities. Recently, authors measured close to 6 µg·g−1dw THg in roots of Typha domingensis collected in Valdeazogues River (Spain) ([24]). Overall, reported data from the field of ratio of uptaken Hg to measured Hg in the exposure water (L·kg−1) present log10 values between 4 and 6 for both macrophytes and phytoplankton ([4,19,22]), suggesting important bioaccumulation potential and evidencing the importance of Hg uptake in primary producers for its further trophic transfer. Notably, bioaccumulation in primary producers was repeatedly calculated as the biggest bioconcentration step in aquatic food webs ([11,19,20,25]). Nonetheless few studies compared uptake in identical experimental conditions, limiting inter-species comparison and generalizations.
Before entering in contact with aquatic primary producers, Hg has first to diffuse from the bulk medium to the bio-interface ([19,26]). The lability and mobility of the IHg/MeHg -complexes and Hg concentrations in waters are thus key parameters (Figure 1). Nevertheless, the transport across the plasma membrane is most likely the limiting step of Hg absorption in cells ([1]). The presence of the cell wall in primary producers represents a supplemental barrier for Hg intracellular uptake ([7]). However, the transport of Hg across membranes is poorly understood in aquatic primary producers.
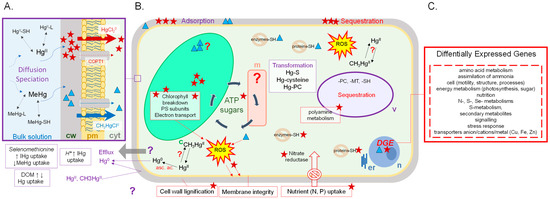
Figure 1.
Summary of the current concepts on IHg (star) and MeHg (triangle) uptake (A), effects (B) and transcriptomic response (C) in cells of aquatic primary producers (c, chloroplast; cyt, cytosol; cw, cell wall; er, endoplasmic reticulum; m, mitochondrion; n, nucleus; pm, plasma membrane; v, vacuole; other definitions in the text).
Pioneer studies with model membranes and in bacteria suggested that neutral species of Hg, such as HgCl2 and CH3HgCl, diffuse through biological membranes, whereas other species such as HgCl+, CH3Hg+, HgCl42− adsorb at the membrane surface ([27,28]). In the marine diatom Thalassiosira weissflogii, the diffusion of the neutral chloro-complexes HgCl20 and CH3HgCl0 was hypothesized to be the main uptake route of IHg and MeHg ([29]), while uptake kinetics of MeHg in Selenastrum capricornutum supported the involvement of 2 unidentified transporters, including one active ([30]). Decreased pH from 6.5 to 5.5 enhanced IHg uptake in the microalga Chlamydomonas reinhardtii exposed 30 min to 80 nM ([9]). Authors hypothesized a potential H+-IHg cotransport for internalization in C. reinhardtii ([9]). In the submerged macrophyte, E. nuttallii, exposure at low temperatures (4 °C) or exposure of dead plants to IHg and MeHg significantly reduced their accumulation ([22]). Data suggested that IHg might be accumulated in E. nuttallii shoots by diffusion through the cell wall, but IHg internalization in the cell sap was clearly linked to the metabolism ([22]). Moreover, a competition with Cu+ (more than with Cu2+) strongly reduced IHg accumulation in E. nuttallii ([22]), and supported that the Cu transporters COPT/CTRs could be the major routes for IHg assimilation, as several studies concerning eukaryotic model organisms have shown the involvement of this transporter family in Cu+ acquisition from the environment ([31,32]).
The most recent studies on transport of IHg and MeHg in aquatic primary producers and in other organisms investigate the possibility of the carrier mediated uptake of IHg and MeHg ([28]; [9,22,30]). However, while the biomagnification of MeHg vs. IHg has frequently been evidenced in the field, the precise mechanisms resulting in these differences of fate still need to be elucidated. There is clearly a need of further investigations on the effects of Hg species on essential metals’ homestasis, amino acid (e.g., selenium, methionine containing compounds) and protonated transport systems (Figure 1).
In organisms, toxicity is expected to be linked to intracellular bioaccumulation. In macrophytes and most of the phytoplanktonic organisms, cell walls and vacuoles serve as sink for Hg sequestration and protect the cellular machinery from toxicity. In E. nuttallii, the distribution of Hg at the subcellular level was similar in shoots exposed for 24 h to 0.4 nM IHg and 0.1 nM MeHg in presence of 1 mgL−1 DOM, namely close to 65% of THg was internalized, most probably in the vacuole, whereas around 40% was bound to cell walls, and 5% to membranes ([22,33]). In field observations, in the salt marsh plant Halimione portulacoides the importance of cell wall immobilization of Hg was highlighted ([34]). Hg could be retained in cell walls by means of extracellular carbohydrates, such as pectic sites or hystidyl groups forming very stable complexes. They could also be associated with thiols or cysteines of cell wall proteins like extensins ([35]). However, in C. reinhardtii, at 1 and 10 nM IHg in absence of DOM, 16 ± 1% of IHg was intracellular, while 53 ± 12% of MeHg was intracellular at similar concentrations ([36]). Similarly, in E. nuttallii exposed to 10 pM IHg and MeHg in absence of DOM, 33% of Hg was intracellular for IHg treatment, while 100% of Hg was found intracellularly for MeHg treatment ([7]). Differences in binding affinity of IHg and MeHg to DOM and cell walls could result in the higher bioaccumulation of MeHg than IHg in primary producers in natural waters, as well as consequently in a higher toxicity of MeHg in natural waters ([7]).
3. Effects of Hg Exposure on Different Levels of Biological Organization in Aquatic Primary Producers
3.1. Physiological Responses
Once uptaken, Hg compounds bind to various primordial cellular components, including DNA, proteins and enzymes and induce ROS production, potentially resulting in oxidative stress in primary producers. Exposure to 100 nM IHg in Potamogeton crispus, but also 50 nM IHg in the marine macrophyte Posidonia oceanica, resulted in an increased lipid peroxidation ([37,38,39]). Lipid peroxidation can further impair the integrity of membranes, leading potentially to cell death (Figure 1). The activity of enzymes involved in ROS level regulation like class III peroxidases (POD), superoxide dismutase (SOD), catalase, or lipoxygenase was altered by Hg stress at the same concentrations mentioned above, supporting the generation of ROS upon Hg exposure ([38,39]). For example, in the macrophyte E. nuttallii, SOD activity showed a 1.3× increase after 24 h to 0.4 and 400 nM IHg, while a 2.1× and 4.6× decrease of POD activity was measured, respectively ([40]). Excessive ROS generation in C. reinhardtii was observed by AC-dielectrophoresis with fluorescence detection sensor at short-term exposure to 100 nM IHg or 1 nM MeHg ([41]).
Beside oxidative stress, numerous studies in primary producers revealed the impact of Hg, mostly IHg, on photosynthesis, including the breakdown of chlorophyll ([19,23,39,42,43]). Photosynthesis is affected by IHg binding to several sites in the photosynthetic apparatus, notably the photosystem II (PSII) ([44,45,46]). Effect of IHg on electron transport was reported in cyanobacteria ([46,47]). Numerous studies evidenced a decrease in photosynthesis efficiency due to IHg ([47,48]). Nevertheless, a recent study in C. reinhardtii showed on the opposite an increase in photosynthesis efficiency after 2 h exposure to 10 nM and 10 pM MeHg, suggesting an hormesis effect, that is seen as an overcompensation of a moderate stress, in line with lower concentrations tested in these studies ([36,49,50]). A difference in the impact of IHg and MeHg on photosynthesis was observed in the marine diatom Thalassiosira weissflogii; where 72 h exposure to 200 nM IHg increased the lifetime of chlorophyll fluorescence by blocking the photosynthetic electron chain, whereas 72 h exposure to up to 27.8 µM MeHg exposure did not have an effect ([51]). Similarly in E. nuttallii, IHg reduced the chlorophyll content, while MeHg induced a significant anti-oxidant response ([7]). These observations suggested that IHg directly affected chloroplasts membrane integrity, whereas MeHg disturbed organelle metabolism in the cytoplasm in E. nuttallii. In an older study, however, structural damage of chloroplast was observed in Elodea densa after 25 d exposure to 0.75 nM MeHg ([52]). A more recent study reported that 24 h exposure to 0.1 nM MeHg did not result in significant effect on chlorophyll content in E. nuttallii ([33]). Overall, the photosynthetic machinery appears to be an important target of IHg toxic action, with the substitution of Mg by IHg in chlorophyll ([53]) and the chelation with chloroplast proteins, while MeHg toxicity would directly targets the cellular machinery, resulting in increased ROS production (Figure 1).
IHg was also shown to affect nutrient metabolism. In the submerged macrophyte Vallisneria spiralis and the floating fern Azolla pinnata, phosphate and nitrate uptake were inhibited by 6 d exposure to 500 nM IHg ([54]). In an earlier study with V. spiralis, 4 d exposure to 500 nM IHg decreased by 13% the activity of the nitrate reductase, which catalyzes the first step in nitrate assimilation ([55]). Nitrate reductase is a cysteine-rich protein, and the authors suggested that it might be inhibited due to binding of Hg to the –SH groups of the cysteine residues ([55]).
Finally, the growth of T. weissflogii was compared in presence of IHg or MeHg and showed that cell growth was reduced by 50% at 0.5 nM IHg and MeHg but authors suggested different cellular toxicity pathways based on two-photon excitation chlorophyll measurement: MeHg reduced growth rate, while IHg induced strong cell damage but rate of division was unchanged vs. control ([51]). In E. nuttallii, exposure 7 d to 0.35 nM IHg reduced root growth and increased lignification of cell walls, while exposure 7 d to 0.1 nM MeHg had no effect on those endpoints ([33]).
To summarize, the toxicity of Hg in aquatic primary producers is hypothesized to result from binding to sulfhydryl (-SH) groups of various proteins, with phosphate groups and active groups of ADP or ATP, and the replacement of essential ions (mainly major cations) ([42,56]). More specifically, IHg impact on the photosynthesis and induction of oxidative stress has been widely observed in phytoplankton and macrophytes. Additionally, effects of IHg on nitrogen metabolism were reported in aquatic plants. Toxicity of MeHg has been less studied than IHg in aquatic primary producers. Globally, MeHg has been shown to have similar targets than IHg (e.g., oxidative stress, photosynthesis), but it impacts other cellular toxicity pathways than IHg when analyzed in identical experimental settings. Most recent studies used pM and nM concentrations that are more relevant to environmental conditions, and provided effective concentrations, as well as some analysis or modelling of speciation to assess exposure conditions in details. In future research an effort should be made to always provide this information, as well as to conduct studies at relevant concentrations in controlled exposure as well as conduct more studies in the field in realistic exposure conditions. Assessing Hg toxicity at lower level of biological organization and new analytical methods for determining metal speciation and spatial distribution (i.e., metallomics) will result in a more sensitive and better mechanistic understanding of the IHg and MeHg molecular toxicity pathways ([57]). Short exposure time allows detection of early molecular toxicity responses that are more specific to each contaminant than a longer exposure time ([50]). In this context, omics are promising approaches, because they are sensitive and suitable for low concentration and short time of exposure ([14]).
3.2. Response at the Protein and the Gene Level
The use of omics has increased in ecotoxicological studies in recent years, thanks to the development first of DNA microarray and more recently of new sequencing technologies (e.g., RNA-Seq) ([14,58]). Transcriptomic aims at analyzing the differential gene expression in response to certain conditions vs. a reference condition, for instance under metal-stress vs. absence of metal-stress (control). Currently it can be efficiently applied to both sequenced model organisms and non-model organisms of interest. Its use is however much accurate for sequenced species, because for non-sequenced species users have to build a de novo transcriptome with an increased risk of artefacts during data processing, management, and analysis. Nevertheless, for all organisms, a main limitation of high-throughput omics is to associate a gene to a function, by comparison with known sequences available in databases for other species. Indeed, a large part of genomes has not been annotated and several gene products appeared as hypothetical proteins or unknown. However, transcriptomic informs on the level of expression of genes and allows comparisons among treatments. Nonetheless, in most cases the cellular functions of a particular gene are carried out by its protein. Proteomics informs on the level of expression of proteins and in addition on the post-transcriptional regulations that can occur. Thus, compared to RNA-seq, proteomics should provide a more direct link between stresses and resulting phenotypes. Its use is however still limited by the availability of database of known proteins profiles ([14]). Eventually, the result of all cellular regulatory activities can be assessed by metabolomics. Analyses of effects of Hg by transcriptomic and proteomic were reported in several terrestrial plants ([59,60,61]) but still rarely in aquatic primary producers (Table 1), while metabolomics was not yet applied.

Table 1.
Effects of Hg revealed by transcriptomic (T) and proteomic (P) studies in macrophytes and algae.
Pioneer work with proteomic in the macrophyte E. nuttallii revealed that 24 h-exposure to 0.4 nM IHg dysregulated about 30 proteins involved in photosynthesis, sugar metabolism and cell structure, however 0.1 nM MeHg did not result in significant response for MeHg ([33]) (Table 1). Analysis of effects of 0.004, 0.4 and 4 µM IHg 24 h was also conducted through whole transcriptome analysis in E. nuttallii ([62]) (Table 1). The macrophyte responded to IHg treatment by the up-regulation of genes coding for proteins (e.g., chaperones) known for their stress response function. A modification of genes involved in energy metabolism, notably sugar-catabolizing proteins, certainly caused by inhibited production of energy reserves through photosynthesis was observed ([62]). Down-regulation of metal transporters and genes related to homeostasis appeared to most probably control and reduce accumulation of Hg ([62]). These results also supported the involvement of oxidative stress and effects on protein structure in the mechanism of toxicity of IHg, and further highlighted that sub-lethal concentrations might result in significant changes in the metabolic production of energy and adaptation of the nutrition pathways, as well as the induction of a protective response. Moreover, data revealed the down regulation of the EnCOPT1 gene by increasing concentrations of HgII, as well as CuII, supporting the hypothesis of HgII uptake via high affinity Cu transporters ([62]). Recently, in E. nuttallii, 24 h exposure to 0.4 µM IHg induced a significant up-regulation of genes involved in lipid biosynthesis and transport of ions as well as down-regulation of genes involved in photosynthesis ([40]) (Table 1). In addition to shedding light on the early cellular toxicity pathways of Hg, the attempt of using gene expression as biomarker of exposure has shown promising results. In E. nuttallii, using the nCounter method (Nanostring), a global analysis of expression of selected genes could be realized directly on RNA ([62]). Hierarchical clustering of treatments based on gene expression signatures enabled to discriminate nM IHg from pM IHg exposure concentrations and additional clustering according to measured uptake ([62]). Another study conducted in situ, in the Olt River (Romania) contaminated by effluents of a chlor-alkali plant, on caged C. reinhardtii and E. nuttallii showed in both species a strong gene regulation after 2 h exposure, congruent with expected Hg impact at higher concentrations, although Hg concentration was in the pM range, and exposure resulted in non-significant uptake and physiological impact ([6]).
Using transcriptomics could help to gain a better mechanistic understanding of different aspects of interactions of organisms with chemicals. Above mentioned studies showed that gene and protein regulation occurred at much lower Hg concentration than other parameters, e.g., bioaccumulation or physiological endpoints, and was congruent with effects observed at higher level of organizations (see Section 3.1). Moreover, by allowing the simultaneous study of more than a thousand genes and proteins, transcriptomics and proteomics are analogous to a multibiomarker approach. Indeed, the analysis of gene and protein expression level in response to IHg and MeHg seems promising to develop rapid and sensitive molecular biomarkers of effect or exposure that are sensitive, specific, reproducible, and reliable and will allow the early detection of Hg contamination. Nonetheless, to nicely complement the standard tests, biomarkers based on omics analysis, needs to be further validated in wild populations for normalization of background levels, as is currently done in animal models ([64]). Nevertheless, few studies are available concerning primary producers yet and the various experimental settings limits still interspecific comparisons and hence generalizations to the ecosystem level. Further work is clearly needed to link molecular responses in primary producers to population and community risk assessment in the environment.
4. A Case Study: Comparison of Responses to IHg and MeHg Exposure in C. reinhardtii and E. nuttallii
We recently reported the bioaccumulation, the transcriptomic and physiological responses following 2 h exposure to IHg or MeHg in the green microalga C. reinhardtii and the macrophyte E. nuttallii in identical experimental conditions: exposure to a wide range of concentrations varying between 10 pM and 10 nM IHg or MeHg in a model freshwater exposure medium mimicking Lake Geneva water without DOM ([7,36,42,50]). IHg and MeHg impact in each species were compared ([7,36]). Briefly, in C. reinhardtii, bioavailability of MeHg was up to 27× higher than for IHg. At the cell level, all MeHg, 10 pM IHg as well as 0.1 nM IHg treatments increased photosynthesis efficiency, suggesting a hormesis effect. Only one treatment (0.1 nM MeHg) resulted in a significant increase of the cellular ROS and oxidative stress, suggesting that the microalga could efficiently cope with the range of concentrations of IHg and MeHg tested here (10 pM to 10 nM), at least for a short-exposure period. However, genes involved in cell processes, energy metabolism, transport, cell motility, nutrition, and amino acids metabolism were dysregulated by both Hg species, including for 10 pM treatment, revealing a broader impact on C. reinhardtii metabolism than expected based on physiological endpoints (Table 1). Moreover, at similar intracellular THg concentration, MeHg dysregulated a larger number of genes and with a stronger fold-change than IHg supporting a stronger molecular impact of the former, in line with the observed hormesis effect on photosynthesis of MeHg.
In the aquatic plant E. nuttallii. IHg and MeHg also regulated genes involved in energy metabolism, development, transport, secondary metabolism, while MeHg similarly regulated more genes than IHg, also supporting a higher molecular impact of the former than the latter (Table 1) in line with the higher internalization of MeHg during pM treatments ([7,36]). At the organism level, MeHg induced antioxidants (e.g., anthocyanin, POD), while IHg decreased chlorophyll content, supporting different cellular toxicity pathways of MeHg and IHg, but also evidenced a stronger impact of those treatments in the macrophyte than the microalgae.
Overall, both the number of regulated contigs identified by RNA-Seq and cellular endpoints pointed to different molecular toxicity of MeHg vs. IHg in C. reinhardtii and E. nuttallii. Thereafter, a comparative study of data in the microalga and the macrophyte was conducted to uncover more in details dissimilarities, as well as different sensitivities to Hg stress, to highlight potential specie-specific (e.g., morphological or phylogenetical) or common responses. Analysis of the perturbations of the cell’s functions will help to derive a detailed mechanistic understanding of differences in cellular handling of IHg and MeHg resulting in MeHg having a stronger impact on biota.
4.1. Comparison of IHg and MeHg Bioaccumulation
The intracellular IHg and MeHg concentrations were assessed in both organisms ([7,36,42,50]). Comparing the concentration in organism to concentration in media ratios, the highest values were obtained at the lower MeHg exposure concentration for both C. reinhardtii and E. nuttallii (Figure 2). This result suggests a high affinity uptake of MeHg. This uptake could be carrier mediated, as previously described in S. capricornutum at MeHg concentrations below 10 nM ([30]). Moreover, in both organisms, we found a higher proportion of adsorbed Hg for IHg than MeHg. In natural waters with low Hg concentrations, it is thus likely that the binding of IHg to DOM and cell walls significantly reduces IHg uptake compared to MeHg. We also observed that the proportions of adsorbed vs. intracellular Hg were higher for C. reinhardtii than E. nuttallii. This difference can be attributed to the unicellularity of the alga vs. the pluricellularity of the macrophyte, in which internalization to the internal cell layers may need longer exposure. Additionally, the surface-to-volume ratio is expected to be higher in the unicellular organism and, thus, to result in higher uptake ([65]). However, here intracellular concentration of Hg in E. nuttallii appeared to be higher than for C. reinhardtii (Figure 2), in line with observations in the field showing high uptake in this species and the well-known high uptake capacity of this genus for metals ([22]).
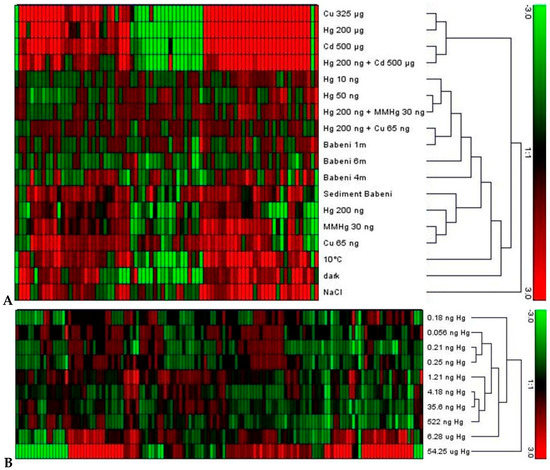
Figure 2.
(A) Clustering of differentially expressed genes as revealed by nCounter analysis. Elodea nuttallii were exposed in the laboratory to various metal concentrations (where MMHg is monomethyl-mercury), to Hg contaminated sediment collected in the Olt river in Romania (Sediment Babeni), and to other abiotic factors temperature, darkness, and salinity (NaCl). A set of plants was exposed in the Olt river at 1, 4, and 6 m water depth (Babeni 1 m, 4 m, 6 m). Hierarchical clustering with average linkage was performed using Genesis software; (B) Clustering of differentially expressed genes in response to gradually increasing effective Hg concentration in controlled conditions. Gene signature was positively correlated with Hg concentration from 1 pM (0.2 ng·L−1) (the figure was adapted from the original data and figure in [62]).
4.2. Comparison of Whole Transcriptome Analyses
The response of the whole transcriptome by RNA-Seq to exposure of the increasing IHg and MeHg concentrations was compared in C. reinhardtii and E. nuttallii. For both organisms, the number of significantly dysregulated genes was higher for MeHg than IHg at similar intracellular THg concentrations, suggesting a stronger biological response to MeHg. Therefore, here we compared the biological pathways affected by IHg and MeHg in the microalga and the aquatic plant. To this end, the number of genes in a functional category was normalized by the number of genes in the whole transcriptome. In addition, for IHg, we compared the treatments that showed the highest number of dysregulated genes, i.e., 1 nM and 10 nM IHg for E. nuttallii and C. reinhardtii, respectively. For MeHg, the pathways represented by genes that answered commonly to the four exposure concentrations in C. reinhardtii were compared to the one regulated commonly by 1 nM and 10 nM MeHg in E. nuttallii (Figure 3). The top 10 biological pathways affected by Hg species exposure (represented by ~50% of the dysregulated genes) are compared and discussed below.
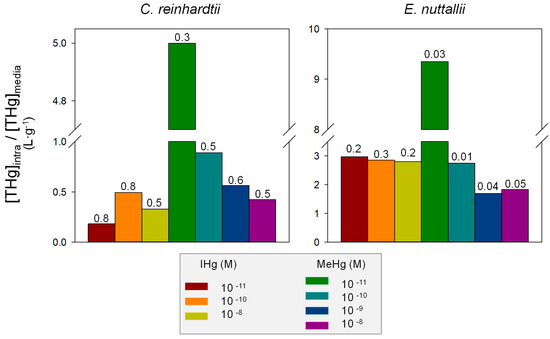
Figure 3.
Mean intracellular concentration ([THg]intra) vs. IHg and MeHg exposure concentrations ([THg]media) in C. reinhardtii and E. nuttallii. Numbers indicate the proportion of adsorbed concentration vs. total concentration (adsorbed + absorbed).
Genes involved in photosynthesis were up-regulated in response to IHg in C. reinhardtii only and to MeHg in both organisms (Figure 4). Genes involved in sugar metabolism represented one third of E. nuttallii response to both IHg (down-regulated) and MeHg (up-regulated), while in C. reinhardtii impact on the expression of genes involved in sugar metabolism was limited to down-regulated genes involved in gluconeogenesis by both IHg and MeHg exposures. Genes involved in tetrapyrrole synthesis were down-regulated in response to IHg in C. reinhardtii, while up-regulated in E. nuttallii and both up and down regulated by MeHg in E. nuttallii, but not in C. reinhardtii. Gene expression of photosynthetic genes was, thus, altered in the two organisms here, confirming a previous proteomic study in E. nuttallii exposed 24 h to 0.35 nM IHg ([33]). IHg however down-regulated genes involved in photosynthesis in E. nuttallii after 24 h exposure to 0.4 µM IHg ([40]). These apparent divergences can be attributed to different experimental conditions, including medium (artificial vs. natural lake water) and exposure length.
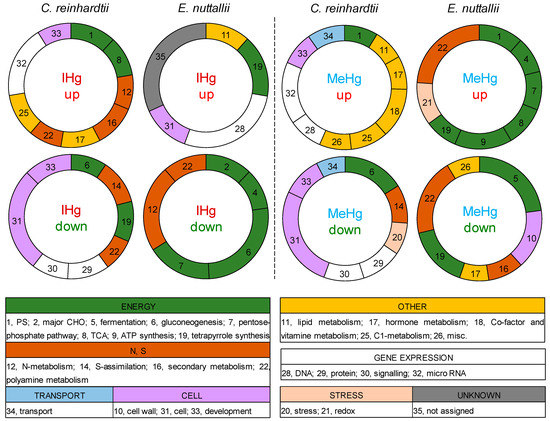
Figure 4.
Distribution of the top functional categories up- or down- regulated by IHg (10−8 M IHg for C. reinhardtii and E. nuttallii) or MeHg (common genes all MeHg treatments for C. reinhardtii, common 10−8 and 10−9 M MeHg for E. nuttallii) in C. reinhardtii or E. nuttallii. Gene numbers were normalized by the total number of genes assigned to a category in the whole transcriptome.
Genes involved in nitrogen (N) and polyamine metabolisms were dysregulated in E. nuttallii and C. reinhardtii exposed to IHg: genes involved in N-metabolism were up-regulated and genes involved in polyamine metabolism were down-regulated (Figure 4). Similar observations were found for E. nuttallii exposed to MeHg, however these categories were not significantly dysregulated in C. reinhardtii exposed to MeHg. The effect on genes involved in N-metabolism was consistent with previous experiments in E. nuttallii that showed common down-regulation of genes involved in N-metabolism (e.g., nitrite reductase, ammonia transport) after 24 h exposure to 0.4 nM, 0.4 and 4 µM IHg ([62]), suggesting a higher impact of Hg on nutrition in the macrophyte than the microalga. N homeostasis in plants as sessile organisms might be more sensitive and adaptable than in a mobile microalga.
Stress was among the 10 most enriched biological pathways for MeHg in C. reinhardtii, and E. nuttallii, but not for IHg, supporting the higher impact of MeHg on primary producers. Similarly, transport was among the 10 most enriched biological pathways for MeHg in C. reinhardtii, evidencing a strong effect of MeHg on nutrient and metal homeostasis. In addition, a down-regulation of the amino acid transporter AOT4 in response to MeHg, and many Zn transporters were dysregulated for IHg in C. reinhardtii. Zn transporters were also highly dysregulated in E. nuttallii, together with cobalt and molybdate transporters. The vacuolar proton pumps (ATPase, v-PPase) were overrepresented in up-regulated contigs by 1 and 10 µg·L−1 MeHg in the macrophyte, suggesting a vacuolar transport of MeHg for detoxification. Nevertheless, further experiments (competition, kinetics) are necessary to better understand the uptake and internalization pathways of IHg and MeHg at the subcellular level.
4.3. Comparison of Physiological Data
At the physiological level, methods used were different and do not allow direct comparison. Nevertheless, in E. nuttallii, the chlorophyll content measured by acetone extraction and spectrophotometry was reduced by IHg, but not by MeHg. In C. reinhardtii, chlorophyll content was not impacted when measured with the Trilogy Laboratory Fluorometer equipped with the chlorophyll a in vivo module (Turner Designs, Sunnyvale, CA, USA.), but we observed by fast repetition rate fluorometry (FRRf) an increased PSII efficiency linked with up-regulation of photosynthetic and chloroplastic genes for both Hg forms, suggesting a hormesis effect ([48]). While in E. nuttallii, repression of genes involved in photosynthesis and down-stream sugar metabolism is consistent with the decreased chlorophyll content. These observations support that energy metabolism disturbance was different by Hg forms in each species, and that E. nuttallii was more impacted than C. reinhardtii by treatments. The same conclusion was driven in an in situ study on C. reinhardtii and E. nuttallii conducted in the Olt River (Romania) contaminated by Hg in the pM range, because the number of regulated genes was significantly higher for the macrophyte than the microalgae after 2 h-long exposure ([6]).
Impact on the photosynthesis revealed by the transcriptomic analyses and at the physiological level can lead to an increase of intracellular ROS. Indeed, here the exposure to MeHg resulted in ROS content increase in C. reinhardtii and in an increase of POD activity, involved in anti-oxidant responses, in E. nuttallii. For both organisms, physiological evidence of MeHg induced oxidative stress followed a bell-shape, with a maximum effect at 10−10 M MeHg, suggesting that at higher concentrations, an efficient anti-oxidant response was established. Moreover, a high content of anthocyanin found in E. nuttallii exposed to MeHg as well as the expression of genes involved in metal chelation and coding for anti-oxidant enzymes, e.g., catalase and SOD in C. reinhardtii, are consistent with this hypothesis.
5. Conclusions
The present review focused on Hg effects on different level of biological organization of primary producers. The potential of transcriptomics to assess the mode of action of IHg and MeHg is illustrated comparing omics data in a microalgae and a macrophyte. Recent, research conducted on the submerged macrophyte E. nuttallii and the unicellular green alga C. reinhardtii were complementary and increased our mechanistic knowledge on IHg and MeHg cellular toxicity pathways. The work brought new insight on the toxicity targets of MeHg and the possible responses triggered, including vacuolar detoxification of MeHg, impact on photosynthesis and oxidative stress. While the measurement of intracellular concentration in the alga and the macrophyte enabled to assess the bioavailability of IHg and MeHg in our exposure medium, gene expression informed on the actual biological effect. Indeed, the interaction of Hg with biomolecules can only be assessed by measuring molecular responses. Data also showed interspecific differences in uptake, as well as in gene and physiological responses. This review highlighted the need to study more species than only well-studied model organisms. The use of omics appears as a promising endpoint to develop new biomarkers as it is sensitive and specific, but the direct application of omics datasets to risk assessments from a regulatory perspective remains a long-way goal. Indeed, a current challenge is to link the expression of genes and proteins with impacts at population levels to use these tools in an ecotoxicological context. Nonetheless, there are several knowledge gaps that can be currently addressed in terms of how omics can be used as part of identifying molecular toxicity targets and responses. The general lack of diverse experimental conditions used to produce omics data limits our ability to make robust extrapolations in support of risk assessment. For example, the outcomes of the present review and case study pointed new questions to consider for future research. Namely: (i) a better understanding of the time dependence of the gene expression should be conducted to follow the dynamics of the transcriptomic response in relation with physiological response. In the present work, the exposure was performed at one-time point although the expression of genes and proteins is known to be time dependent; therefore, future research should include more time points; (ii) gene expression analysis should be conducted in different plant compartments (e.g., leaves vs. roots) or organelles (e.g., chloroplast, mitochondria); (iii) in the same line of research, more proteomic and metabolomic analyses should be undertaken to further investigate the hypotheses elaborated based on transcriptomics ([66,67]). Moreover, methods have recently been proposed to isolate cell wall proteomes ([68,69]). Studying chloroplast transcriptomes or cell wall proteomes may help in targeting proteins involved in Hg handling in aquatic primary producers; (iv) future research should undertake similar experiments with additional species of primary producers, to confirm and confidently generalize biological pathways found here. It is expected that fundamental biological systems such as detoxification pathways to be conserved across species, but small structural, functional or life form variations can lead to substantial differences in chemical sensitivity even in conserved pathways ([70]).
Although currently, omics datasets do not provide sufficient evidence to characterize risk for an ecosystem, they improve our understanding of the molecular mechanisms involved in aquatic organisms’ response to pollutants, in particular at environmental concentrations. In coming years, those approaches are expected to broaden the research and the development concerning exposure biomarkers in aquatic primary producers to assess the health of a whole ecosystem. A better knowledge of the respective physiology of primary producers will be really useful for their further use in ecotoxicological studies. Indeed, a current research priority is to gain insight into cellular toxicity pathways of IHg and MeHg across different taxonomic groups of primary producers.
Author Contributions
R.B.-F. performed experiments and analyzed data under V.I.S. and C.C. supervision; R.B.-F., V.I.S. and C.C. wrote the text.
Funding
This research was funded by the Swiss National Science Foundation grant number 205321_138254, 200020_157173. RBF was funded by funds Constantin Topali.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Dranguet, P.; Fluck, R.; Regier, N.; Cosio, C.; Le Faucheur, S.; Slaveykova, V.I. Towards mechanistic understanding of mercury availability and toxicity to aquatic primary producers. Chimia 2014, 68, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Kocman, D.; Wilson, S.; Amos, H.; Telmer, K.; Steenhuisen, F.; Sunderland, E.; Mason, R.; Outridge, P.; Horvat, M. Toward an assessment of the global inventory of present-day mercury releases to freshwater environments. Int. J. Environ. Res. Public Health 2017, 14, 138. [Google Scholar] [CrossRef] [PubMed]
- Cossa, D.; Averty, B.; Pirrone, N. The origin of methylmercury in open Mediterranean waters. Limnol. Oceanogr. 2009, 54, 837–844. [Google Scholar] [CrossRef]
- Bravo, A.G.; Cosio, C.; Amouroux, D.; Zopfi, J.; Cheualley, P.A.; Spangenberg, J.E.; Ungureanu, V.G.; Dominik, J. Extremely elevated methyl mercury levels in water, sediment and organisms in a Romanian reservoir affected by release of mercury from a chlor-alkali plant. Water Res. 2014, 49, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Bank, M.S. Mercury in the Environment: Pattern and Process, 1st ed.; University of California Press: Berkeley, CA, USA, 2012; 358p. [Google Scholar]
- Dranguet, P.; Cosio, C.; Le Faucheur, S.; Beauvais-Fluck, R.; Freiburghaus, A.; Worms, I.A.M.; Petit, B.; Civic, N.; Docquier, M.; Slaveykova, V.I. Transcriptomic approach for assessment of the impact on microalga and macrophyte of in-situ exposure in river sites contaminated by chlor-alkali plant effluents. Water Res. 2017, 121, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Beauvais-Fluck, R.; Slaveykova, V.I.; Skyllberg, U.; Cosio, C. Molecular effects, speciation and competition of inorganic and methyl mercury in the aquatic plant Elodea nuttallii. Environ. Sci. Technol. 2018, 52, 8876–8884. [Google Scholar] [CrossRef] [PubMed]
- Powell Kipton, J.; Brown Paul, L.; Byrne Robert, H.; Gajda, T.; Hefter, G.; Sjöberg, S.; Wanner, H. Chemical speciation of environmentally significant heavy metals with inorganic ligands. Part 2005, 1, The Hg2+–Cl–, OH–, CO32–, SO42–, and PO43– aqueous systems (IUPAC Technical Report). Pure Appl. Chem. 2009. [Google Scholar] [CrossRef]
- Le Faucheur, S.; Tremblay, Y.; Fortin, C.; Campbell, P.G.C. Acidification increases mercury uptake by a freshwater alga, Chlamydomonas reinhardtii. Environ. Chem. 2011, 8, 612–622. [Google Scholar] [CrossRef]
- Bravo, A.G.; Loizeau, J.L.; Bouchet, S.; Richard, A.; Rubin, J.F.; Ungureanu, V.G.; Amouroux, D.; Dominik, J. Mercury human exposure through fish consumption in a reservoir contaminated by a chlor-alkali plant: Babeni reservoir (Romania). Environ. Sci. Pollut. Res. 2010, 17, 1422–1432. [Google Scholar] [CrossRef] [PubMed]
- Marshall, B.G.; Forsberg, B.R.; Thome-Souza, M.; Peleja, R.; Moreira, M.Z.; Freitas, C.E. Evidence of mercury biomagnification in the food chain of the cardinal tetra Paracheirodon axelrodi (Osteichthyes: Characidae) in the Rio Negro, central Amazon, Brazil. J. Fish Biol. 2016, 89, 220–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Cai, Y.; O’Driscoll, N.J. Environmental Chemistry and Toxicology of Mercury; Wiley: Hoboken, NJ, USA, 2012; p. 600. [Google Scholar]
- Crane, M.; Babut, M. Environmental quality standards for water framework directive priority substances: Challenges and opportunities. Integr. Environ. Assess. Manag. 2007, 3, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Snape, J.R.; Maund, S.J.; Pickford, D.B.; Hutchinson, T.H. Ecotoxicogenomics: The challenge of integrating genomics into aquatic and terrestrial ecotoxicology. Aquat. Toxicol. 2004, 67, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Noges, T.; Luup, H.; Feldmann, T. Primary production of aquatic macrophytes and their epiphytes in two shallow lakes (Peipsi and Vrtsjarv) in Estonia. Aquat. Ecol. 2010, 44, 83–92. [Google Scholar] [CrossRef]
- Bravo, A.G.; Loizeau, J.-L.; Dranguet, P.; Makri, S.; Björn, E.; Ungureanu, V.G.; Slaveykova, V.I.; Cosio, C. Persistent Hg contamination and occurrence of Hg methylating gene (hgcA) downstream a chlor-alkali plant in the Olt River (Romania). Environ. Sci. Pollut. Res. 2016, 23, 10529–10541. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, D.S.; Poulain, A.J. A little bit of light goes a long way: The role of phototrophs on mercury cycling. Metallomics 2014, 6, 396–407. [Google Scholar] [CrossRef] [PubMed]
- Le Faucheur, S.; Campbell, P.G.C.; Fortin, C.; Slaveykova, V.I. Interactions between mercury and phytoplankton: Speciation, bioavailability, and internal handling. Environ. Toxicol. Chem. 2014, 33, 1211–1224. [Google Scholar] [CrossRef] [PubMed]
- Cosio, C.; Flück, R.; Regier, N.; Slaveykova, V.I. Effects of macrophytes on the fate of mercury in aquatic systems. Environ. Toxicol. Chem. 2014, 33, 1225–1237. [Google Scholar] [CrossRef] [PubMed]
- Benbassat, D.; Mayer, A.M. Volatilization of mercury by algae. Physiol. Plant. 1975, 33, 128–131. [Google Scholar] [CrossRef]
- Regier, N.; Larras, F.; Bravo, A.G.; Ungureanu, V.-G.; Amouroux, D.; Cosio, C. Mercury bioaccumulation in the aquatic plant Elodea nuttallii in the field and in microcosm: Accumulation in shoots from the water might involve copper transporters. Chemosphere 2013, 90, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, G.; Bishopp, A.; Heenatigala, P.P.M.; Hu, S.; Chen, Y.; Wu, Z.; Kumar, S.; Duan, P.; Yao, L.; et al. A comparison of growth on mercuric chloride for three Lemnaceae species reveals differences in growth dynamics that effect their suitability for use in either monitoring or remediating Ecosystems Contaminated with Mercury. Front. Chem. 2018, 6, 112. [Google Scholar] [CrossRef] [PubMed]
- Lominchar, M.A.; Sierra, M.J.; Jimenez-Moreno, M.; Guirado, M.; Martin-Doimeadios, R.C.R.; Millan, R. Mercury species accumulation and distribution in Typha domingensis under real field conditions (Almaden, Spain). Environ. Sci. Pollut. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Wang, Q.; Zhang, Z.; Zhou, X. Mercury distribution along the food chain of a wetland ecosystem at Sanjiang Plain, Northeast China. Bull. Environ. Contam. Toxicol. 2017, 98, 162–166. [Google Scholar]
- Slaveykova, V.I.; Wilkinson, K.J. Physicochemical aspects of lead bioaccumulation by Chlorella vulgaris. Environ. Sci. Technol. 2002, 36, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Boudou, A.; Delnomdedieu, M.; Georgescauld, D.; Ribeyre, F.; Saouter, E. Fundamental roles of biological barriers in mercury accumulation and transfer in fresh-water ecosystems—(Analysis at organism, organ, cell and molecular-levels). Water Air Soil Pollut. 1991, 56, 807–822. [Google Scholar] [CrossRef]
- Morel, F.M.M.; Kraepiel, A.M.L.; Amyot, M. The chemical cycle and bioaccumulation of mercury. Annu. Rev. Ecol. Syst. 1998, 29, 543–566. [Google Scholar] [CrossRef]
- Mason, R.P.; Reinfelder, J.R.; Morel, F.M.M. Uptake, toxicity, and trophic transfer of mercury in a coastal diatom. Environ. Sci. Technol. 1996, 30, 1835–1845. [Google Scholar] [CrossRef]
- Moye, H.A.; Miles, C.J.; Phlips, E.J.; Sargent, B.; Merritt, K.K. Kinetics and uptake mechanisms for monomethylmercury between freshwater algae and water. Environ. Sci. Technol. 2002, 36, 3550–3555. [Google Scholar] [CrossRef] [PubMed]
- Hassett, R.; Kosman, D.J. Evidence for Cu(II) reduction as a component of copper uptake by Saccharomyces cerevisae. J. Biol. Chem. 1995, 270, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Page, M.D.; Kropat, J.; Hamel, P.P.; Merchant, S.S. Two Chlamydomonas CTR copper transporters with a novel Cys-Met motif are localized to the plasma membrane and function in copper assimilation. Plant Cell 2009, 21, 928–943. [Google Scholar] [CrossRef] [PubMed]
- Larras, F.; Regier, N.; Planchon, S.; Poté, J.; Renaut, J.; Cosio, C. Physiological and proteomic changes suggest an important role of cell walls in the high tolerance to metals of Elodea nuttallii. J. Hazard. Mater. 2013, 263 Pt 2, 575–583. [Google Scholar] [CrossRef]
- Castro, R.; Pereira, S.; Lima, A.; Corticeiro, S.; Valega, M.; Pereira, E.; Duarte, A.; Figueira, E. Accumulation, distribution and cellular partitioning of mercury in several halophytes of a contaminated salt marsh. Chemosphere 2009, 76, 1348–1355. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Gil, S.; Alvarez-Fernandez, A.; Sobrino-Plata, J.; Millan, R.; Carpena-Ruiz, R.O.; Leduc, D.L.; Andrews, J.C.; Abadia, J.; Hernandez, L.E. Complexation of Hg with phytochelatins is important for plant Hg tolerance. Plant Cell Environ. 2011, 34, 778–791. [Google Scholar] [CrossRef] [PubMed]
- Beauvais-Fluck, R.; Slaveykova, V.I.; Cosio, C. Cellular toxicity pathways of inorganic and methyl mercury in the green microalga Chlamydomonas reinhardtii. Sci. Rep. 2017, 7, 8034. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.B.; Vajpayee, P.; Tripathi, R.D.; Rai, U.N.; Kumar, A.; Singh, N.; Behl, H.M.; Singh, S.P. Mercury bioaccumulation induces oxidative stress and toxicity to submerged macrophyte Potamogeton crispus L. Bull. Environ. Contam. Toxicol. 2000, 65, 573–582. [Google Scholar] [CrossRef] [PubMed]
- Ferrat, L.; Romeo, M.; Gnassia-Barelli, M.; Pergent-Martini, C. Effects of mercury on antioxidant mechanisms in the marine phanerogam Posidonia oceanica. Dis. Aquat. Org. 2002, 50, 157–160. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-A.; Chi, W.-C.; Huang, T.-L.; Lin, C.-Y.; Quynh Nguyeh, T.T.; Hsiung, Y.-C.; Chia, L.-C.; Huang, H.-J. Mercury-induced biochemical and proteomic changes in rice roots. Plant Physiol. Biochem. 2012, 55, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Regier, N.; Beauvais-Flück, R.; Slaveykova, V.I.; Cosio, C. Elodea nuttallii exposure to mercury exposure under enhanced ultraviolet radiation: Effects on bioaccumulation, transcriptome, pigment content and oxidative stress. Aquat. Toxicol. 2016, 180, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Siebman, C.; Velev, O.; Slaveykova, V. Two-dimensional algal collection and assembly by combining AC-Dielectrophoresis with fluorescence detection for contaminant-induced oxidative stress sensing. Biosensors 2015, 5, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Patra, M.; Sharma, A. Mercury toxicity in plants. Bot. Rev. 2000, 66, 379–422. [Google Scholar] [CrossRef]
- Beauvais-Fluck, R.; Slaveykova, V.I.; Cosio, C. Effects of two-hour exposure to environmental and high concentrations of methylmercury on the transcriptome of the macrophyte Elodea nuttallii. Aquat. Toxicol. 2018, 194, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Samson, G.; Morissette, J.C.; Popovic, R. Determination of 4 apparent mercury interaction sites in photosystem-II by using a new modification of the Stern-Volmer analysis. Biochem. Biophys. Res. Commun. 1990, 166, 873–878. [Google Scholar] [CrossRef]
- Samson, G.; Popovic, R. Inhibitory effects of mercury on Photosystem-II photochemistry in Dunaliella tertiolecta under in vivo conditions. J. Photochem. Photobiol. B-Biol. 1990, 5, 303–310. [Google Scholar] [CrossRef]
- Sersen, F.; Kralova, K.; Bumbalova, A. Action of mercury on the photosynthetic apparatus of spinach chloroplasts. Photosynthetica 1998, 35, 551–559. [Google Scholar] [CrossRef]
- Singh, R.; Dubey, G.; Singh, V.; Srivastava, P.; Kumar, S.; Prasad, S. High light intensity augments mercury toxicity in cyanobacterium Nostoc muscorum. Biol. Trace Elem. Res. 2012, 149, 262–272. [Google Scholar] [CrossRef] [PubMed]
- Juneau, P.; Dewez, D.; Matsui, S.; Kim, S.-G.; Popovic, R. Evaluation of different algal species sensitivity to mercury and metolachlor by PAM-fluorometry. Chemosphere 2001, 45, 589–598. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Cabot, C.; Martos, S.; Gallego, B.; Barcelo, J. Do toxic ions induce hormesis in plants? Plant Sci. 2013, 212, 15–25. [Google Scholar] [CrossRef] [PubMed]
- Beauvais-Fluck, R.; Slaveykova, V.I.; Cosio, C. Transcriptomic and physiological responses of the green microalga Chlamydomonas reinhardtii during short-term exposure to subnanomolar methylmercury concentrations. Environ. Sci. Technol. 2016, 50, 7126–7134. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zeng, Y.; Qu, J.Y.; Wang, W.-X. Mercury effects on Thalassiosira weissflogii: Applications of two-photon excitation chlorophyll fluorescence lifetime imaging and flow cytometry. Aquat. Toxicol. 2012, 110–111, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Mortimer, D.C.; Czuba, M. Structural damage to leaf chloroplasts of Elodea densa caused by methylmercury accumulated from water. Ecotoxicol. Environ. Saf. 1982, 6, 193–195. [Google Scholar] [CrossRef]
- Küpper, H.; Küpper, F.; Spiller, M. Environmental relevance of heavy metal-substituted chlorophylls using the example of water plants. J. Exp. Bot. 1996, 47, 259–266. [Google Scholar] [CrossRef]
- Rai, P.K.; Tripathi, B.D. Comparative assessment of Azolla pinnata and Vallisneria spiralis in Hg removal from GB Pant Sagar of Singrauli Industrial region, India. Environ. Monit. Assess. 2009, 148, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Chandra, P. Bioaccumulation and toxicity of mercury in rooted-submerged macrophyte Vallisneria spiralis. Environ. Pollut. 1998, 103, 327–332. [Google Scholar] [CrossRef]
- Patra, M.; Bhowmik, N.; Bandopadhyay, B.; Sharma, A. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ. Exp. Bot. 2004, 52, 199–223. [Google Scholar] [CrossRef]
- Schaumlöffel, D. The Position of Metallomics within Other Omics Fields. In Metallomics Analytical Techniques and Speciation Methods; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 3–16. [Google Scholar]
- Schirmer, K.; Fischer, B.B.; Madureira, D.J.; Pillai, S. Transcriptomics in ecotoxicology. Anal. Bioanal. Chem. 2010, 397, 917–923. [Google Scholar] [CrossRef] [PubMed]
- Heidenreich, B.; Mayer, K.; Sandermann, H.; Ernst, D. Mercury-induced genes in Arabidopsis thaliana: Identification of induced genes upon long-term mercuric ion exposure. Plant Cell Environ. 2001, 24, 1227–1234. [Google Scholar] [CrossRef]
- Venkatachalam, P.; Srivastava, A.K.; Raghothama, K.G.; Sahi, S.V. Genes induced in response to mercury-ion-exposure in heavy metal hyperaccumulator Sesbania drummondii. Environ. Sci. Technol. 2009, 43, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Montero-Palmero, M.B.; Martín-Barranco, A.; Escobar, C.; Hernández, L.E. Early transcriptional responses to mercury: A role for ethylene in mercury-induced stress. New Phytol. 2014, 201, 116–130. [Google Scholar] [CrossRef] [PubMed]
- Regier, N.; Baerlocher, L.; Munsterkotter, M.; Farinelli, L.; Cosio, C. Analysis of the Elodea nuttallii transcriptome in response to mercury and cadmium pollution: Development of sensitive tools for rapid ecotoxicological testing. Environ. Sci. Technol. 2013, 47, 8825–8834. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Yang, Z.M. Mercury toxicity, molecular response and tolerance in higher plants. Biometals 2012, 25, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Gouveia, D.; Chaumot, A.; Charnot, A.; Almunia, C.; Francois, A.; Navarro, L.; Armengaud, J.; Salvador, A.; Geffard, O. Ecotoxico-proteomics for aquatic environmental monitoring: First in situ application of a new proteomics-based multibiomarker assay using caged amphipods. Environ. Sci. Technol. 2017, 51, 13417–13426. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, C.; Fiksen, O.; Andersen, K.H.; Aksnes, D.L. Scaling laws in phytoplankton nutrient uptake affinity. Front. Mar. Sci. 2016, 3, 26. [Google Scholar] [CrossRef]
- Hossain, Z.; Makino, T.; Komatsu, S. Proteomic study of beta-aminobutyric acid-mediated cadmium stress alleviation in soybean. J. Proteom. 2012, 75, 4151–4164. [Google Scholar] [CrossRef] [PubMed]
- Hossain, Z.; Komatsu, S. Contribution of proteomic studies towards understanding plant heavy metal stress response. Front. Plant Sci. 2013, 3, 310. [Google Scholar] [CrossRef] [PubMed]
- Feiz, L.; Irshad, M.; Pont-Lezica, R.F.; Canut, H.; Jamet, E. Evaluation of cell wall preparations for proteomics: A new procedure for purifying cell walls from Arabidopsis hypocotyls. Plant Methods 2006, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Witzel, K.; Shahzad, M.; Matros, A.; Mock, H.-P.; Mühling, K.H. Comparative evaluation of extraction methods for apoplastic proteins from maize leaves. Plant Methods 2011, 7, 48. [Google Scholar] [CrossRef] [PubMed]
- Clemens, S.; Ma, J.F. Toxic heavy metal and metalloid accumulation in crop plants and foods. Annu. Rev. Plant Biol. 2016, 67, 489–512. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).