

Evolution of Coastal Subarctic Lakes in the Context of Climatic and Geological Changes and Human Occupation (North-Central Labrador, Canada)


Abstract
1. Introduction
1.1. Human Occupation in the Context of Environmental Change
1.2. Palaeogeographic Context
1.3. Palaeoclmatic Context
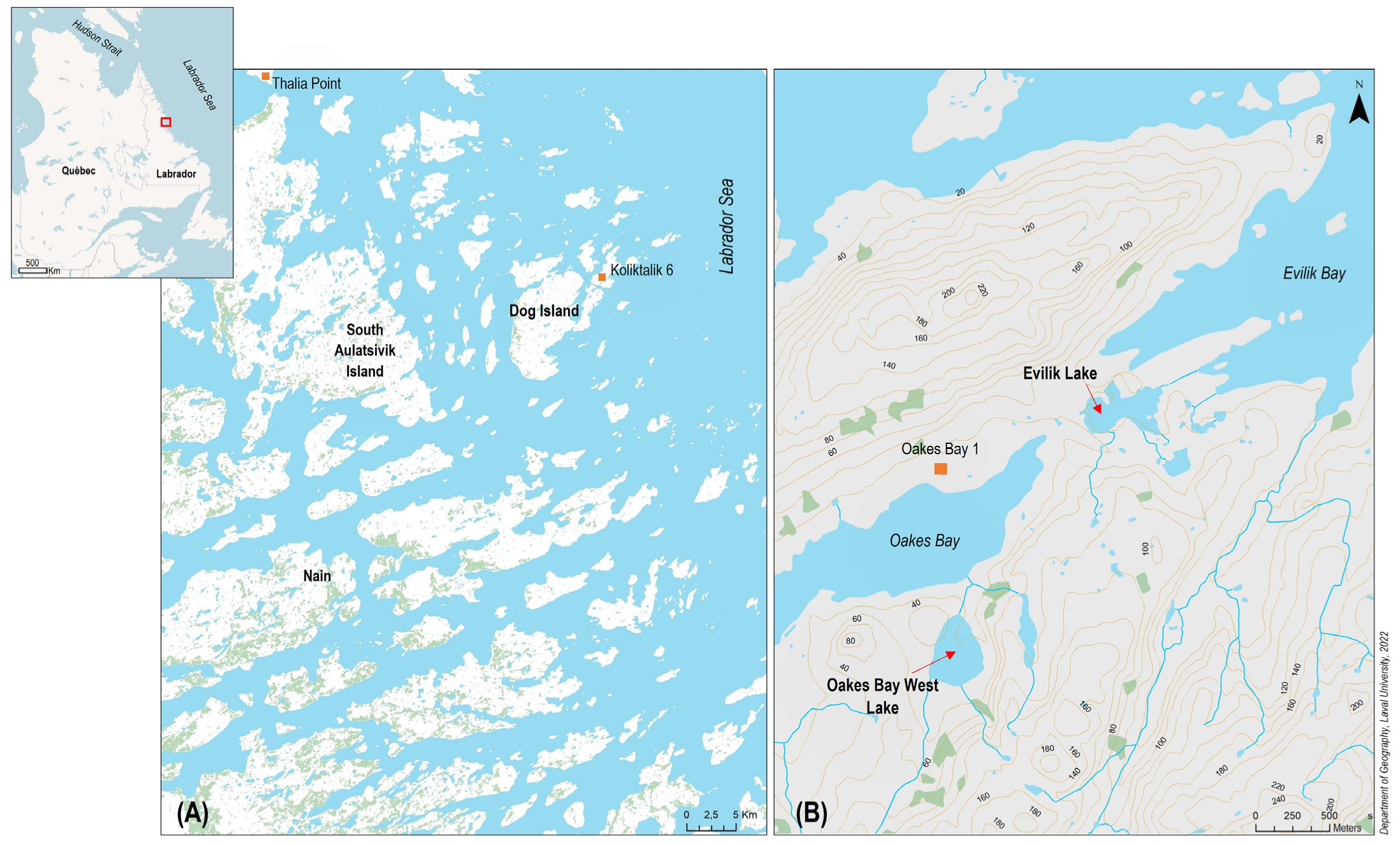
2. Study Site
3. Materials and Methods
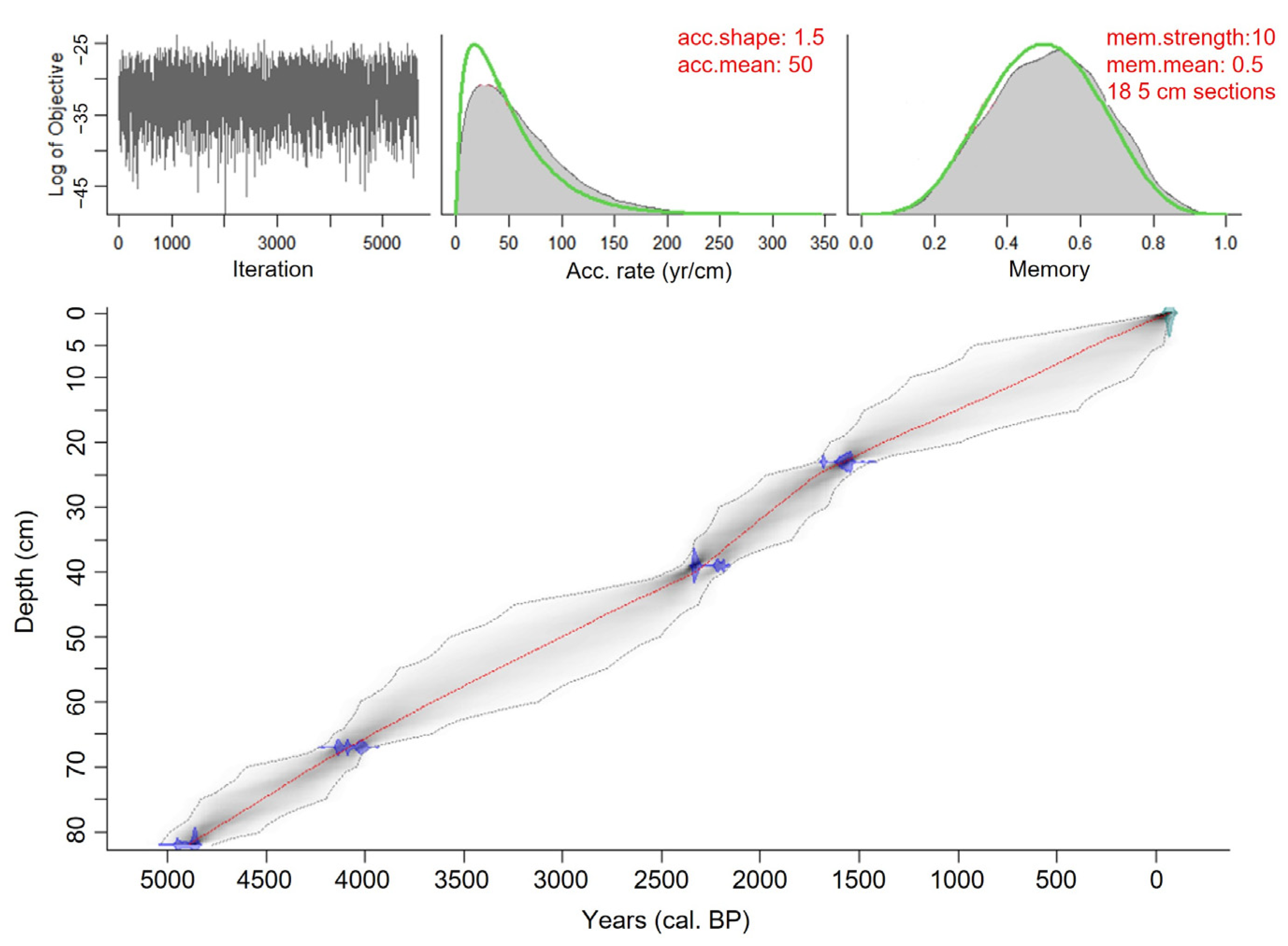
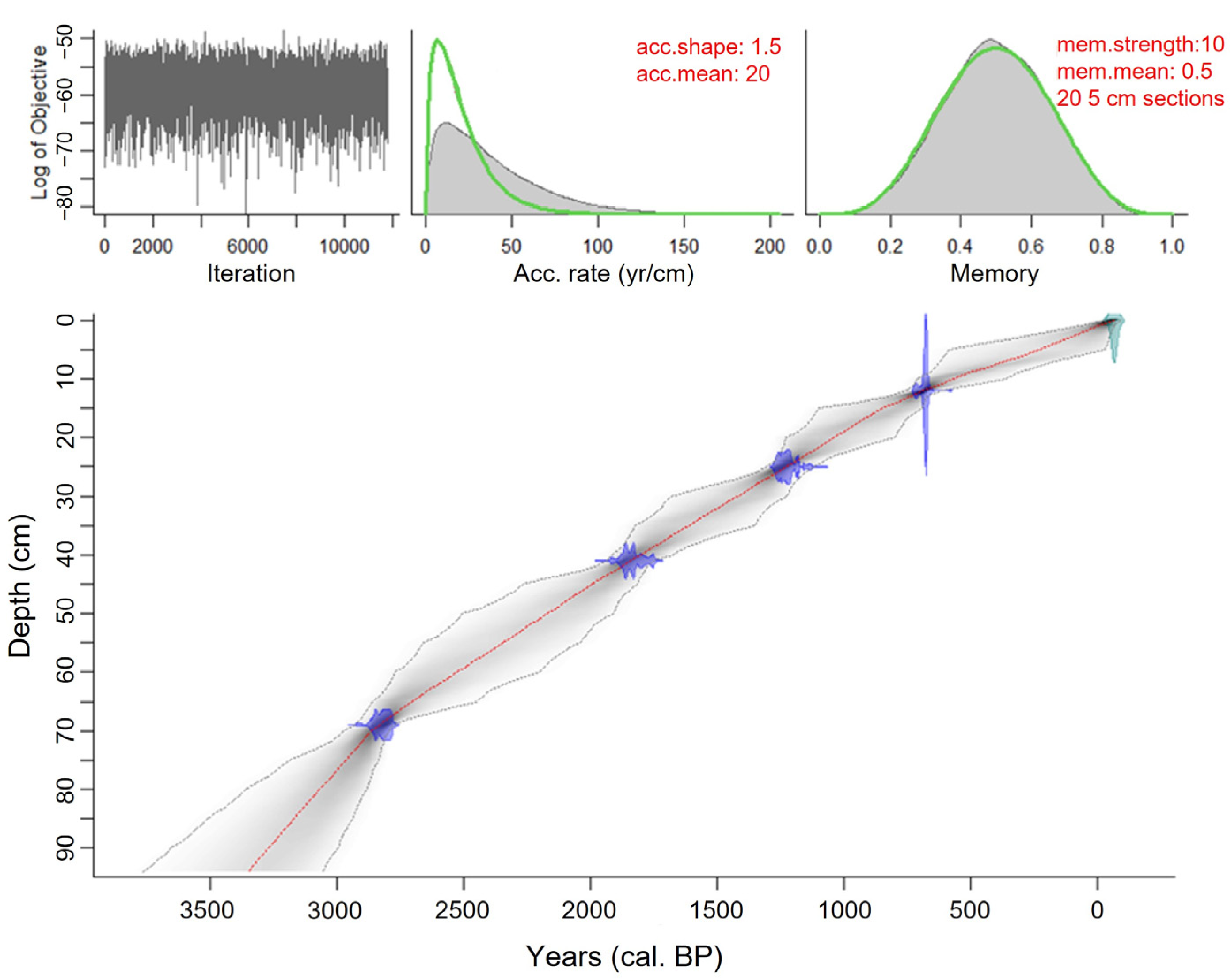
3.1. Dating and Chronology
3.2. Organic Matter and Water Content
3.3. Grain Size Analysis
3.4. Magnetic Susceptibility and Geochemical Analysis
3.5. Diatom Analyses
3.6. Numerical Analyses
4. Results
4.1. Oakes Bay West Lake
4.1.1. General Stratigraphy
4.1.2. Chronology
4.1.3. Lithology
4.1.4. Grain Size
4.1.5. Geochemical Data
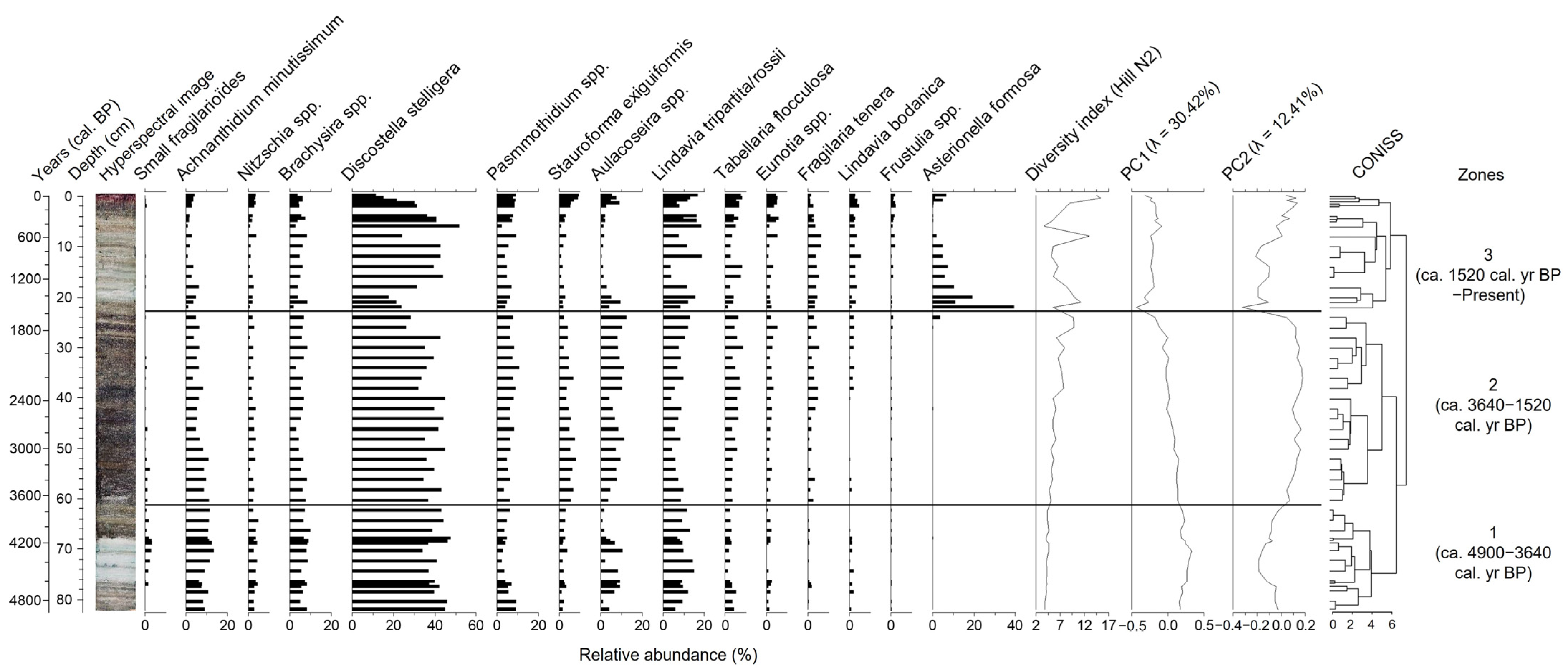
4.1.6. Diatom Data
4.2. Evilik Lake
4.2.1. General Stratigraphy
4.2.2. Chronology
4.2.3. Lithology
4.2.4. Grain Size
4.2.5. Geochemical Data
4.2.6. Diatom Data
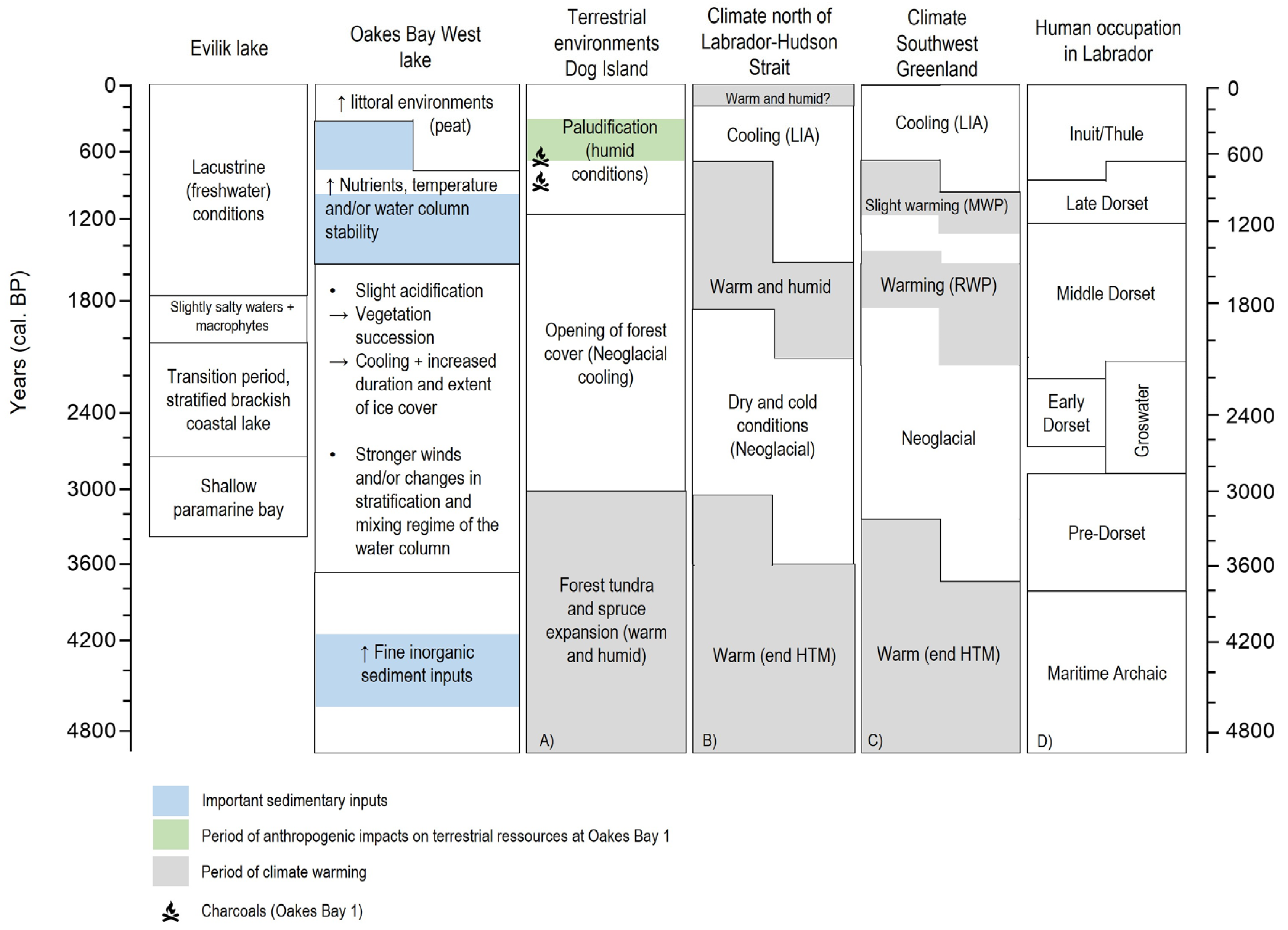
5. Discussion
5.1. Palaeoenvironmental and Palaeoclimatic Reconstruction of Oakes Bay West Lake
5.1.1. End of Holocene Thermal Maximum (ca. 4900–3640 cal. yr BP)
5.1.2. Neoglacial Period (ca. 3640–1520 cal. yr BP)
5.1.3. Roman and Medieval Warm Periods to Present Conditions (ca. 1520 cal. yr BP–Present)
5.2. Palaeogeographic Evolution of Evilik Lake
5.2.1. Glacio-Marine Environment (ca. 3350–2700 cal. yr BP)
5.2.2. Isolation Process and Brackish Conditions (ca. 2700–1990 cal. yr BP)
5.2.3. Freshwater Conditions (ca. 1990 cal. yr BP–Present)
5.3. Relative Sea Level Curves, Climate Changes, and Archaeological Implications
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smol, J.P.; Wolfe, A.P.; Birks, J.B.; Douglas, M.S.V.; Jones, V.J.; Korhola, A.; Pienitz, R.; Rühland, K.; Sorvari, S.; Antoniades, D.; et al. Climate-driven regime shifts in the biological communities of arctic lakes. Proc. Natl. Acad. Sci. USA 2005, 102, 4397–4402. [Google Scholar] [CrossRef]
- Brown, R.; Lemay, M. Variabilité et changements climatiques dans la péninsule du Nunavik et du Nunatsiavut (IRIS de la région subarctique de l’Est du Canada). In De la Science aux Politiques Publiques. Une Étude Intégrée D’impact Régional des Changements Climatiques et de la Modernisation; Allard, M., Lemay, M., Eds.; ArcticNet Inc.: Québec, QC, Canada, 2013; 318p. [Google Scholar]
- Finnis, J.; Bell, T. An analysis of recent observed climate trends and variability in Labrador. Can. Geogr. Geogr. Can. 2015, 59, 151–166. [Google Scholar] [CrossRef]
- Woollett, J. Oakes Bay 1: A Preliminary Reconstruction of a Labrador Inuit Seal Hunting Economy in the Context of Climate Change. Geogr. Tidsskr. Dan. J. Geogr. 2010, 110, 245–259. [Google Scholar] [CrossRef]
- Roy, N.; Bhiry, N.; Woollett, J. Environmental Change and Terrestrial Resource Use by the Thule and Inuit of Labrador, Canada. Geoarchaeology 2012, 27, 18–33. [Google Scholar] [CrossRef]
- Roy, N.; Woollett, J.; Bhiry, N. Paleoecological perspectives on landscape history and anthropogenic impacts at Uivak Point, Labrador, since AD 1400. Holocene 2015, 25, 1742–1755. [Google Scholar] [CrossRef]
- Roy, N.; Bhiry, N.; Woollett, J.; Delwaide, A. A 550-Year Record of the Disturbance History of White Spruce Forests Near Two Inuit Settlements in Labrador, Canada. J. North Atl. 2017, 16, 1–14. [Google Scholar] [CrossRef]
- Rosol, R.; Powell-Hellyer, S.; Man Chan, H. Impacts of decline harvest of country food on nutrient intake among Inuit in Arctic Canada: Impact of climate change and possible adaptation plan. Int. J. Circumpolar Health 2016, 75, 31127. [Google Scholar] [CrossRef] [PubMed]
- Pienitz, R.; Douglas, M.S.V.; Smol, J.P. Paleolimnological research in polar regions: An introduction. In Long-Term Environmental Change in Arctic and Antarctic Lakes; Pienitz, R., Douglas, M.S.V., Smol, J.P., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 1–17. [Google Scholar]
- Sawada, M.; Gajewski, K.; de Vernal, A.; Richard, P. Comparison of marine and terrestrial Holocene climatic reconstructions from northeastern North America. Holocene 1999, 9, 267–277. [Google Scholar] [CrossRef]
- Kaplan, M.R.; Wolfe, A.O. Spatial and temporal variability of Holocene temperature in the North Atlantic region. Quat. Res. 2006, 65, 223–231. [Google Scholar] [CrossRef]
- Smol, J.P.; Cumming, B.F. Tracking long-term changes in climate using algal indicators in lake sediments. J. Phycol. 2000, 36, 986–1011. [Google Scholar] [CrossRef]
- Fitzhugh, W. Environmental Archeology and Cultural Systems in Hamilton Inlet, Labrador: A Survey of the Central Labrador Coast from 3000 B.C. to the Present; Smithsonian Institution Press: Washington, WA, USA, 1972; 299p. [Google Scholar]
- D’Andrea, W.J.; Huang, Y.; Fritz, S.C.; Anderson, N.J. Abrupt Holocene climate change as an important factor for human migration in West Greenland. Proc. Natl. Acad. Sci. USA 2011, 108, 9765–9769. [Google Scholar] [CrossRef] [PubMed]
- Fitzhugh, W. Population movement and culture change on the central Labrador coast. Ann. N. Y. Acad. Sci. 1977, 288, 481–497. [Google Scholar] [CrossRef]
- NICH-Arctic. Available online: http://nicharctic.ca/about/ (accessed on 2 December 2022).
- Schledermann, P. The Effect of Climatic/Ecological Changes on the Style of Thule Culture Winter Dwellings. Arct. Alp. Res. 1976, 8, 37–47. [Google Scholar] [CrossRef]
- Barry, R.G.; Arundale, W.H.; Andrews, J.T.; Nichols, H.; Bradley, R.S. Environmental Change and Cultural Change in the Eastern Canadian Arctic during the Last 5000 Years. Arct. Alp. Res. 1977, 9, 193–210. [Google Scholar] [CrossRef]
- Woollett, J. Labrador Inuit subsistence in the context of environmental change: An initial landscape history perspective. Am. Anthropol. 2007, 109, 69–84. [Google Scholar] [CrossRef]
- Couture, A.; Bhiry, N.; Monette, Y.; Woollett, J. A geochemical analysis of 18thcentury Inuit communal house floors in northern Labrador. J. Archaeol. Sci. Rep. 2016, 6, 71–81. [Google Scholar] [CrossRef]
- Foury, Y. Inuit Winter Settlement Occupation at Oakes Bay 1, (HeCg-08), Labrador, Canada: Micromorphology and Zooarchaeology of Waste Disposal Sites. Master’s Thesis, Université Laval, Quebec, QC, Canada, 2017. [Google Scholar]
- Clark, P.U.; Fitzhugh, W. Late Deglaciation of the Central Labrador Coast and Its Implication for the Age of Glacial Lakes Naskaupi and McLean and for Prehistory. Quat. Res. 1990, 34, 296–305. [Google Scholar] [CrossRef]
- Vacchi, M.; Engelhart, S.E.; Nikitina, D.; Ashe, E.L.; Peltier, R.W.; Roy, K.; Kopp, R.E.; Horton, B.P. Postglacial relative sea-level histories along the eastern Canadian coastline. Quat. Sci. Rev. 2018, 201, 124–146. [Google Scholar] [CrossRef]
- AMS Dating Wood. Available online: https://www.radiocarbon.com/amsdating-wood.htm (accessed on 24 July 2022).
- Canadian Archaeological Radiocarbon Database (CARD 2.1). Available online: http://www.canadianarchaeology.ca (accessed on 24 July 2022).
- d’Arrigo, R.; Buckley, B.; Kaplan, S.; Woollett, J. Interannual to multidecadal modes of Labrador climate variability inferred from tree rings. Clim. Dyn. 2003, 20, 219–228. [Google Scholar] [CrossRef]
- Richerol, T.; Fréchette, B.; Rochon, A.; Pienitz, R. Holocene climate history of the Nunatsiavut (northern Labrador, Canada) established from pollen and dinoflagellate cyst assemblages covering the past 7000 years. Holocene 2016, 26, 44–60. [Google Scholar] [CrossRef]
- Short, S.K.; Nichols, H. Holocene Pollen Diagrams from Subarctic Labrador Ungava: Vegetanional History and Climatic Change. Arct. Antarct. Alp. Res. 1977, 9, 265–290. [Google Scholar] [CrossRef]
- Lamb, H.F. Palynological Evidence for Postglacial Change in the Position of Tree Limit in Labrador. Ecol. Monogr. 1985, 55, 241–258. [Google Scholar] [CrossRef]
- Levac, E.; de Vernal, A. Postglacial changes of terrestrial and marine environments along the Labrador coast: Palynological evidence from cores 91-045-005 and 91-045-006, Cartwright Saddle. Can. J. Earth Sci. 1997, 34, 1358–1365. [Google Scholar] [CrossRef]
- Fallu, M.-A.; Pienitz, R.; Walker, I.R.; Lavoie, M. Paleolimnology of a shrub-tundra lake and response of aquatic and terrestrial indicators to climatic change in arctic Québec, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 215, 183–203. [Google Scholar] [CrossRef]
- Saulnier-Talbot, É.; Larocque-Tobler, I.; Gregory-Eaves, I.; Pienitz, R. Response of lacustrine biota to late Holocene climate and environmental conditions in northernmost Ungava (Canada). Arctic 2015, 68, 153–168. [Google Scholar] [CrossRef]
- Laing, T.E.; Pienitz, R.; Payette, S. Evaluation of Limnological Responses to Recent Environmental Change and Caribou Activity in the Riviere George Region, Northern Quebec, Canada. Arct. Antarct. Alp. Res. 2002, 34, 454–464. [Google Scholar] [CrossRef]
- Richerol, T.; Pienitz, R.; Rochon, A. Recent anthropogenic and climatic history of Nunatsiavut fjords (Labrador, Canada). Paleoceanography 2014, 29, 869–892. [Google Scholar] [CrossRef]
- Roy, N.; Woollett, J.; Bhiry, N.; Lemus-Lauzon, I.; Delwaide, A.; Marguerie, D. Anthropogenic and climate impacts on subarctic forests in the Nain region, Nunatsiavut: Dendroecological and historical approaches. Ecoscience 2021, 28, 361–376. [Google Scholar] [CrossRef]
- Lemus-Lauzon, I.; Woollett, J.; Bhiry, N. Napâttuit: Wood use by Labrador Inuit and its impact on the forest landscape. Inuit Stud. 2012, 36, 113–137. [Google Scholar] [CrossRef]
- Lemus-Lauzon, I.; Bhiry, N.; Woollett, J. Assessing the effects of climate change and land use on northern Labrador forest stands based on paleoecological data. Quat. Res. 2016, 86, 260–270. [Google Scholar] [CrossRef]
- Wheeler, E.P., 2nd. The Nain. Okak Section of Labrador. Geogr. Rev. 1935, 25, 240–254. [Google Scholar] [CrossRef]
- Ullah, W.; Beersing, A.; Blouin, A.; Wood, C.H.; Rodgers, A. Water Resources Atlas of Newfoundland; Water Resources Division, Department of Environment and Lands, Government of Newfoundland and Labrador: St John’s, NL, Canada, 1992; pp. ii–80.
- Macdonald, G.M. Some Holocene palaeoclimatic and palaeoenvironmental perspectives on Arctic/Subarctic climate warming and the IPCC 4th Assessment Report. J. Quat. Sci. 2010, 25, 39–47. [Google Scholar] [CrossRef]
- Gibb, O.; Steinhauer, S.; Fréchette, B.; de Vernal, A.; Hillaire-Marcel, C. Diachronous evolution of sea surface conditions in the Labrador Sea and Baffin Bay since the last deglaciation. Holocene 2015, 25, 1882–1897. [Google Scholar] [CrossRef]
- Données Climatiques Historiques. Available online: https://climat.meteo.gc.ca/ (accessed on 16 July 2022).
- Payette, S.; Fortin, M.; Gamache, I. The subarctic forest-tundra: The structure of a biome in a changing climate. Bioscience 2001, 51, 709–718. [Google Scholar] [CrossRef]
- Lemus-Lauzon, I.; Bhiry, N.; Arseneault, D.; Woollett, J.; Delwaide, A. Tree-ring evidence of changes in the subarctic forest cover linked to human disturbance in northern Labrador (Canada). Écoscience 2018, 50, 135–151. [Google Scholar] [CrossRef]
- Blaauw, M.; Christen, J.A. rbacon: Age-Depth Modelling Using Bayesian Statistics. R Package Version 2.5.7. Available online: https://cran.rproject.org/web/packages/rbacon/index.html (accessed on 14 January 2021).
- Blaauw, M.; Christen, J.A. Flexible paleoclimate age-depth models using an autoregressive gamma process. Bayesian Anal. 2011, 6, 457–474. [Google Scholar] [CrossRef]
- Trachsel, M.; Telford, R.J. All age–depth models are wrong, but are getting better. Holocene 2016, 27, 860–869. [Google Scholar] [CrossRef]
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Blott, S.J.; Pye, K. GRADISTAT: A grain size distribution and statistics package for the analysis of unconsolidated sediments. Earth Surf. Process. Landf. 2001, 26, 1237–1248. [Google Scholar] [CrossRef]
- Richter, T.O.; Van der Gaast, S.; Koster, B.; Vaars, A.; Gieles, R.; de Stigter, H.C.; de Haas, H.; van Weering, T.C.E. The Avaatech XRF Core Scanner: Technical description and applications to NE Atlantic sediments. In New Techniques in Sediment Core Analysis; Rothwell, R.G., Ed.; Geological Society: London, UK, 2006; Special Publication 267; pp. 39–50. [Google Scholar]
- Weltje, G.J.; Tjallingii, R. Calibration of XRF core scanners for quantitative geochemical logging of sediment cores: Theory and application. Earth Planet. Sci. Lett. 2008, 274, 423–438. [Google Scholar] [CrossRef]
- Davies, S.J.; Lamb, H.F.; Roberts, S.J. Micro-XRF Core Scanning in Palaeolimnology: Recent Developments. In Developments in Paleoenvironmental Research: Micro-XRF Studies of Sediment Cores; Croudace, I.W., Rothwell, R.G., Eds.; Springer: Dordrecht, The Netherlands, 2015; Volume 17, pp. 189–226. [Google Scholar]
- Rothwell, R.G.; Croudace, I.W. Twenty Years of XRF Core Scanning Marine Sediments: What Do Geochemical Proxies Tell Us? In Micro-XRF Studies of Sediment Cores. Developments in Paleoenvironmental Research; Croudace, I.W., Rothwell, R.G., Eds.; Springer: Dordrecht, The Netherlands, 2015; Volume 3, pp. 25–102. [Google Scholar]
- Rioja: Analysis of Quaternary Science Data. R Package Version 0.9-21. Available online: https://cran.r-project.org/web/packages/rioja/index.html (accessed on 9 July 2021).
- Vegan: Community Ecology Package. R Package Version 2.5-7. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 9 July 2021).
- Borcard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer International Publishing: Cham, Germany, 2018; 435p. [Google Scholar]
- Scherer, R. A new method for the determination of absolute abundance of diatoms and other silt-sized sedimentary particles. J. Paleolimnol. 1994, 12, 171–179. [Google Scholar] [CrossRef]
- Battarbee, R.W.; Kneen, M.J. The use of electronically counted microspheres in absolute diatom analysis. Limnol. Oceanogr. 1982, 27, 184–188. [Google Scholar] [CrossRef]
- Krammer, K. The genus Pinnularia. In Diatoms of Europe—Diatoms of the European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2000; Volume 1, 703p. [Google Scholar]
- Krammer, K. Cymbella. In Diatoms of Europe—Diatoms of the European Inland Waters and Comparable Habitats; Lange- Bertalot, H., Ed.; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2002; Volume 3, 584p. [Google Scholar]
- Krammer, K. Cymbopleura, Delicata, Navicymbula, Gomphocymbellopsis, Afrocymbella. In Diatoms of Europe—Diatoms of the European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag, K.G.: Ruggell, Liechtenstein, 2003; Volume 4, 530p. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 1. Teil: Naviculaceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1986; 876p. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1988; 596p. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; New York, NY, USA, 1991; 576p. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. Bacillariophyceae 4. Bacillariophyceae 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema. In Süßwasserflora von Mitteleuropa; Ettl, H., Gerloff, J., Heynig, H., Mollenhauer, D., Eds.; Gustav Fischer Verlag: Stuttgart, Germany; Jena, Germany, 1991; 473p. [Google Scholar]
- Campeau, S.; Pienitz, R.; Héquette, A. Diatoms from the Beaufort Sea Coast, Southern Arctic-Ocean (Canada). Modern Analogues for Reconstructing Late Quaternary Environments and Relative Sea Levels; Cramer: Stuttgart, Germany, 1999; Bibliotheca Diatomologica; Volume 42, 244p. [Google Scholar]
- Fallu, M.-A.; Allaire, N.; Pienitz, R. Freshwater Diatoms from Northern Québec and Labrador (Canada). Species-Environment Relationships in Lakes of Boreal Forest, Forest-Tundra and Tundra Regions; Cramer: Stuttgart, Germany, 2000; Bibliotheca Diatomologica; Volume 45, 200p. [Google Scholar]
- Witkowski, A.; Lange-Bertalot, H.; Metzeltin, D. Diatom Flora of Marine Coasts; A.R.G. Gantner Verlag: Ruggell, Liechtenstein, 2000; Iconographia Diatomologica; Volume 7, 925p. [Google Scholar]
- Lange-Bertalot, H. Navicula sensu stricto, 10 genera separated from Navicula sensu lato, Frustulia. In Diatoms of Europe—Diatoms of the 90 European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag: Ruggell, Liechtenstein, 2001; Volume 2, 526p. [Google Scholar]
- Antoniades, D.A.; Hamilton, P.B.; Douglas, M.S.V.; Smol, J.P. Diatoms of North America: The Freshwater Floras of Prince Patrick, Ellef Ringnes and Northern Ellesmere Islands from the Canadian Arctic Archipelago; A.R.G. Gantner Verlag: Ruggell, Liechtenstein, 2008; Iconographia Diatomologica; Volume 17, 649p. [Google Scholar]
- Zimmermann, C.; Poulin, M.; Pienitz, R. The Pliocene-Pleistocene Freshwater Flora of Bylot Island, Nunavut, Canadian High Arctic; A.R.G. Gantner Verlag: Ruggell, Liechtenstein, 2010; Iconographia Diatomologica; Volume 21, 407p. [Google Scholar]
- Lange-Bertalot, H.; Bak, M.; Witkowski, A. Eunotia and some related genera. In Diatoms of Europe—Diatoms of the European Inland Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; A.R.G. Gantner Verlag: Ruggell, Liechtenstein, 2011; Volume 6, 747p. [Google Scholar]
- Juggins, S. Software for Ecological and Palaeoecological Data Analysis and Visualisation; C2 Version 1.7.6; University of Newcastle upon Tyne: Newcastle upon Tyne, UK, 2014. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing Version 4.1.3: Vienna, Austria, 2022. [Google Scholar]
- RStudio Team. Rstudio: Integrated Development for R; RStudio Version 2021.09.0+351, PBC: Boston, MA, USA, 2022. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Saulnier-Talbot, É.; Pienitz, R.; Stafford, T.W. Establishing Holocene sediment core chronologies for northern Ungava lakes, Canada, using humic acids (AMS 14C) and 210Pb. Quat. Geochronol. 2009, 4, 278–287. [Google Scholar] [CrossRef]
- Narancic, B.; Pienitz, R.; Chapligin, B.; Meyer, H.; Francus, P.; Guilbault, J.-P. Postglacial environmental succession of Nettilling Lake (Baffin Island, Canadian Arctic) inferred from biogeochemical and microfossil proxies. Quat. Sci. Rev. 2016, 147, 391–405. [Google Scholar] [CrossRef]
- Pienitz, R.; Smol, J.P.; MacDonald, G.M. Paleolimnological reconstruction of holocene climatic trends from two boreal treeline lakes, Northwest Territories, Canada. Arct. Antarct. Alp. Res. 1999, 31, 82–93. [Google Scholar] [CrossRef]
- Rühland, K.; Smol, J.P. Diatom shifts as evidence for recent Subarctic warming in a remote tundra lake, NWT, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 226, 1–16. [Google Scholar] [CrossRef]
- Perren, B.B.; Douglas, M.S.V.; Anderson, N.J. Diatoms reveal complex spatial and temporal patterns of recent limnological change in West Greenland. J. Paleolimnol. 2008, 42, 233–247. [Google Scholar] [CrossRef]
- Narancic, B.; Saulnier-Talbot, É.; St-Onge, G.; Pienitz, R. Diatom sedimentary assemblages and Holocene pH reconstruction from the Canadian Arctic Archipelago’s largest lake. Écoscience 2021, 28, 347–360. [Google Scholar] [CrossRef]
- Rühland, K.M.; Paterson, A.M.; Smol, J.P. Lake diatom responses to warming: Reviewing the evidence. J. Paleolimnol. 2015, 54, 1–35. [Google Scholar] [CrossRef]
- Laing, T.E.; Pienitz, R.; Smol, J.P. Freshwater diatom assemblages from 23 lakes located near Norilsk, Siberia: A comparison with assemblages from other circumpolar treeline regions. Diatom Res. 1999, 14, 285–305. [Google Scholar] [CrossRef]
- Engstrom, D.R.; Fritz, S.C.; Almendinger, J.E.; Juggins, S. Chemical and biological trends during lake evolution in recently deglaciated terrain. Nature 2000, 408, 161–166. [Google Scholar] [CrossRef]
- Saulnier-Talbot, É.; Pienitz, R.; Vincent, W. Holocene Lake succession and palaeooptics of a Subarctic lake, northern Québec, Canada. Holocene 2003, 13, 517–526. [Google Scholar] [CrossRef]
- Levac, E.; de Vernal, A.; Blake, W., Jr. Sea-surface conditions in northernmost Baffin Bay during the Holocene: Palynological evidence. J. Quat. Sci. 2001, 16, 353–363. [Google Scholar] [CrossRef]
- Kaplan, M.R.; Wolfe, A.P.; Miller, G.H. Holocene environmental variability in southern Greenland inferred from lake sediments. Quat. Res. 2002, 58, 149–159. [Google Scholar] [CrossRef]
- Møller, H.S.; Jensen, K.G.; Kuijpers, A.; Aagaard-Sørensen, S.; Seidenkrantz, M.-S.; Prins, M.; Endler, R.; Mikkelsen, N. Late-Holocene environment and climatic changes in Ameralik Fjord, southwest Greenland: Evidence from the sedimentary record. Holocene 2006, 16, 685–695. [Google Scholar] [CrossRef]
- Moros, M.; Jensen, K.; Kuijpers, A. Mid- to late-Holocene hydrological and climatic variability in Disko Bugt, central west Greenland. Holocene 2006, 16, 357–367. [Google Scholar] [CrossRef]
- Seidenkrantz, M.-S.; Aagaard-Sorensen, S.; Sulsbrueck, H.; Kuijpers, A.; Jensen, K.G.; Kunzendorf, H. Hydrography and climate of the last 4400 years in a SW Greenland fjord: Implications for Labrador Sea palaeoceanography. Holocene 2007, 17, 387–401. [Google Scholar] [CrossRef]
- Sha, L.; Jiang, H.; Seidenkrantz, M.-S.; Knudsen, K.L.; Olsen, J.; Kuijpers, A.; Liu, Y. A diatom-based sea-ice reconstruction for the Vaigat Strait Disko Bugt, West Greenland over the last 5000 yr. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 403, 66–79. [Google Scholar] [CrossRef]
- Kerwin, M.W.; Overpeck, J.T.; Webb, R.S.; Anderson, K.H. Pollen-based summer temperature reconstructions for the eastern Canadian boreal forest, subarctic, and Arctic. Quat. Sci. Rev. 2004, 23, 1901–1924. [Google Scholar] [CrossRef]
- Lochte, A.A.; Repschläger, J.; Seidenkrantz, M.-S.; Kienast, M.; Blanz, T.; Schneider, R.R. Holocene water mass changes in the Labrador Current. Holocene 2019, 29, 676–690. [Google Scholar] [CrossRef]
- Lloyd, J.M.; Kuijpers, A.; Long, A.; Moros, M.; Park, L.A. Foraminiferal reconstruction of mid to late-Holocene Ocean circulation and climate variability in Disko Bugt, West Greenland. Holocene 2007, 17, 1079–1091. [Google Scholar] [CrossRef]
- Michelutti, N.; Wolfe, A.P.; Briner, J.P.; Miller, G.H. Climatically controlled chemical and biological development in Arctic lakes. J. Geophys. Res. 2007, 112, 1–10. [Google Scholar] [CrossRef]
- Rühland, K.; Priesnitz, A.; Smol, J.P. Paleolimnological Evidence from Diatoms for Recent Environmental Changes in 50 Lakes across Canadian Arctic Treeline. Arct. Antarct. Alp. Res. 2003, 35, 110–123. [Google Scholar] [CrossRef]
- Finkelstein, S.A.; Bunbury, J.; Gajewski, K.; Wolfe, A.P.; Adams, J.K.; Delvin, J.E. Evaluating diatom-derived Holocene pH reconstructions for Arctic lakes using an expanded 171-lake training set. J. Quat. Sci. 2014, 29, 249–260. [Google Scholar] [CrossRef]
- Wolfe, A.P. Climate modulates the acidity of arctic lakes on millennial time scales. Geology 2002, 30, 215. [Google Scholar] [CrossRef]
- Saros, J.E.; Anderson, N.J. The ecology of the planktonic diatom Cyclotella and its implications for global environmental change studies. Biol. Rev. 2015, 90, 522–541. [Google Scholar] [CrossRef]
- Sivarajah, B.; Rühland, K.M.; Labaj, A.L.; Paterson, A.M.; Smol, J.P. Why is the relative abundance of Asterionella formosa increasing in a Boreal Shield lake as nutrient levels decline? J. Paleolimnol. 2016, 55, 357–367. [Google Scholar] [CrossRef]
- Paull, T.M.; Finkelstein, S.A.; Gajewski, K. Interactions between climate and landscape drive Holocene ecological change in a High Arctic lake on Somerset Island, Nunavut, Canada. Arct. Sci. 2017, 3, 17–38. [Google Scholar] [CrossRef]
- St-Onge, G.; Mulder, T.; Piper, D.J.W.; Hillaire-Marcel, C.; Stoner, J.S. Earthquake and flood-induced turbidities in the Saguenay Fjord (Québec): A Holocene paleoseismicity record. Quat. Sci. Rev. 2004, 23, 283–294. [Google Scholar] [CrossRef]
- Pienitz, R. Analyse des microrestes végétaux: Diatomées. In Écologie des Tourbières du Québec-Labrador; Payette, S., Rochefort, L., Eds.; Les Presses de l’Université Laval: Quebec City, QC, Canada, 2001; pp. 311–326. [Google Scholar]
- Vos, P.C.; de Wolf, H. Diatoms as a tool for reconstructing sedimentary environments in coastal wetlands; methodological aspects. Hydrobiologia 1993, 269/270, 285–296. [Google Scholar] [CrossRef]
- Pienitz, R.; Lortie, G.; Allard, M. Isolation of Lacustrine Basins and Marine Regression in the Kuujjuaq Area, Northern Québec, as Inferred from Diatom Analysis. Géographie Phys. Quat. 1991, 45, 155–174. [Google Scholar] [CrossRef]
- Oliva, F.; Peros, M.; Viau, A. A review of the spatial distribution of and analytical techniques used in paleotempestological studies in the western North Atlantic Basin. Prog. Phys. Geogr. 2017, 41, 171–190. [Google Scholar] [CrossRef]
- Sabbe, K.; Vyverman, W. Taxonomy, morphology and ecology of some widespread representatives of the diatom genus Opephora. Eur. J. Phycol. 1995, 30, 235–249. [Google Scholar] [CrossRef]
- Weckström, K.; Juggins, S. Coastal Diatom-Environment Relationships from the Gulf of Finland, Baltic Sea. J. Phycol. 2006, 42, 21–35. [Google Scholar] [CrossRef]
- Sawai, Y.; Horton, B.P.; Kemp, A.C.; Hawkes, A.D.; Nagumo, T.; Nelson, A.R. Relationships between diatoms and tidal environments in Oregon and Washington, USA. Diatom Res. 2016, 31, 17–38. [Google Scholar] [CrossRef]
- Cuven, S.; Francus, P.; Lamoureux, S. Mid to Late Holocene hydroclimatic and geochemical records from the varved sediments of East Lake, Cape Bounty, Canadian High Arctic. Quat. Sci. Rev. 2011, 30, 2651–2665. [Google Scholar] [CrossRef]
- Denys, L. Diatom assemblages along a former intertidal gradient: A palaeoecological study of a subboreal clay layer (western coastal plain, Belgium). Neth. J. Aquat. Ecol. 1994, 28, 85–96. [Google Scholar] [CrossRef]
- Pearce, C.; Weckström, K.; Sha, L.; Miettinen, A.; Seidenkrantz, M. The Holocene marine diatom flora of Eastern Newfoundland bays. Diatom Res. 2014, 29, 441–454. [Google Scholar] [CrossRef]
- Oksman, M.; Juggins, S.; Miettinen, A.; Witkowski, A.; Weckström, K. The biogeography and ecology of common diatom species in the northern North Atlantic, and their implications for paleoceanographic reconstructions. Mar. Micropaleontol. 2019, 148, 1–28. [Google Scholar] [CrossRef]
- Balascio, N.L.; Zhang, Z.; Bradley, R.S.; Perren, B.; Dahl, S.O.; Bakke, J. A multiproxy approach to assessing isolation basin stratigraphy from the Lofoten Islands, Norway. Quat. Res. 2011, 75, 288–300. [Google Scholar] [CrossRef]
- Retelle, M.J.; Bradley, R.S.; Stuckenrath, R. Relative Sea Level Chronology Determined from Raised Marine Sediments and Coastal Isolation Basins, Northeastern Ellesmere Island, Arctic Canada. Arct. Alp. Res. 1989, 21, 113–125. [Google Scholar] [CrossRef]
- Saulnier-Talbot, É.; Pienitz, R. Isolation au postglaciaire d’un bassin côtier près de Kuujjuaraapik-Whapmagoostui, en Hudsonie (Québec): Une analyse biostratigraphique diatomifère. Géographie Phys. Quat. 2001, 55, 63–74. [Google Scholar] [CrossRef]
- Kasper, J.N.; Allard, M. Late-holocene climatic changes as detected by the growth and decay of ice wedges on the southern shore of Hudson strait, northern Québec, Canada. Holocene 2001, 11, 563–577. [Google Scholar] [CrossRef]
- Ouzilleau Samson, D.; Bhiry, N.; Lavoie, M. Late-Holocene palaeoecology of a polygonal peatland on the south shore of Hudson Strait, northern Québec, Canada. Holocene 2010, 20, 525–536. [Google Scholar] [CrossRef]
- Köster, D.; Pienitz, R.; Wolfe, B.B.; Barry, S.; Foster, D.R.; Dixit, S.S. Paleolimnological assessment of human-induced impacts on Walden Pond (Massachusetts, USA) using diatoms and stable isotopes. Aquat. Ecosyst. Health Manag. 2005, 8, 117–131. [Google Scholar] [CrossRef]
- Hausmann, S.; Pienitz, R. Seasonal water chemistry and diatom changes in six boreal lakes of the Laurentian Mountains (Québec, canada): Impacts of climate and timber harvesting. Hydrobiol. Int. J. Aquat. Sci. 2009, 635, 1–14. [Google Scholar] [CrossRef]
- Douglas, M.S.V.; Smol, J.P.; Savelle, J.M.; Blais, J.M. Prehistoric Inuit whalers affected arctic freshwater ecosystems. Proc. Natl. Acad. Sci. USA 2004, 101, 1613–1617. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Oakes Bay West | Evilik |
|---|---|---|
| Coordinates | 56°39′24.188″ N 61°8′47.544″ W | 56°39′53.762″ N 61°7′05.19″ W |
| Altitude (m asl) | 40 | 14 |
| Maximum depth (m) | 23.4 | 6.2 |
| Area (km2) | 0.11 | 0.12 |
| pH | 6.83 | 7.13 |
| Temperature surface (°C) | 10.34 | 12.2 |
| Temperature bottom (°C) | 5.32 | 9.8 |
| Conductivity (µS/cm) | 53.4 | 71.12 |
| * DO (mg/L) | 12.83 | 11.35 |
| * DOC (mg/L) | 2.4 | 3 |
| * DIC (mg/L) | 1.5 | 1.9 |
| Lab Number | ID | Depth (cm) | Material | Conventional Radiocarbon Age (14C Year BP) | Calibrated 2σ Age Interval (cal. Year BP) | Calibrated Median/ Mean Probability (cal. Year BP) |
|---|---|---|---|---|---|---|
| Oakes Bay West Lake | ||||||
| ULA-9764 | OB13 | 13–13.5 | Bulk sediment | 1645 ± 15 | 1418–1568 | 1530/1520 * |
| ULA-9774 | OB23 | 23–23.5 | Bulk sediment | 1680 ± 20 | 1531–1689 | 1570/1580 |
| ULA-9775 | OB39 | 39–39.5 | Bulk sediment | 2280 ± 15 | 2181–2345 | 2330/2300 |
| ULA-9776 | OB67 | 67–67.5 | Bulk sediment | 3730 ± 15 | 3987–4150 | 4080/4070 |
| ULA-9777 | OB76 | 76–76.5 | Bulk sediment | 5320 ± 20 | 6001–6192 | 6080/6090 * |
| ULA-8952 | OB82 | 81.5–82 | Bulk sediment | 4320 ± 20 | 4840–4960 | 4860/4880 |
| Evilik Lake | ||||||
| ULA-9778 | EL12 | 12–12.5 | Bulk sediment | 745 ± 15 | 663–690 | 680/680 |
| ULA-9779 | EL25 | 25–25.5 | Bulk sediment | 1265 ± 15 | 1173–1275 | 1230/1230 |
| ULA-9763 | EL41 | 41–41.5 | Bulk sediment | 1920 ± 15 | 1747–1888 | 1840/1840 |
| ULA-9762 | EL69 | 69–69.5 | Bulk sediment | 2740 ± 15 | 2779–2865 | 2820/2820 |
| ULA-8951 | EL93 | 92.5–93 | Bulk sediment | 2060 ± 20 | 1942–2101 | 2020/2010 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latourelle-Vigeant, C.; Pienitz, R.; Bhiry, N. Evolution of Coastal Subarctic Lakes in the Context of Climatic and Geological Changes and Human Occupation (North-Central Labrador, Canada). Geosciences 2023, 13, 97. https://doi.org/10.3390/geosciences13040097
Latourelle-Vigeant C, Pienitz R, Bhiry N. Evolution of Coastal Subarctic Lakes in the Context of Climatic and Geological Changes and Human Occupation (North-Central Labrador, Canada). Geosciences. 2023; 13(4):97. https://doi.org/10.3390/geosciences13040097
Chicago/Turabian StyleLatourelle-Vigeant, Camille, Reinhard Pienitz, and Najat Bhiry. 2023. "Evolution of Coastal Subarctic Lakes in the Context of Climatic and Geological Changes and Human Occupation (North-Central Labrador, Canada)" Geosciences 13, no. 4: 97. https://doi.org/10.3390/geosciences13040097
APA StyleLatourelle-Vigeant, C., Pienitz, R., & Bhiry, N. (2023). Evolution of Coastal Subarctic Lakes in the Context of Climatic and Geological Changes and Human Occupation (North-Central Labrador, Canada). Geosciences, 13(4), 97. https://doi.org/10.3390/geosciences13040097