

A Middle Permian Oasis for Vertebrate and Invertebrate Life in a High-Energy Fluvial Palaeoecosystem of Southern Gondwana (Karoo, Republic of South Africa)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
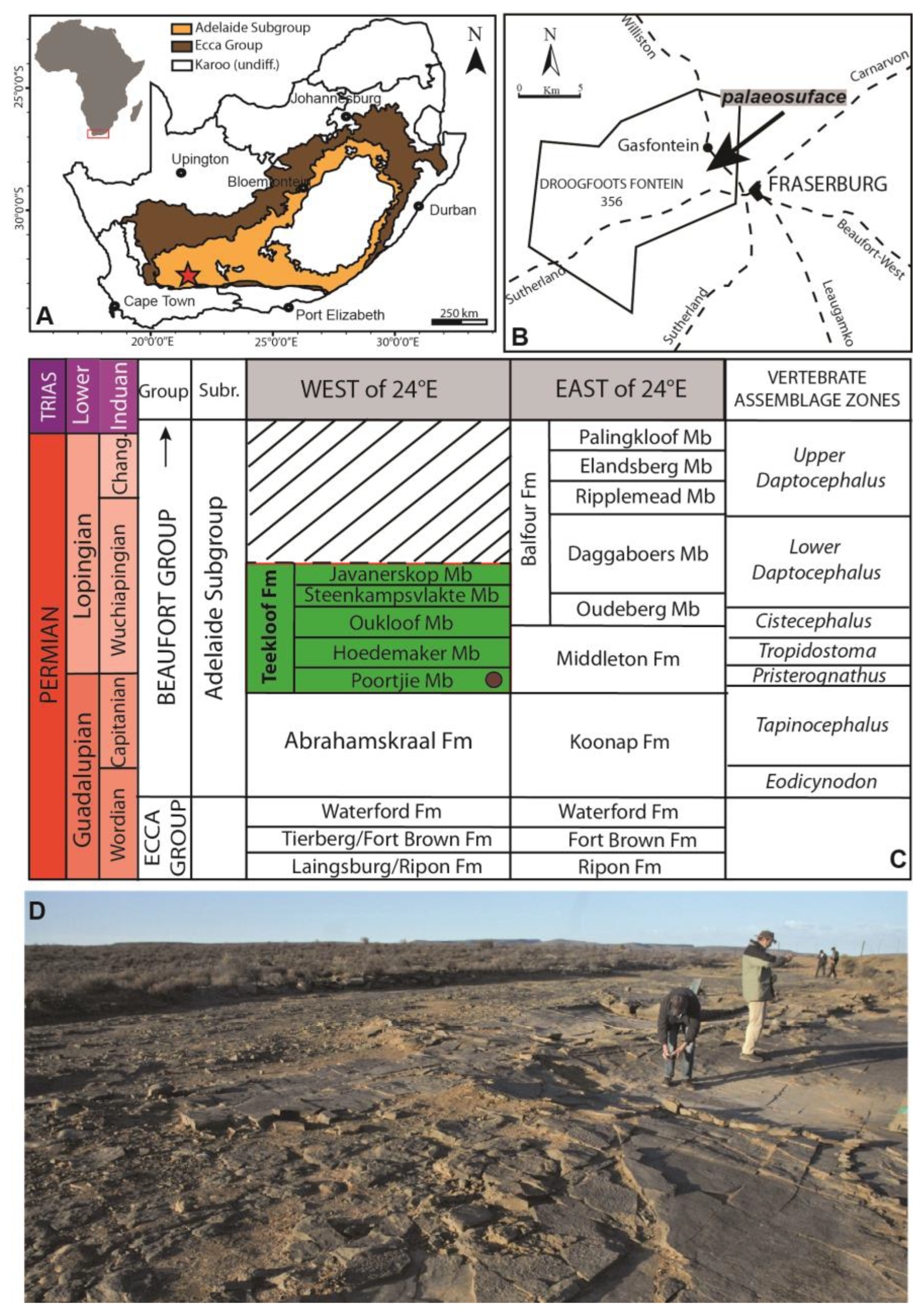
3. Geological Setting
4. The Gansfontein Palaeosurface
5. Results
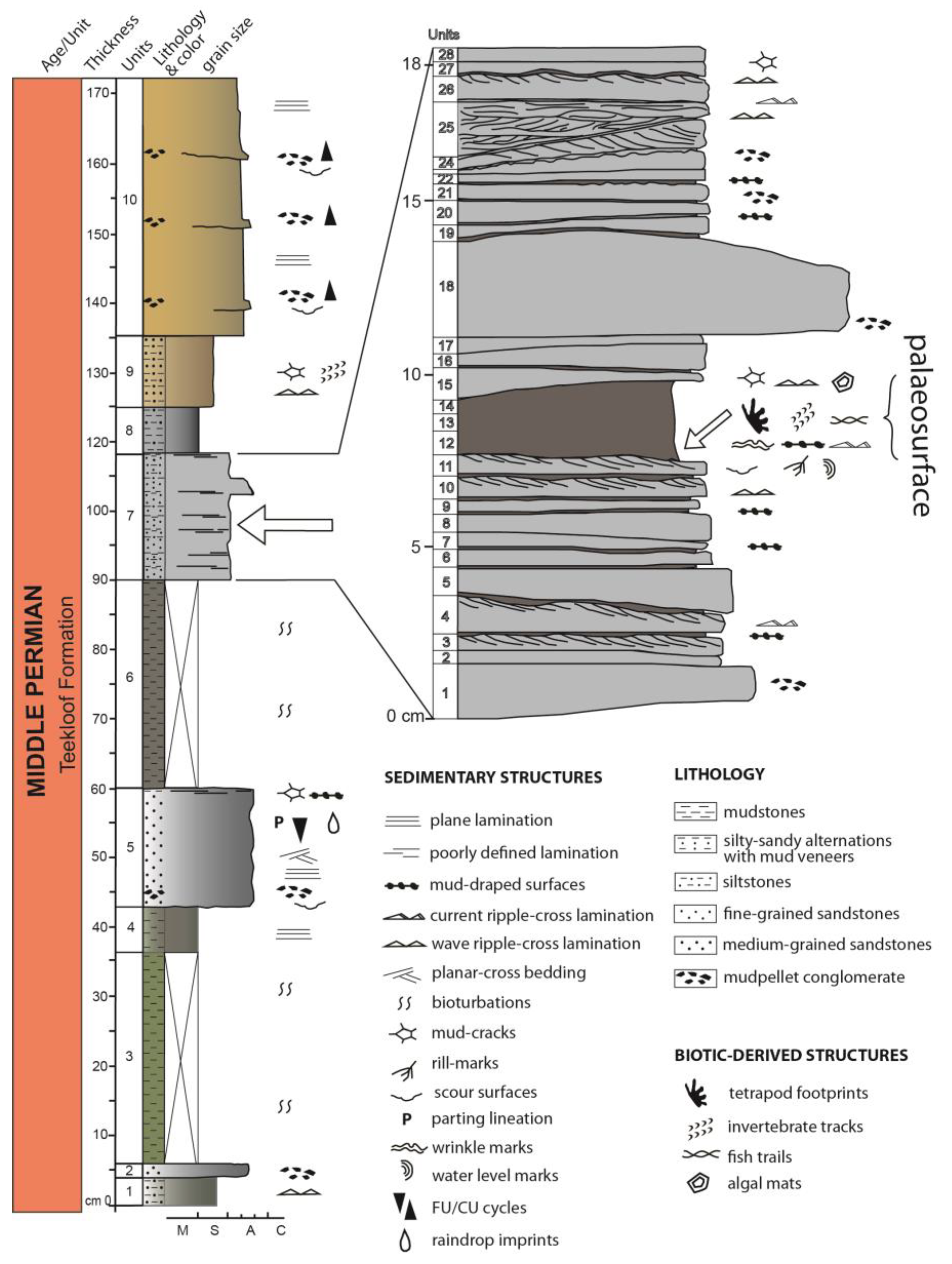
5.1. Lithostratigraphy
- Unit 1: 4 cm thick dark grey, greenish to silver-colored, massive siltstone layer with straight to sinuous wave ripples;
- Unit 2: 2 cm thick grey-colored, medium coarse sandstones with sparse mud pellets;
- Unit 3: 30 cm thick dark olive-green-colored, finely laminated, sub-covered mudstones;
- Unit 4: 7 cm thick dark gray-greenish-colored, laminated mudstones;
- Unit 5: 17 cm thick, well-cemented, FU (fining upwards) grey-colored, medium-grained sandstones with erosive base and parallel to low angle ripple cross-lamination; mud pellet layer in the upper part and mud-cracked veneer at the top; raindrop-like imprints also recorded;
- Unit 6: about 30 cm thick sub-covered, dark grey-greenish-colored, fissile mudstones;
- Unit 7: about 18 cm thick, massive to finely laminated, grey-colored, very fine sandstones/siltstones with localized dark grey-colored mud veneers. In the first 7 cm from the base, a fine repetition of siltstones/mudstones occurs. Above, a thin mud veneer (about 3 cm) belonging to the middle part of this unit represents the main palaeosurface layer: it allows the preservation of vertebrate and invertebrate traces, several types of sedimentary structures like different kinds of ripples, mud-cracks, algal mats, rill marks, scour marks; in the last 9 cm thick dark gray-colored fine sandstones interspaced with fine siltstones and mm-scale mud surfaces occur again; a wave ripples layer and very large mud cracks at the top; abundant green-colored mud chips layers occur laterally;
- Unit 8: about 6 cm thick, fissile and cracked, dark grey-colored siltstones/mudstones with mud cracks; sparse calcareous nodules;
- Unit 9: about 10 cm thick grey to light brown-colored, massive, sandy siltstones with mud-chip conglomerate at the base, ripples, mud cracks, and invertebrate traces;
- Unit 10: about 37 cm thick yellowish to light green-colored, medium-grained, well-cemented, and sorted feldspathic sandstones with ripple cross-lamination; repeated occurrence of shallow scour surfaces with mud chips, particularly occurring at the base of the unit under even lamination. Such channel sandstones frequently show a lag represented by a mud pellet conglomerate. These intraformational lenticular and thin conglomerates characteristically line channel bottoms on the basal scoured surfaces of cyclothems [52].
5.2. Sedimentary Structures
5.3. Vertebrate Trace Fossils
5.3.1. Tetrapod Footprints
5.3.2. Fish Trails
5.3.3. Invertebrate Trace Fossils
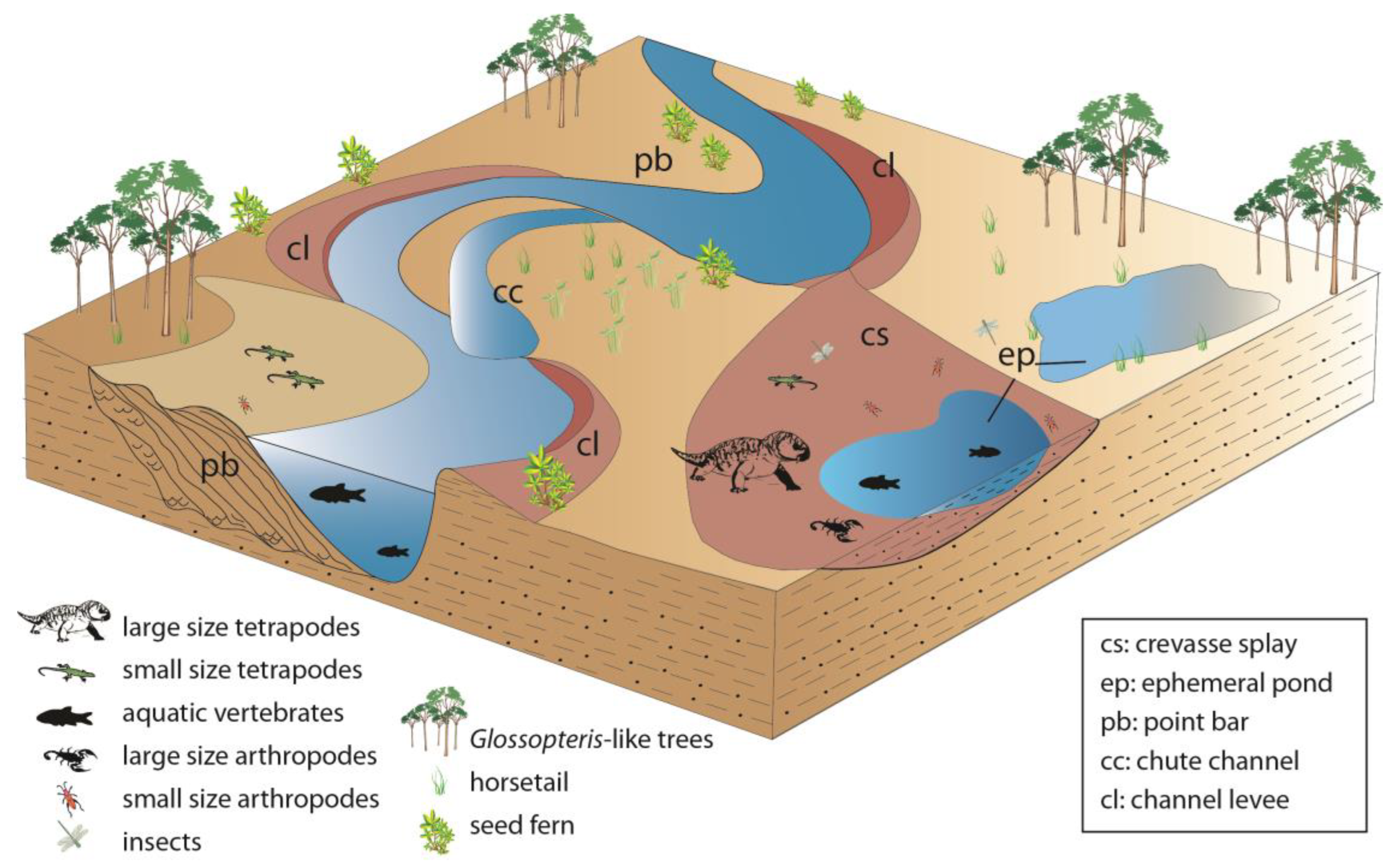
6. Discussion
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, A.M. Fish trails from the Early Permian of South Africa. Palaeontology 1976, 19, 397–409. [Google Scholar]
- Anderson, A.M. The Umfolozia arthropod trackways in the Permian Dwyka and Ecca series of South Africa. J. Paleont. 1981, 55, 84–108. [Google Scholar]
- De Beer, C.H. Surface markings, reptilian footprints and trace fossils on a palaeosurface in the Beaufort Group near Fraserburg, C.P. Ann. Geol. Surv. S. Afr. 1986, 20, 129–140. [Google Scholar]
- Smith, R.M.H. Sedimentology and Ichnology of Floodplain Palaeosurfaces in the Beaufort Group (Late Permian), Karoo Sequence, South Africa. Palaios 1993, 8, 339–357. [Google Scholar] [CrossRef]
- de Klerk, W.J. A dicynodont trackway from the Cistecephalus assemblage zone in the Karoo, east of Graaff-Reinet, South Africa. Paleont. Afr. 2002, 38, 73–91. [Google Scholar]
- Bordy, E.M.; Linkermann, S.; Prevec, R. Palaeoecological aspects of some invertebrate trace fossils from the mid-to upper Permian Middleton formation (Adelaide Subgroup, Beaufort group, Karoo Supergroup), eastern Cape, South Africa. J. Afr. Earth Sci. 2011, 61, 238–244. [Google Scholar] [CrossRef]
- Marchetti, L.; Klein, H.; Buchwitz, M.; Ronchi, A.; Smith, R.; deKlerk, W.; Sciscio, L.; Groenwald, G. Permian-Triassic vertebrate footprints from South Africa: Ichnotaxonomy, producers and biostratigraphy through two major faunal crises. Gondw. Res. 2019, 72, 139–168. [Google Scholar] [CrossRef]
- Bordy, E.M. Field Guide: A guide to the ichnology and geology of the Main Karoo Basin South Africa and Lesotho. In Proceedings of the 2nd International Conference on Continental Ichnology, Western Cape Winelands, South Africa, 1–8 October 2017. [Google Scholar]
- Buatois, L.A.; Mángano, M.G. Ichnology: Organism-Substrate Interactions in Space and Time; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Buatois, L.A.; Wisshak, M.; Wilson, M.A.; Mángano, M.G. Categories of architectural designs in trace fossils: A measure of ichnodisparity. Earth-Sci. Rev. 2017, 164, 102–181. [Google Scholar] [CrossRef]
- Marchetti, L.; Belvedere, M.; Voigt, S.; Klein, H.; Castanera, D.; Díaz-Martínez, I.; Farlow, J.O. Defining the morphological quality of fossil footprints. Problems and principles of preservation in tetrapod ichnology with examples from the Palaeozoic to the present. Earth-Sci. Rev. 2019, 193, 109–145. [Google Scholar] [CrossRef]
- Neveling, J. Stratigraphic and sedimentological investigation of the contact between the Lystrosaurus and the Cynognathus Assemblage Zones (Beaufort Group: Karoo Supergroup). Council Geosci. Bull. 2004, 137, 164. [Google Scholar]
- Rubidge, B.S. 27th Du Toit Memorial Lecture: Re-uniting lost continents—Fossil reptiles from the ancient Karoo and their wanderlust. S. Afr. J. Geol. 2005, 108, 135–172. [Google Scholar] [CrossRef]
- Smith, R.M.; Ward, P.D. Pattern of vertebrate extinctions across an event bed at the Permian-Triassic boundary in the Karoo Basin of South Africa. Geology 2001, 29, 1147–1150. [Google Scholar] [CrossRef]
- Retallack, G.J.; Metzger, C.A.; Greaver, T.; Jahren, A.H.; Smith, R.M.; Sheldon, N.D. Middle-Late Permian mass extinction on land. GSA Bull. 2006, 118, 1398–1411. [Google Scholar] [CrossRef]
- Coney, L.; Reimold, W.U.; Hancox, P.J.; Mader, D.; Koeberl, C.; McDonald, I.; Struck, U.; Vajda, V.; Kamo, S.L. Geochemical and mineralogical investigation of the Permian–Triassic boundary in the continental realm of the southern Karoo Basin. South Africa. Palaeoworld 2007, 16, 67–104. [Google Scholar] [CrossRef]
- Fildani, A.; Weislogel, A.; Drinkwater, N.J.; McHargue, T.; Tankard, A.; Wooden, J.; Hodgson, D.; Flint, S. U-Pb zircon ages from the southwestern Karoo Basin, South Africa— implications for the Permian-Triassic boundary. Geology 2009, 37, 719–722. [Google Scholar] [CrossRef]
- Lanci, L.; Tohver, E.; Wilson, A.; Flint, S. Upper Permian magnetic stratigraphy of the lower Beaufort group, Karoo basin. Earth Plan. Sci. Lett. 2013, 375, 123–134. [Google Scholar] [CrossRef]
- Rubidge, B.S.; Erwin, D.H.; Ramezani, J.; Bowring, S.A.; de Klerk, W.J. High-precision temporal calibration of Late Permian vertebrate biostratigraphy: U-Pb zircon constraints from the Karoo Supergroup, South Africa. Geology 2013, 41, 363–366. [Google Scholar] [CrossRef]
- Gastaldo, R.A.; Kamo, S.L.; Neveling, J.; Geissman, J.W.; Bamford, M.; Looy, C.V. Is the vertebrate-defined Permian-Triassic boundary in the Karoo Basin, South Africa, the terrestrial expression of the end-Permian marine event? Geology 2015, 43, 939–942. [Google Scholar] [CrossRef]
- Day, M.O.; Ramezani, J.; Bowring, S.A.; Sadler, P.M.; Erwin, D.H.; Abdala, F.; Rubidge, B.S. When and how did the terrestrial mid-Permian mass extinction occur? Evidence from the tetrapod record of the Karoo Basin, South Africa. Proc. Royal Soc. B 2015, 282, 20150834. [Google Scholar] [CrossRef]
- Day, M.O.; Ramezani, J.F.; Ryan, E.; Rubidge, B.S. U-Pb zircon age constraints on the vertebrate assemblages and palaeomagnetic record of the Guadalupian Abrahamskraal Formation, Karoo Basin, South Africa. J. Afr. Earth Sci. 2022, 186. [Google Scholar] [CrossRef]
- Turner, B.R. Uranium mineralization in the Karoo Basin, South Africa. Economic Geol. 1985, 80, 256–269. [Google Scholar] [CrossRef]
- Smith, R.M.H. Morphology and depositional history of exhumed Permian point-bars in the southwestern Karoo, South Africa. J. Sediment. Petrol. 1987, 57, 19–29. [Google Scholar]
- Smith, R.M.H. Alluvial palaeosols and pedofacies sequences in the Permian Lower Beaufort of the southwestern Karoo Basin, South Africa. J. Sediment. Petrol. 1990, 60, 258–276. [Google Scholar]
- Stear, W.M. Morphological characteristics of ephemeral stream channel and overbank splay sandstone bodies in the Permian Lower Beaufort Group, Karoo Basin, South Africa. In Modern and Ancient Fluvial Systems; Collinson, J.D., Lewin, J., Eds.; International Association of Sedimentologists IAS: Princeton, NJ, USA, 1983; Volume 6, pp. 405–420. [Google Scholar]
- Smith, R.M.H. Changing fluvial environments across the Permian-Triassic boundary in the Karoo Basin, South Africa and possible causes of tetrapod extinctions Palaeogeogr. Palaeoclimatol. Palaeoecol. 1995, 117, 81–104. [Google Scholar] [CrossRef]
- Smith, R.M.H. Sedimentology and Taphonomy of the Lower Beaufort Strata Near Beaufort West, Cape Province. Master’s Thesis, University of Witwatersrand, Johannesburg, South Africa, 1981; p. 126. [Google Scholar]
- Loock, J.C.; Brynard, H.J.; Heard, R.G.; Kitchin, J.W.; Rubidge, B.S. The stratigraphy of the Lower Beaufort Group in an area north of Laingsburg, South Africa. J. Afr. Earth Sci. 1994, 18, 185–195. [Google Scholar] [CrossRef]
- Johnson, M.R.; Van Vuuren, C.J.; Visser, J.N.J.; Cole, D.I.; Wickens, H.D.V.; Christie, A.D.M.; Roberts, D.L. The Foreland Karoo Basin, South Africa. In African Basins. Sedimentary Basins of the World, 3; Selly, R.C., Ed.; Elsevier Science B.V: Amsterdam, The Netherlands, 1997; pp. 269–317. [Google Scholar]
- Johnson, M.R.; Van Vauuren, C.J.; Visser, J.N.J.; Cole, D.I.; de Wickens, H.; Christie, A.D.M.; Roberts, D.L.; Brandl, G. Sedimentary Rocks of the Karoo Supergroup. In The Geology of South Africa; Johnson, M.R., Anhaeusser, C.R., Thomas, R.J., Eds.; Geological Society of South Africa, Johannesburg/Council for Geoscience: Pretoria, South Africa, 2006; pp. 461–501. [Google Scholar]
- Keyser, A.W.; Smith, R.H.M. Vertebrate biozonation of the Beaufort Group with special reference to the western Karoo basin. Ann. Geol. Surv. South Africa 1979, 12, 1–3. [Google Scholar]
- Mason, R.; Rubidge, B.; Hancox, J. Terrestrial vertebrate colonisation and the Ecca-Beaufort boundary in the southeastern main Karoo Basin, South Africa: Implications for Permian Basin evolution. S. Afr. J. Geol. 2015, 118, 145–156. [Google Scholar] [CrossRef]
- Rubidge, B.S.; Hancox, P.J.; Catuneanu, O. Sequence analysis of the Ecca-Beaufort contact in the southern Karoo of South Africa. S. Afr. J. Geol. 2000, 103, 81–96. [Google Scholar] [CrossRef]
- Smith, R.M.H. A review of stratigraphy and sedimentary environments of the Karoo Basin of South Africa. J. Afr. Earth Sci. 1990, 10, 117–137. [Google Scholar] [CrossRef]
- Le Roux, P.J. Palaeochannels and Uranium Mineralisation in the Main Karoo Basin of South Africa. Ph.D. Thesis, University of Port Elizabeth, Port Elizabeth, South Africa, 1985; p. 250. [Google Scholar]
- Jirah, S.; Rubidge, B.S. Refined stratigraphy of the middle Permian Abrahamskraal Formation (Beaufort Group) in the southern Karoo basin. J. Afr. Earth Sci. 2014, 100, 121–135. [Google Scholar] [CrossRef]
- Cole, D.I.; Johnson, M.R.; Day, M.O. Lithostratigraphy of the Abrahamskraal Formation (Karoo Supergroup), South Africa. S. Afr. J. Geol. 2016, 119, 415–424. [Google Scholar] [CrossRef]
- Day, M.O.; Rubidge, B.S. A brief lithostratigraphic review of the Abrahamskraal and Koonap formations of the Beaufort Group, South Africa: Towards a basin-wide stratigraphic scheme for the Middle Permian Karoo. J. Afr. Earth Sci. 2014, 100, 227–242. [Google Scholar] [CrossRef]
- Rubidge, B.S. Biostratigraphy of the Beaufort Group (Karoo Supergroup); Biostratigraphic Series 1; South African Commission for Stratigaphy: Pretoria, South Africa, 1995; pp. 1–45. [Google Scholar]
- Smith, R.M.H.; Keyser, A. Biostratigraphy of the Tapinocephalus Assemblage Zone. In Biostratigraphy of the Beaufort Group (Karoo Supergroup); South African Committee for Stratigraphy, Biostratigraphic Series; Rubidge, B.S., Ed.; Council for Geoscience: Pretoria, South Africa, 1995; Volume 1, pp. 8–12. [Google Scholar]
- Smith, R.M.H.; Keyser, A.W. Biostratigraphy of the Pristerognathus Assemblage Zone. In Biostratigraphy of the Beaufort Group (Karoo Supergroup); South African Committee for Stratigraphy, Biostratigraphic Series; Rubidge, B.S., Ed.; Council for Geoscience: Pretoria, South Africa, 1995; Volume 1, pp. 13–17. [Google Scholar]
- Smith, R.M.H. Biostratigraphy of the Cistecephalus Assemblage Zone (Beaufort Group, Karoo Supergroup), South Africa. S. Afr. J. Geol. 2020, 123, 181–190. [Google Scholar] [CrossRef]
- Mason, R. A Bio- and Litho- Stratigraphic Study of the Ecca-Beaufort contact in the southeastern Karoo Basin (Albany District, Eastern Cape Province). Master’s Thesis, University of the Witwatersrand, Johannesburg, South Africa, 2007; p. 147. [Google Scholar]
- Viglietti, P.A.; Rubidge, B.S.; Smith, R.M.H. Revised lithostratigraphy of the Upper Permian Balfour and Teekloof formations of the main Karoo Basin, South Africa. S. Afr. J. Geol. 2017, 120, 45–60. [Google Scholar] [CrossRef]
- Lucas, S.G. Timing andmagnitude of tetrapod extinctions across the Permo-Triassic boundary. J. Asian Earth Sci. 2009, 36, 491–502. [Google Scholar] [CrossRef]
- Day, M.O.; Güven, S.; Abdala, F.; Jirah, S.; Rubidge, B.; Almond, J. Youngest dinocephalian fossils extend the Tapinocephalus Zone, Karoo Basin, South Africa. S. Afr. J. Sci. 2015, 111, e2014-0309. [Google Scholar] [CrossRef]
- Day, M.O.; Smith, R.M.H. Biostratigraphy of the Endothiodon Assemblage Zone (Beaufort Group, Karoo Supergroup), South Africa. S. Afr. J. Geol. 2020, 123, 165–180. [Google Scholar] [CrossRef]
- Lanci, L.; Galeotti, S.; Ratcliffe, K.; Tohver, E.; Wilson, A.; Flint, S. Astronomically forced cycles in Middle Permian fluvial sediments from Karoo Basin (South Africa). Palaeogeogr. Palaeoclimatol, Palaeoecol. 2022, 596, 110973. [Google Scholar] [CrossRef]
- Turner, B.R. Sedimentary patterns of uranium mineralization the Beaufort Group of the southern Karoo (Gondwana) basin, South Africa. Can. Soc. Petrol. Geologists 1978, 5, 831–848. [Google Scholar]
- Pretorius, L.E. Aspects of Uranium Mineralization in the Beaufort West Karoo. Master’s Thesis, Stellenbosch University, Stellenbosch, South Africa, 1977; p. 116. [Google Scholar]
- Stear, W.M. Sedimentary structures related to fluctuating hydrodynamic conditions in flood plain deposits of the Beaufort Group near Beaufort West, Cape. Trans. Geol. Soc. S. Afr. 1978, 81, 393–399. [Google Scholar]
- Stear, W.M. The Sedimentary Environment of the Beaufort Group Uranium Province in the Vicinity of Beaufort West, South Africa. Ph.D. Thesis, University of Port Elizabeth, Port Elizabeth, South Africa, 1980; p. 188. [Google Scholar]
- Bordy, E.M.; Smith, R.M.H.; Choiniere, J.N.; Rubidge, B.S. Selected Karoo Geoheritage Sites of Paleontological Significance in South Africa and Lesotho; Geological Society of London: London, UK, 2024. [Google Scholar] [CrossRef]
- Day, M.O.; Rubidge, B.S. Biesiespoort revisited: A case study on the relationship between tetrapod assemblage zones and Beaufort lithostratigraphy south of Victoria West. Palaeont. Afr. 2019, 53, 51–65. [Google Scholar]
- Marchetti, L.; Logghe, A.; Mujal, E.; Barrier, P.; Montenat, C.; Nel, A.; Steyer, J.S. Vertebrate tracks from the Permian of Gonfaron (Provence, Southern France) and their implications for the late Capitanian terrestrial extinction event. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2022, 599, 111043. [Google Scholar] [CrossRef]
- Kammerer, C.F.; Rubidge, B.S. The earliest gorgonopsians from the Karoo Basin of South Africa. J. Afr. Earth Sci. 2022, 194, 104631. [Google Scholar] [CrossRef]
- Day, M.O.; Rubidge, B.S. The Late Capitanian mass extinction of terrestrial vertebrates in the Karoo Basin of South Africa. Frontiers Earth Sci. 2021, 9, 631198. [Google Scholar] [CrossRef]
- Smith, R.M.; Angielczyk, K.D.; Benoit, J.; Fernandez, V. Neonate aggregation in the Permian dicynodont Diictodon (Therapsida, Anomodontia): Evidence for a reproductive function for burrows? Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 569, 110311. [Google Scholar] [CrossRef]
- Klein, G.D. A sedimentary model for determining paleotidal range. Bull. Geol. Soc. Am. 1971, 82, 2585–2592. [Google Scholar] [CrossRef]
- Olsen, H.; Due, P.H.; Clemmensen, L.B. Morphology and genesis of asymmetric adhesion warts--a new adhesion surface structure. Sed. Geol. 1989, 61, 277–285. [Google Scholar] [CrossRef]
- Ramsay, P.J.; Cooper, J.A.G.; Wright, C.I.; Mason, T.R. The occurrence and formation of ladderback ripples in subtidal, shallow-marine sands, Zululand, South Africa. Marine Geol. 1989, 86, 229–235. [Google Scholar] [CrossRef]
- Reineck, H.-E.; Singh, I.B. Depositional Sedimentary Environments; Springer: Berlin/Heidelberg, Germany, 1975; p. 439. [Google Scholar]
- Tanner, W.F. An occurrence of Flat-Topped Ripple Marks. J. Sedim. Petrol. 1958, 28, 95–96. [Google Scholar] [CrossRef]
- Higgs, R. Fish trails in the Upper Carboniferous of southwest England. Palaeontology 1988, 31, 255–272. [Google Scholar]
- Gibert, J.M.; Buatois, L.A.; Fregenal-Martinez, M.A.; Mángano, M.G.; Ortega, F.; Poyato-Ariza, F.J.; Wenz, S. The fish trace fossil Undichna from the Cretaceous of Spain. Palaeontology 1999, 42, 409–427. [Google Scholar] [CrossRef]
- Trewin, N.H. The ichnogenus Undichna, with examples from the Permian of the Falkland Islands. Palaeontology 2000, 43, 979–997. [Google Scholar] [CrossRef]
- Ronchi, A.; Santi, G.; Marchetti, L.; Bernardi, M.; Gianolla, P. First report of swimming trace fossils of fish from the Upper Permian and Lower Triassic of the Dolomites (Italy). Ann. Soc. Geol. Poloniae 2018, 88, 111–125. [Google Scholar] [CrossRef]
- Minter, N.J.; Krainer, K.; Lucas, S.G.; Braddy, S.J.; Hunt, A.P. Palaeoecology of an Early Permian playa lake trace fossil assemblage from Castle Peak, Texas, USA. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2007, 246, 390–423. [Google Scholar] [CrossRef]
- Groenewald, D.P.; Krüger, A.; Day, M.O.; Penn- Clarke, C.R.; Hancox, P.J.; Rubidge, B.S. Unique trackway on Permian Karoo shoreline provides evidence of temnospondyl locomotory behaviour. PLoS ONE 2023, 18, e0282354. [Google Scholar] [CrossRef]
- Prevec, R.; Nel, A.; Day, M.O.; Muir, R.A.; Matiwane, A.; Kirkaldy, A.P.; Barber-James, H.M. South African Lagerstätte reveals middle Permian Gondwanan lakeshore ecosystem in exquisite detail. Commun. Biol. 2022, 5, 1154. [Google Scholar] [CrossRef]
- Minter, N.J.; Braddy, S.J. Ichnology of an early Permian intertidal flat: The Robledo Mountains Formation of southern New Mexico, US. In Special Papers in Paleontology 82; The Palaeontological Association: London, UK, 2009. [Google Scholar]
- Garrouste, R.; Nel, A.; Gand, G. New fossil arthropods (Notostraca and Insecta: Syntonopterida) in the continental Middle Permian of Provence (Bas-Argens Basin, France). Comptes Rendus Palevol 2009, 8, 49–57. [Google Scholar] [CrossRef]
- Moreau, J.-D.; Michelin, A.; Fara, E.; Gand, G.; Galtier, J.; Puech, G.; Fouché, S. Ichnofossils and body fossils from the Permian of the Sorgue Valley (Saint-Affrique Basin, southern France): Palaeoenvironmental implications. Hist. Biol. 2022. [Google Scholar] [CrossRef]
- Stear, W.M. Comparison of the bedform distribution and dynamics of modern and ancient sandy ephemeral flood deposits in the southwestern Karoo region, South Africa. Sed. Geol. 1985, 45, 209–230. [Google Scholar] [CrossRef]
- Rader, R.B.; Voelz, N.J.; Ward, J.V. Post-flood recovery of a macroinvertebrate community in a regulated river: Resilience of an anthropogenically altered ecosystem. Restor. Ecol. 2008, 16, 24–33. [Google Scholar] [CrossRef]
- Davies, N.S.; Shillito, A.P. True substrates: The exceptional resolution and unexceptional preservation of deep time snapshots on bedding surfaces. Sedimentology 2021, 68, 3307–3356. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ronchi, A.; Marchetti, L.; Klein, H.; Groenewald, G.H. A Middle Permian Oasis for Vertebrate and Invertebrate Life in a High-Energy Fluvial Palaeoecosystem of Southern Gondwana (Karoo, Republic of South Africa). Geosciences 2023, 13, 325. https://doi.org/10.3390/geosciences13110325
Ronchi A, Marchetti L, Klein H, Groenewald GH. A Middle Permian Oasis for Vertebrate and Invertebrate Life in a High-Energy Fluvial Palaeoecosystem of Southern Gondwana (Karoo, Republic of South Africa). Geosciences. 2023; 13(11):325. https://doi.org/10.3390/geosciences13110325
Chicago/Turabian StyleRonchi, Ausonio, Lorenzo Marchetti, Hendrik Klein, and Gideon Hendrik Groenewald. 2023. "A Middle Permian Oasis for Vertebrate and Invertebrate Life in a High-Energy Fluvial Palaeoecosystem of Southern Gondwana (Karoo, Republic of South Africa)" Geosciences 13, no. 11: 325. https://doi.org/10.3390/geosciences13110325
APA StyleRonchi, A., Marchetti, L., Klein, H., & Groenewald, G. H. (2023). A Middle Permian Oasis for Vertebrate and Invertebrate Life in a High-Energy Fluvial Palaeoecosystem of Southern Gondwana (Karoo, Republic of South Africa). Geosciences, 13(11), 325. https://doi.org/10.3390/geosciences13110325