
Carbon Isotopic Signature and Organic Matter Composition of Cenomanian High-Latitude Paleosols of Southern Patagonia

 , , ,
, , ,
Abstract
:1. Introduction
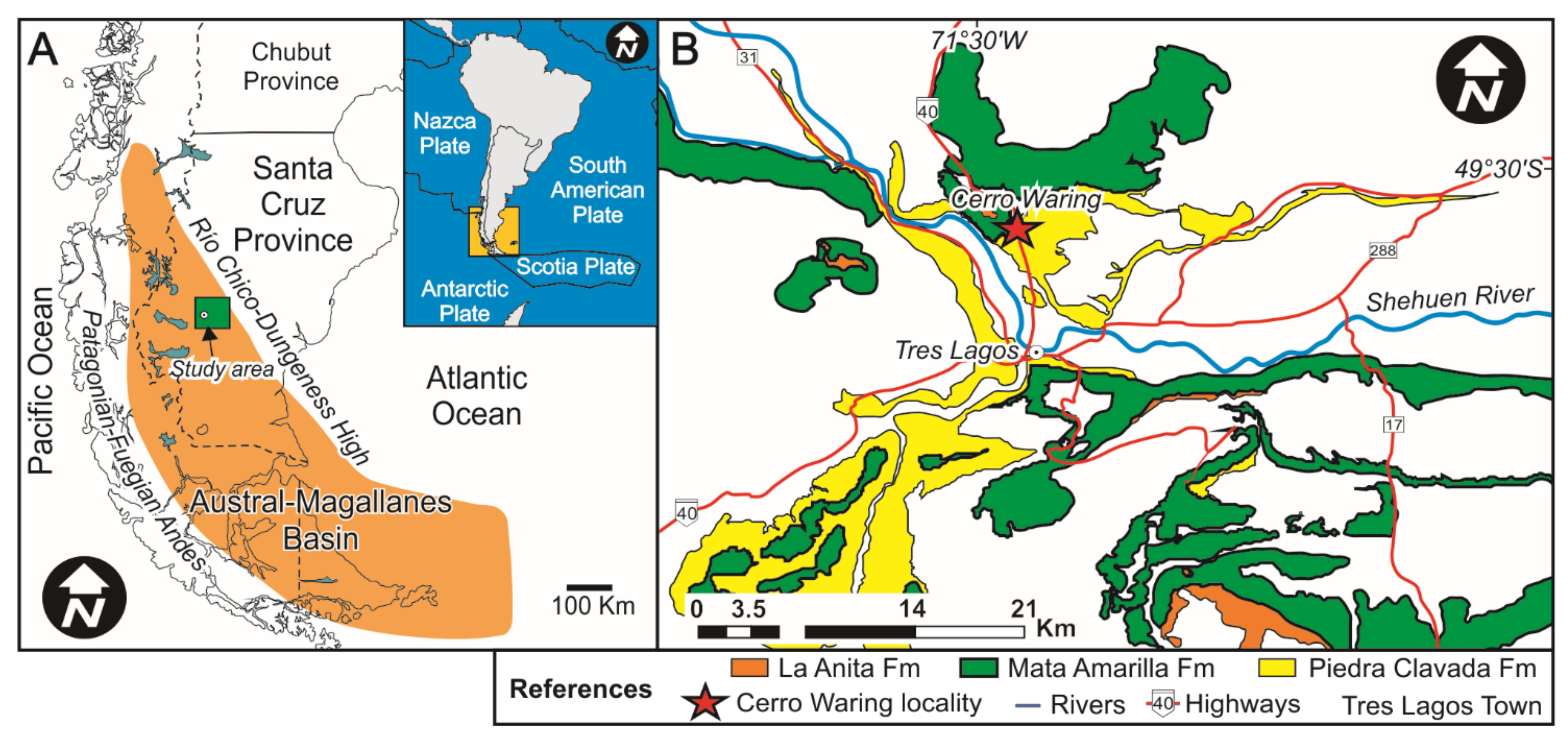
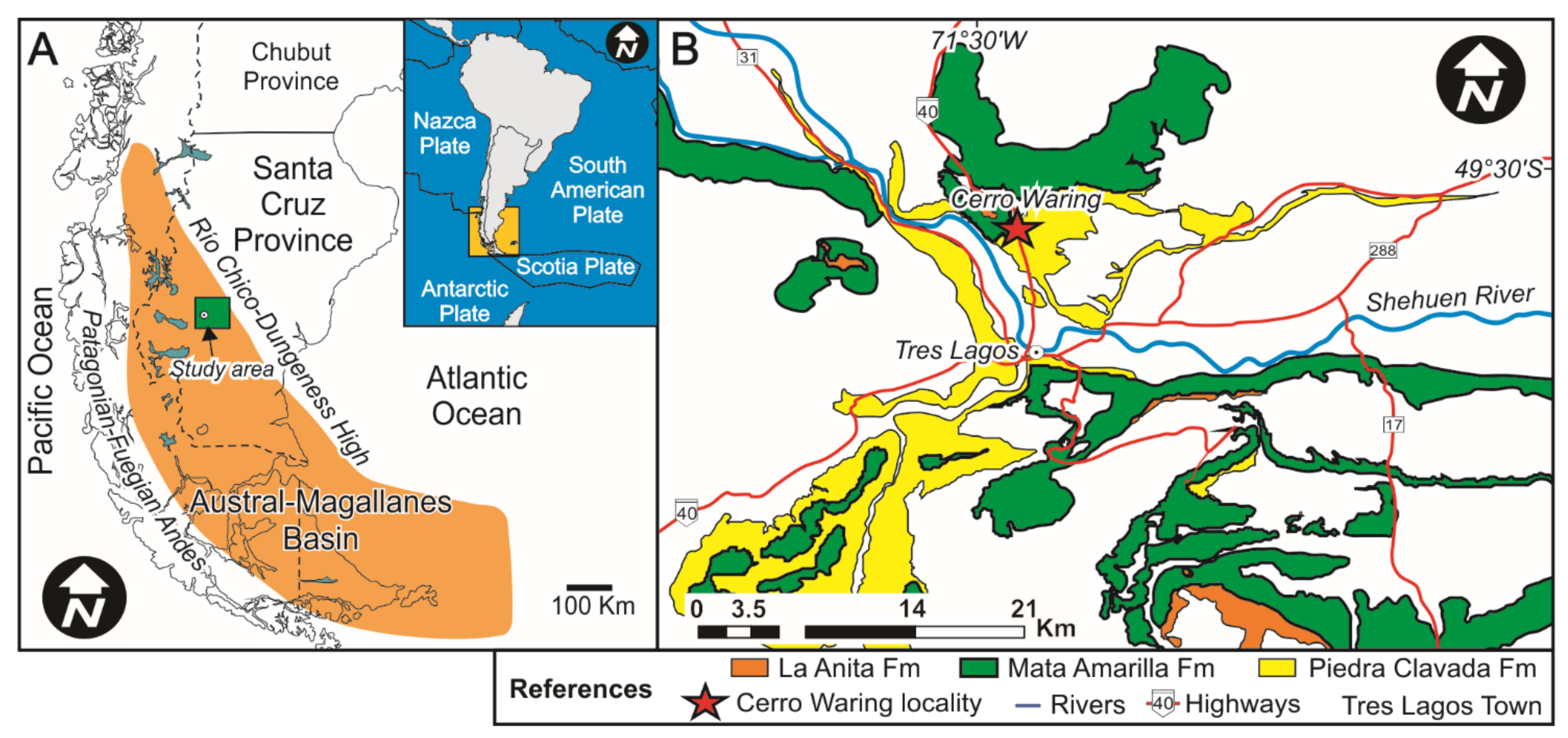
2. Sedimentological-Paleopedological Context
3. Materials and Methods
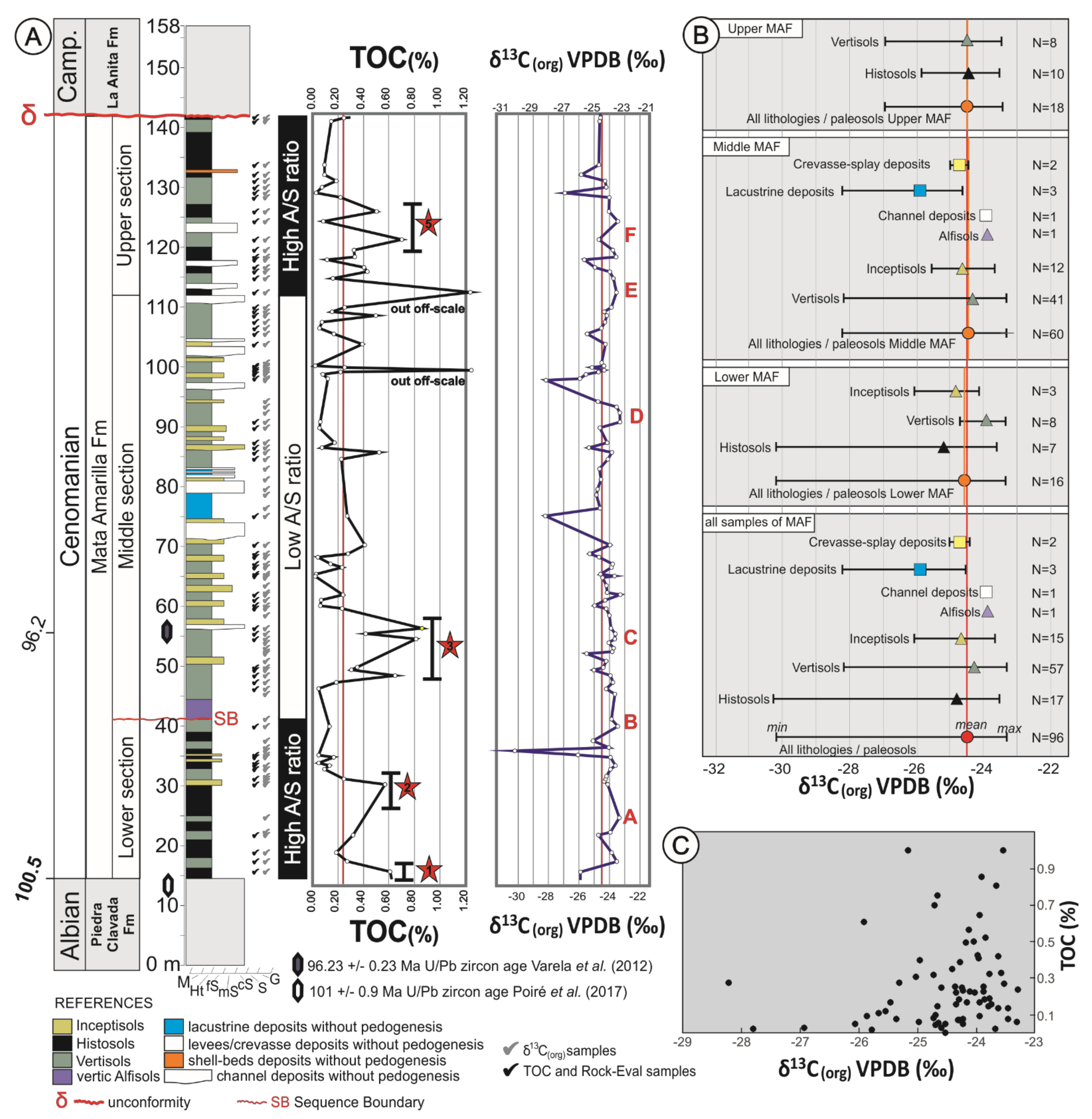
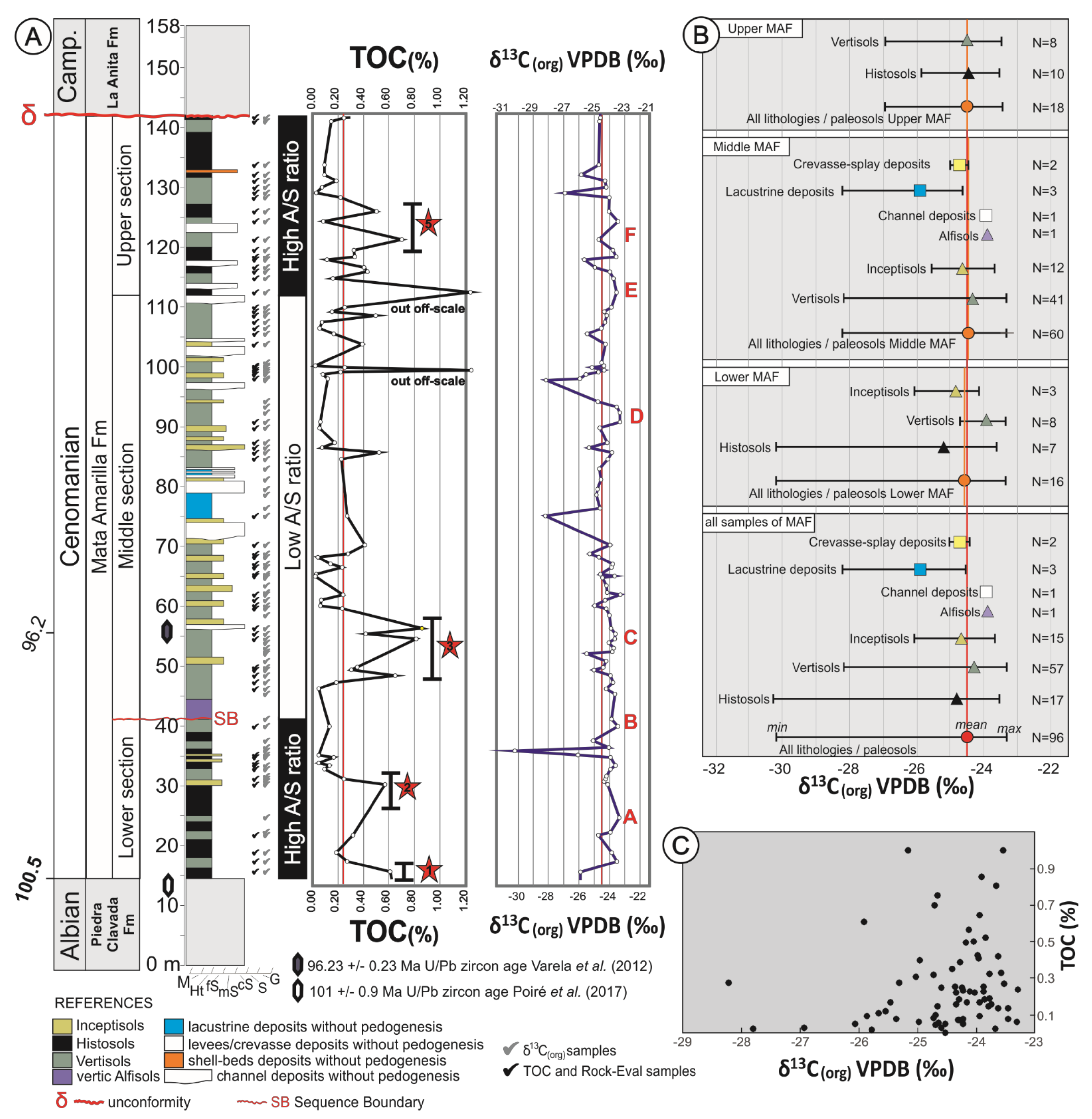
4. Results
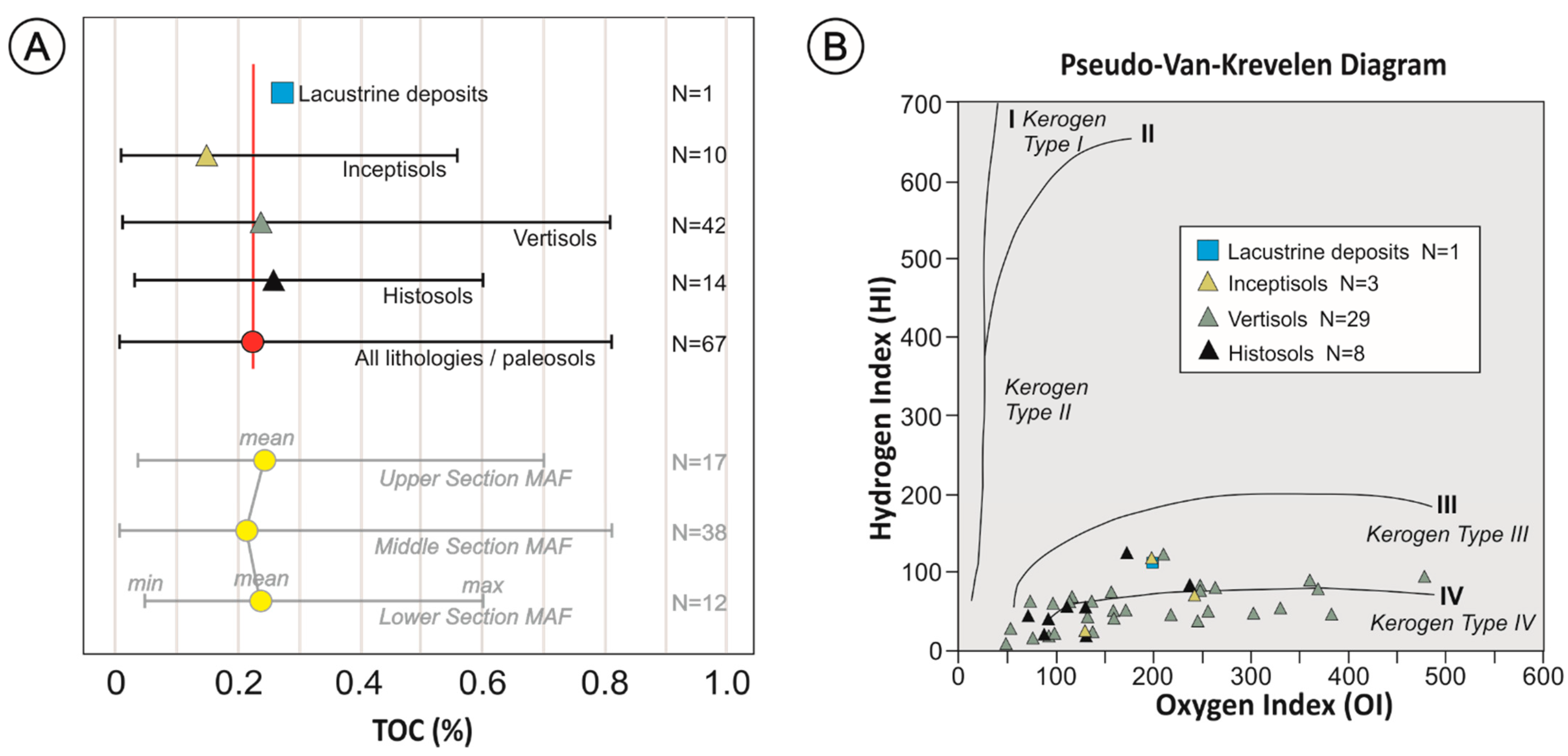
5. TOC and Rock Eval Pyrolysis
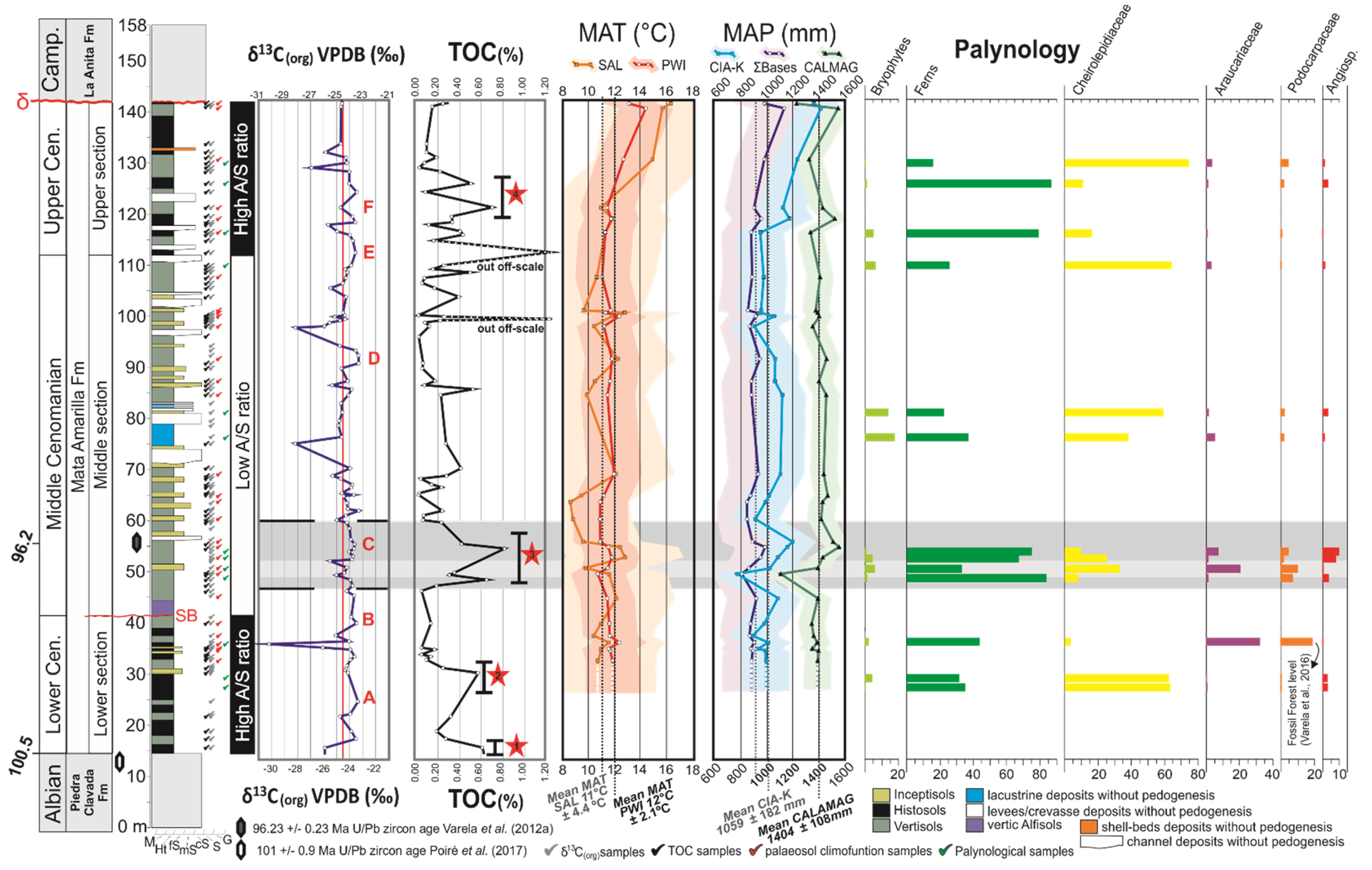
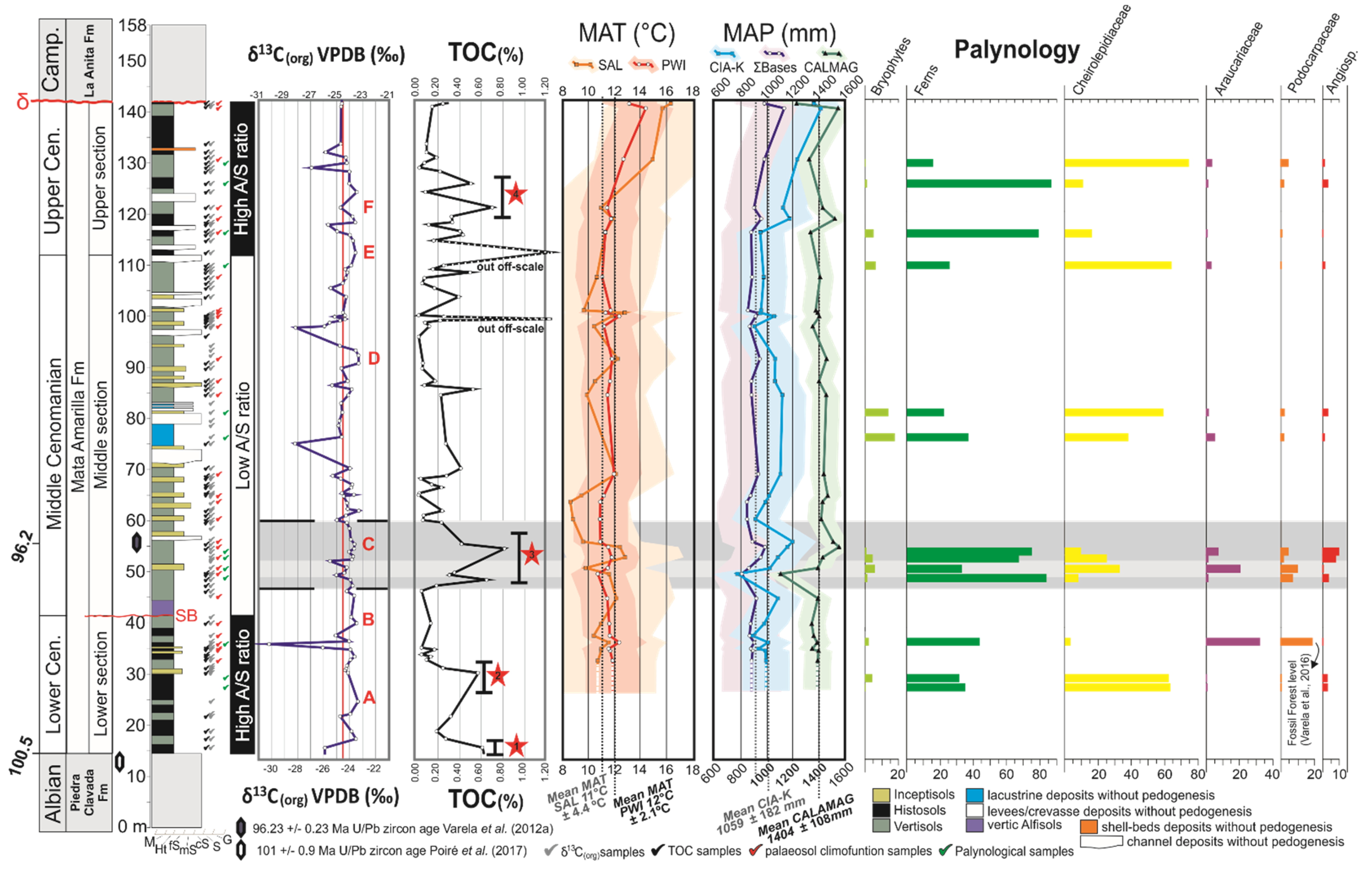
5.1. Carbon Isotope Analysis
5.2. Palynology
6. Discussion
6.1. Composition of the Paleosol-Derived (and Sedimentary) Organic Matter
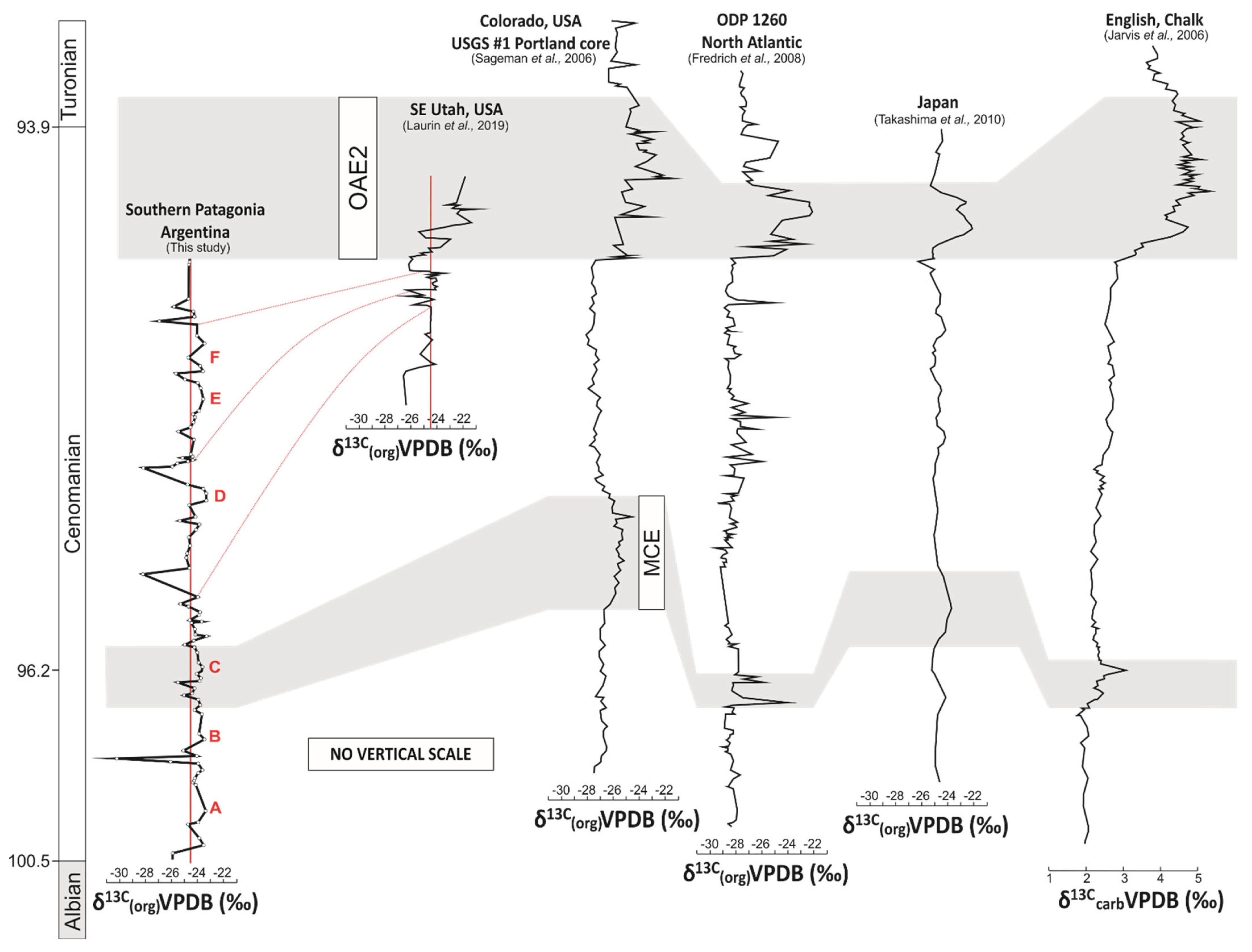
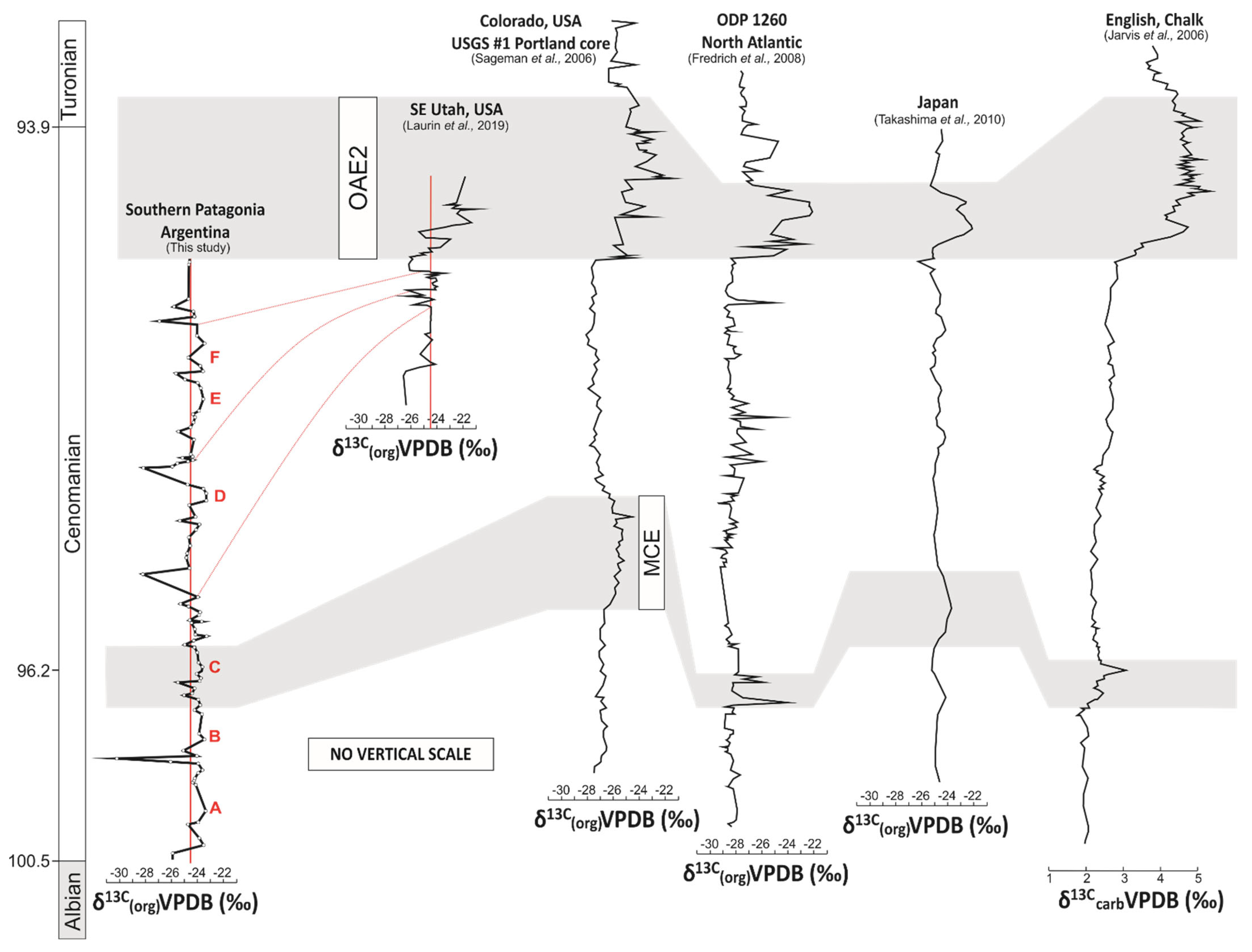
6.2. Comparison with Existing Marine and Terrestrial Carbon Isotope Records
6.3. The Mata Amarilla Formation as Environmental Archive
7. Conclusions
- This study provided a new detailed δ13Corg Cenomanian record for a continental sedimentary succession from southern South America.
- The carbon isotope record from the MAF was largely independent of paleosol type and depositional environment, demonstrating that the δ13C signature of soil-derived OC was not strongly biased due to taphonomic/pedogenic processes.
- Comparisons of the δ13Corg curve of southern Patagonia with high-resolution trends from elsewhere in the world revealed the absence of the characteristic positive CIE expected to mark the Cenomanian-Turonian boundary. The absence of the Cenomanian-Turonian CIE was interpreted to reflect erosional processes and/or non-deposition as indicated by an unconformity surface at the very base of the overlying Campanian deposits of the La Anita Formation.
- The comparison of the δ13Corg record, TOC values, and palynological data with paleosol-derived MAP provided a better understanding of the mid-Cretaceous climate/biota interaction in the Southern Hemisphere. During warmer and moister periods, fern prairies developed at the expense of the background conifer-dominated forest. Angiosperms formed only a minor component of the Cenomanian Southern Hemisphere high-latitude vegetation.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Clapham, M.E.; Renne, P.R. Flood basalts and mass extinctions. Annu. Rev. Earth Planet. Sci. 2019, 47, 275–303. [Google Scholar] [CrossRef]
- Barron, E.J. A warm equable Cretaceous: The nature of the problem. Earth-Sci. Rev. 1983, 19, 305–338. [Google Scholar] [CrossRef]
- Arthur, M.A.; Dean, W.E.; Schlanger, S.O. Variations in the global carbon cycle during the Cretaceous related to climate, volcanism and changes in atmospheric CO2. In The Carbon Cycle and Atmospheric CO2: Natural Variations Archean to Present; AGU Geophysical Monograph; Sundquist, E.T., Broecker, W.S., Eds.; American Geophysical Union: Washington, DC, USA, 1985; Volume 32, pp. 504–529. [Google Scholar]
- Poulsen, C.J.; Pollard, D.; White, T.S. General circulation model simulation of the δ18O content of continental precipitation in the middle Cretaceous: A model-proxy comparison. Geology 2007, 35, 199–202. [Google Scholar] [CrossRef]
- Bodin, S.; Meissner, P.; Janssen, N.M.M.; Steuber, T.; Mutterlose, J. Large igneous provinces and organic carbon burial: Controls on global temperature and continental weathering during the Early Cretaceous. Glob. Planet. Change 2015, 133, 238–253. [Google Scholar] [CrossRef]
- Jenkyns, H.C.; Dickson, A.J.; Ruhl, M.; Van Den Boorn, S.H.J.M. Basalt-seawater interaction, the Plenus cold event, enhanced weathering and geochemical change: Deconstructing oceanic anoxic event 2 (Cenomanian-Turonian, Late Cretaceous). Sedimentology 2017, 64, 16–43. [Google Scholar] [CrossRef]
- O’Brien, C.L.; Robinson, S.A.; Pancost, R.D.; Sinninghe Damsté, J.S.; Schouten, S.; Lunt, D.J.; Alsenz, H.; Bornemann, A.; Bottini, C.; Brassell, S.C.; et al. Cretaceous sea-surface temperature evolution: Constraints from TEX86 and planktonic foraminiferal oxygen isotopes. Earth-Sci. Rev. 2017, 172, 224–247. [Google Scholar] [CrossRef]
- Varela, A.N.; Raigemborn, M.S.; Richiano, S.; White, T.; Poiré, D.G.; Lizzoli, S. Late Cretaceous paleosols as paleoclimate proxies of high-latitude Southern Hemisphere: Mata Amarilla Formation, Patagonia, Argentina. Sediment. Geol. 2018, 363, 83–95. [Google Scholar] [CrossRef]
- O’Connor, L.K.; Robinson, S.A.; Naafs, B.D.A.; Jenkyns, H.C.; Henson, S.; Clarke, M.; Pancost, R.D. Late Cretaceous temperature evolution of the southern high latitudes: A TEX86 perspective. Paleoceanogr. Paleoclimatol. 2019, 34, 436–454. [Google Scholar] [CrossRef] [Green Version]
- Klages, J.P.; Salzmann, U.; Bickert, T.; Hillenbrand, C.D.; Gohl, K.; Kuhn, G.; Bohaty, S.M.; Titschack, J.; Müller, J.; Frederichs, T.; et al. Temperate rainforests near the South Pole during peak Cretaceous warmth. Nature 2020, 580, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Barron, E.J.; Hay, W.W.; Thompson, S. The hydrologic cycle. A major variable during Earth history. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1989, 75, 157–174. [Google Scholar] [CrossRef]
- White, T.; González, L.; Ludvigson, G.; Poulsen, C. Middle Cretaceous greenhouse hydrologic cycle of North America. Geology 2001, 29, 363–366. [Google Scholar] [CrossRef]
- White, T.; Witzke, B.; Ludvigson, G.; Brenner, R. Distinguishing base-level change and climate signals in a Cretaceous alluvial sequence. Geology 2005, 33, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Van Helmond, N.A.G.M.; Sluijs, A.; Reichart, G.-J.; Sinninghe Damsté, J.S.; Slomp, C.P.; Brinkhuis, H. A perturbed hydrological cycle during Oceanic Anoxic Event 2. Geology 2014, 42, 123–126. [Google Scholar] [CrossRef]
- Ludvigson, G.A.; Joeckel, R.M.; Murphy, L.R.; Stockli, D.F.; González, L.A.; Suarez, C.A.; Kirkland, J.I.; Al-Suwaidi, A. The emerging terrestrial record of Aptian-Albian global change. Cretac. Res. 2015, 56, 1–24. [Google Scholar] [CrossRef]
- Föllmi, K.B.; Weissert, H.; Bisping, M.; Funk, H. Phosphogenesis, carbon isotope stratigraphy, and carbonate-platform evolution along the Lower Cretaceous northern Tethyan margin. Geol. Soc. Am. Bull. 1994, 106, 729–746. [Google Scholar] [CrossRef]
- Turgeon, S.; Brumsack, H.-J. Anoxic vs dysoxic events reflected in sediment geochemistry during the Cenomanian–Turonian Boundary Event (Cretaceous) in the Umbria–Marche Basin of central Italy. Chem. Geol. 2006, 234, 321–339. [Google Scholar] [CrossRef]
- Schlanger, S.O.; Jenkyns, H.C. Cretaceous Oceanic Anoxic Events: Causes and Consequences. Geol. Mijnb. 1976, 55, 179–184. [Google Scholar]
- Jenkyns, H.C. Cretaceous anoxic events: From continents to oceans. J. Geol. Soc. 1980, 137, 171–188. [Google Scholar] [CrossRef]
- Jenkyns, H.C. Geochemistry of oceanic anoxic events. Geochem. Geophys. 2010, 11, 3. [Google Scholar] [CrossRef]
- Robinson, S.A.; Heimhofer, U.; Hesselbo, S.P.; Petrizzo, M.R. Mesozoic climates and oceans – a tribute to Hugh Jenkyns and Helmut Weissert. Sedimentology 2017, 64, 1–15. [Google Scholar] [CrossRef]
- Cramer, B.D.; Jarvis, I. Carbon Isotope Stratigraphy. In Geologic Time Scale; Gradstein, F.M., Ogg, J.G., Schmitz, M.D., Ogg, G.M., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 1, pp. 309–343. [Google Scholar]
- Meyers, S.R.; Siewert, S.E.; Singer, B.S.; Sageman, B.B.; Condon, D.J.; Obradovich, J.D.; Jicha, B.R.; Sawyer, D.A. Intercalibration of radioisotopic and astrochronologic time scales for the Cenomanian-Turonian boundary interval, Western Interior Basin, USA. Geology 2012, 40, 7–10. [Google Scholar] [CrossRef] [Green Version]
- Eldrett, J.S.; Ma, C.; Bergmann, S.C.; Lutz, B.; Gregory, F.J.; Dodsworth, P.; Phipps, M.; Hardas, P.; Minisini, D.; Ozkan, A.; et al. An astronomically calibrated stratigraphy of the Cenomanian, Turonian and earliest Coniacian from the cretaceous Western Interior Seaway, USA: Implications for global cronostratigraphy. Cretac. Res. 2015, 56, 316–344. [Google Scholar] [CrossRef]
- Voigt, S.; Gale, A.S.; Flögel, S. Midlatitude shelf seas in the Cenomanian–Turonian greenhouse world: Temperature evolution and North Atlantic circulation. Paleoceanography 2004, 19, PA4020. [Google Scholar] [CrossRef] [Green Version]
- Forster, A.; Schouten, S.; Baas, M.; Sinninghe Damste, J.S. Mid-Cretaceous (Albian-Santonian) sea surface temperature record of the tropical Atlantic Ocean. Geology 2007, 35, 919–922. [Google Scholar] [CrossRef]
- Hasegawa, T. Cenomanian-Turonian carbon isotope events recorded in terrestrial organic matter from northern Japan. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 130, 251–273. [Google Scholar] [CrossRef]
- Takashima, R.; Nishi, H.; Yamanaka, T.; Hayashi, K.; Waseda, A.; Obuse, A.; Tomosugi, T.; Deguchi, N.; Mochizuki, S. Prevailing oxic environments in the Pacific Ocean during the mid-Cretaceous Oceanic Anoxic Event 2. Nat. Commun. 2011, 2, 234. [Google Scholar] [CrossRef] [PubMed]
- Barclay, R.S.; McElwain, J.C.; Sageman, B.B. Carbon sequestration activated by a volcanic CO2 pulse during Ocean Anoxic Event 2. Nat. Geosci. 2010, 3, 205–208. [Google Scholar] [CrossRef]
- Laurin, J.; Barclay, R.S.; Sageman, B.B.; Dawson, R.; Pagani, M.; Schmitz, M.; Eaton, J.; McElwain, J.C.; McInerney, F.A. Terrestrial and marginal marine record of the mid-Cretaceous Oceanic Anoxic Event 2 (OAE 2): High-resolution framework, carbon isotopes, CO2 and sea-level change. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 524, 118–136. [Google Scholar] [CrossRef]
- Jenkyns, H.C.; Gale, A.S.; Corfield, R.M. Carbon- and oxygen-isotope stratigraphy of the English Chalk and Italian Scaglia and its palaeoclimatic significance. Geol. Mag. 1994, 131, 1–34. [Google Scholar] [CrossRef] [Green Version]
- Sageman, B.B.; Meyers, S.R.; Arthur, M.A. Orbital time scale and new C-isotope record for Cenomanian–Turonian boundary stratotype. Geology 2006, 34, 125–128. [Google Scholar] [CrossRef]
- Friedrich, O.; Erbacher, J.; Moriya, K.; Wilson, P.A.; Kuhnert, H. Warm saline intermediate waters in the cretaceous tropical Atlantic Ocean. Nat. Geosci. 2008, 1, 453–457. [Google Scholar] [CrossRef]
- Andrieu, S.; Brigaud, B.; Rabourg, T.; Noret, A. The Mid-Cenomanian Event in shallow marine environments: Influence on carbonate producers and depositional sequences (northern Aquitaine Basin, France). Cretac. Res. 2015, 56, 587–607. [Google Scholar] [CrossRef]
- Joo, Y.J.; Sageman, B.B.; Hurtgen, M.T. Data-model comparison reveals key environmental changes leading to Cenomanian-Turonian Oceanic Anoxic Event 2. Earth-Sci. Rev. 2020, 203, 103–123. [Google Scholar] [CrossRef]
- Uramoto, G.-I.; Fujita, T.; Takahashi, A.; Hirano, H. Cenomanian (Upper Cretaceous) carbon isotope stratigraphy of terrestrial organic matter for the Yezo Group, Hokkaido, Japan. Isl. Arc 2007, 16, 465–478. [Google Scholar] [CrossRef]
- Takashima, R.; Nishi, H.; Yamanaka, T.; Hayashi, K.; Waseda, A.; Obuse, A.; Tomosugi, T.; Deguchi, N.; Mochizuki, S. High-resolution terrestrial carbon isotope and planktic foraminiferal records of the Upper Cenomanian to the lower Campanian in the Northwest Pacific. Earth Planet. Sci. Lett. 2010, 289, 570–582. [Google Scholar] [CrossRef]
- Batenburg, S.J.; De Vleeschouwer, D.; Sprovieri, M.; Hilgen, F.J.; Gale, A.S.; Singer, B.S.; Koeberl, C.; Coccioni, R.; Claeys, P.; Montanari, A. Orbital control on the timing of oceani anoxia in the Late Cretaceous, Discuss. Clim. Past. 2016, 12, 1995–2009. [Google Scholar] [CrossRef] [Green Version]
- Scaife, J.D.; Ruhl, M.; Dickson, A.J.; Mather, T.A.; Jenkyns, H.C.; Percival, L.M.E.; Hesselbo, S.P.; Cartwright, J.; Eldrett, J.S.; Bergman, S.C.; et al. Sedimentary mercury enrichments as a marker for submarine large Igneous Province volcanism? Evidence from Mid-Cenomanian event and Oceanic Anoxic Event 2 (late cretaceous). Geochem. Geophys. 2017, 18, 4253–4275. [Google Scholar] [CrossRef] [Green Version]
- Biddle, K.; Uliana, M.; Mitchum, R., Jr.; Fitzgerald, M.; Wright, R. The stratigraphic and structural evolution of central and eastern Magallanes Basin, Southern America. In Foreland Basins; Allen, P., Homewoods, P., Eds.; Blackwell Scientific, Special Publication–International Association of Sedimentologists: New Jersey, NJ, USA, 1986; Volume 8, pp. 41–61. [Google Scholar]
- Cuitiño, J.I.; Varela, A.N.; Ghiglione, M.C.; Richiano, S.; Poiré, D.G. The Austral-Magallanes Basin (Southern Patagonia): A synthesis of its stratigraphy and evolution. Lat. Am. J. Sedimentol. Basin Anal. 2019, 26, 155–166. [Google Scholar]
- Arbe, H.A. Análisis estratigráfico del Cretácico de la Cuenca Austral. In Proceedings of the Geología y Recursos Naturales de Santa Cruz. Relatorio del Decimoquinto Congreso Geológico Argentino, El Calafate, Argentina, 23–26 April 2002; pp. 103–128. [Google Scholar]
- Varela, A.N.; Poiré, D.G.; Martin, T.; Gerdes, A.; Goin, F.J.; Gelfo, J.N.; Hoffmann, S. U-Pb zircon constraints on the age of the Cretaceous Mata Amarilla formation, southern Patagonia, Argentina: Its relationship with the evolution of the Austral basin. Andean Geol. 2012, 39, 359–379. [Google Scholar] [CrossRef] [Green Version]
- Varela, A.N.; Richiano, S.; D’Elia, L.; Moyano Paz, D.; Tettamanti, C.; Poiré, D.G. Sedimentology and stratigraphy of the Puesto El Moro Formation, Patagonia, Argentina: Implications for Upper Cretaceous paleogeographic reconstruction and compartmentalization of the Austral-Magallanes Basin. J.Soutg Am. Earth Sci. 2019, 92, 466–480. [Google Scholar] [CrossRef]
- Van Hinsbergen, D.J.J.; De Groot, L.V.; van Schaik, S.J.; Spakman, W.; Bijl, P.K.; Sluijs, A.; Langereis, C.G.; Brinkhuis, H. A paleolatitude calculator for palaeoclimate studies. PLoS ONE 2015, 10, e0126946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varela, A.N.; Richiano, S.; Poiré, D.G. Tsunami vs storm origin for shell bed deposits in a lagoon environment: An example from the Upper Cretaceous of Southern Patagonia, Argentina. Lat. Am. J. Sedimentol. Basin Anal. 2011, 18, 63–85. [Google Scholar]
- Varela, A.N. Tectonic control of accommodation space and sediment supply within the Mata Amarilla Formation (lower Upper Cretaceous) Patagonia, Argentina. Sedimentology 2015, 62, 867–896. [Google Scholar] [CrossRef]
- Archangelsky, S. Biogeographic implications of Albian Mohria-like spores (family Anemiaceae) in SW Gondwana (Patagonia). Rev. Palaeobot. Palynol. 2009, 157, 301–308. [Google Scholar] [CrossRef]
- Poiré, D.G.; Iglesias, A.; Varela, A.N.; Richiano, S.; Mejía Ibañez, M.; Stroemberg, C. Edades U-Pb en Zircones de Tobas de la Fm. Piedra Clavada, Pcia. de Santa Cruz, Argentina: Un Marcador Albiano Tardio Para la Evolución Tectónica y Biológica de la Cuenca Austral. In Proceedings of the XX Congreso Geológico Argentino, Tucumán, Argentina, 7–11 August 2017. [Google Scholar]
- Moyano Paz, D.; Tettamanti, C.; Varela, A.N.; Cereceda, A.; Poiré, D.G. Depositional processes and stratigraphic evolution of the Campanian deltaic system of La Anita Formation, Austral-Magallanes Basin, Patagonia, Argentina. Lat. Am. J. Sedimentol. Basin Anal. 2018, 25, 69–92. [Google Scholar]
- Moyano Paz, D.; Richiano, S.; Varela, A.N.; Gómez Dacál, A.R.; Poiré, D.G. Ichnological signatures from wave- and fluvial-dominated deltas: The La Anita Formation, Upper Cretaceous, Austral-Magallanes Basin, Patagonia. Mar. Pet. Geol. 2020, 113, 104168. [Google Scholar] [CrossRef]
- Sickmann, Z.T.; Schwartz, T.M.; Graham, S.A. Refining stratigraphy and tectonic history using detrital zircón máximum depositional age: An example from the Cerro Fortaleza Formation, Austral Basin, southern Patagonia. Basin Res. 2018, 30, 708–729. [Google Scholar] [CrossRef]
- Varela, A.N.; Veiga, G.D.; Poiré, D.G. Sequence stratigraphic analysis of Cenomanian greenhouse palaeosols: A case study from southern Patagonia, Argentina. Sediment. Geol. 2012, 271–272, 67–82. [Google Scholar] [CrossRef]
- Varela, A.N.; Iglesias, A.; Poiré, D.G.; Zamuner, A.B.; Richiano, S.; Brea, M. Petrified forests in the Austral Basin marks a Cenomanian forced regression heterogeneous surface. Geobiology 2016, 14, 293–313. [Google Scholar] [CrossRef] [PubMed]
- Iglesias, A.; Zamuner, A.B.; Poiré, D.G.; Larriestra, F. Diversity, taphonomy, and palaeoecology of an angiosperm flora from the Cretaceous (Cenomanian-Coniacian) in Southern Patagonia, Argentina. Palaeontology 2007, 50, 445–466. [Google Scholar] [CrossRef]
- Santamarina, P.E.; Barreda, V.D.; Iglesias, A.; Varela, A.N. Salvinialean megaspores in the Late Cretaceous of southern Patagonia, Argentina. Acta Palaeontol. Pol. 2018, 63, 607–616. [Google Scholar] [CrossRef]
- Santamarina, P.E.; Barreda, V.D.; Iglesias, A.; Varela, A.N. Palynology from the Cenomanian Mata Amarilla Formation, southern Patagonia, Argentina. Cretac. Res. 2020, 109, 104354. [Google Scholar] [CrossRef]
- Lafargue, E.; Marquis, F.; Pillot, D. Rock-Eval 6 applications in hydrocarbon exploration, production, and soil contamination studies. Oil Gas Sci. Technol. 1998, 53, 421–437. [Google Scholar] [CrossRef] [Green Version]
- Behar, F.; Beaumont, V.; Penteado, H.L.D.B. Rock-Eval 6 Technology: Performances and Developments. Oil Gas Sci. Technol. 2001, 56, 111–134. [Google Scholar] [CrossRef]
- Noetinger, S.; Pujana, R.R.; Burrieza, A.; Burrieza, H.P. Use of UV-curable acrylates gels as mounting media for palynological samples. Rev. Del Mus. Argent. De Cienc. Nat. Nueva Ser. 2017, 19, 19–23. [Google Scholar] [CrossRef]
- Raine, J.I.; Mildenhall, D.C.; Kennedy, E.M. New Zealand Fossil Spores and Pollen: An Illustrated Catalogue, 4th ed.; GNS Science Miscellaneous Series: Lower Hutt, New Zealand, 2011. [Google Scholar]
- McCarthy, K.; Rojas, K.; Niemann, M.; Palmowski, D.; Peters, K.; Stankiewicz, A. Basic Petroleum Geochemistry for Source Rock Evaluation. Oilfield Rev. 2011, 23, 32–43. [Google Scholar]
- Abbassi, S.; Edwards, D.S.; George, S.C.; Volk, H.; Mahlstedt, N.; di Primio, R.; Horsfield, B. Petroleum potential and kinetic models for hydrocarbon generation from the Upper Cretaceous to Paleogene Latrobe Group coals and shales in the Gippsland Basin, Australia. Org. Geochem. 2016, 91, 54–67. [Google Scholar] [CrossRef]
- Broz, A.P. Organic matter preservation in ancient soils of Earth and Mars. Life 2020, 10, 113. [Google Scholar] [CrossRef]
- Ferguson, B.; Lukens, W.E.; Masri, B.E.; Stinchcomb, G.E. Alluvial landform and the occurrence of paleosols in a humid-subtropical climate have an effect on long-term soil organic carbon storage. Geoderma 2020, 371, 114388. [Google Scholar] [CrossRef]
- Retallack, G.J. Interflag sandstone laminae, a novel sedimentary structure, with implications for Ediacaran paleoenvironments. Sediment. Geol. 2019, 379, 60–76. [Google Scholar] [CrossRef]
- Lee, Y.W.; Lee, Y.I.; Hisada, K. Paleosols in the Cretaceous Goshoura and Mifune groups, SW Japan and their paleoclimate implications. Palaeogr. Palaeoclimatol. Palaeoecol. 2003, 199, 265–282. [Google Scholar] [CrossRef]
- Mirzaloo, M.; Ghasemi-Nejad, E. Combined use of palynology and organic geochemistry in petroleum potential evaluation and paleoenvironmental interpretation of the Kazhdumi Formation (Aptian-Cenomanian) in the southwestern Zagros Basin, Iran. Geopersia 2012, 21, 27–40. [Google Scholar]
- Masseroli, A.; Villa, S.; Mariani, G.S.; Bollati, I.M.; Pelfini, M.; Sebag, D.; Verrecchia, E.P.; Trombino, L. Reconsidering the compound effect of geomorphology, vegetation, and climate change on paleopedogenesis in sensitive environments (Northern Apennines, Italy). Catena 2021, 197, 104951. [Google Scholar] [CrossRef]
- Nordt, L.C. Stable carbon and oxygen isotopes in soils. In Earth Sciences and Archaeology; Goldberg, P., Holliday, V.T., Ferring, C.R., Eds.; Springer: Boston, MA, USA, 2001; pp. 419–448. [Google Scholar]
- Wynn, J.G. Carbon isotope fractionation during decomposition of organic matter in soils and paleosols: Implications for paleoecological interpretations of paleosols. Palaeogeogr. Palaeoclimat. Palaeoecol. 2007, 251, 437–448. [Google Scholar] [CrossRef]
- Zhu, C.; Wagner, T.; Pan, J.-M.; Pancost, R.D. Multiple sources and extensive degradation of terrestrial sedimentary organic matter across an energetic, wide continental shelf. Geochem. Geophys 2011, 12, Q08011. [Google Scholar]
- Graham, H.V.; Herrera, F.; Jaramillo, C.; Wing, S.L.; Freeman, K.H. Canopy structure in Late Cretaceous and Paleocene forests as reconstructed from carbon isotope analyses of fossil leaves. Geology 2019, 47, 977–981. [Google Scholar] [CrossRef]
- Arens, N.C.; Jahren, A.H. Carbon Isotope Excursion in Atmospheric CO2 at the Cretaceous-Tertiary Boundary: Evidence from Terrestrial Sediments. Palaios 2000, 15, 314–322. [Google Scholar] [CrossRef]
- Weissert, H.; Joachimski, M.; Sarnthein, M. Chemostratigraphy. Newsl. Stratigr. 2008, 42, 145–179. [Google Scholar] [CrossRef]
- Cojan, I. Stable carbon isotope stratigraphy of the Paleogene pedogenic series of southern France as a basis for continental-marine correlation. Geology 2000, 28, 259–262. [Google Scholar] [CrossRef]
- Gröcke, D.R.; Ludvigson, G.A.; Witzke, B.L.; Robinson, S.A.; Joeckel, R.M.; Ufnar, D.F.; Ravn, R.L. Recognizing the Albian– Cenomanian (OAE1d) sequence boundary using plant carbon isotopes Dakota Formation, Western Interior basin, USA. Geology 2006, 34, 193–196. [Google Scholar] [CrossRef]
- Maufrangeas, A.; Leleu, S.; Loisy, C.; Roperch, P.; Jolley, D.; Vinciguerra, C.; Nguyen-Thuye, O. Stratigraphy in Palaeocene continental sedimentary succession of the Northern Pyrenean Basin (Corbières, southern France) using δ13Corg isotopes. J. Geol. Soc. 2020, 177, 752–756. [Google Scholar] [CrossRef]
- Raigemborn, M.S.; Krapovickas, V.; Zucol, A.F.; Zapata, L.; Beilinson, E.; Toledo, N.; Perry, J.; Lizzoli, S.; Martegani, L.; Tineo, D.E.; et al. Paleosols and related soil biota of the early Miocene Santa Cruz Formation (Austral-Magallanes Basin, Argentina): A multidisciplinary approach to reconstructing ancient terrestrial landscapes. Lat. Am. J. Sedimentol. Basin Anal. 2018, 25, 117–148. [Google Scholar]
- Jahren, A.H.; Arens, N.C.; Sarmiento, G.; Guerrero, J.; Amundson, R. Terrestrial record of methane hydrate dissociation in the Early Cretaceous. Geology 2001, 29, 159–162. [Google Scholar] [CrossRef] [Green Version]
- Heimhofer, U.; Hochuli, P.A.; Burla, S.; Andersen, N.; Weissert, H. Terrestrial carbon-isotope records from coastal deposits (Algarve, Portugal): A tool for chemostratigraphic correlation on an intrabasinal and global scale. Terra Nova 2003, 15, 8–13. [Google Scholar] [CrossRef]
- Hesselbo, S.P.; Gröcke, D.R.; Jenkyns, H.C.; Bjerrum, C.J.; Farrimond, P.; Morgans Bell, H.S.; Green, O.R. Massive dissociation of gas hydrate during a Jurassic oceanic anoxic event. Nature 2000, 406, 392–395. [Google Scholar] [CrossRef]
- Robinson, S.A.; Hesselbo, S.P. Fossil-wood carbon-isotope stratigraphy of the non-marine Wealden Group (Lower Cretaceous, southern England). J. Geol. Soc. 2004, 161, 133–145. [Google Scholar] [CrossRef] [Green Version]
- Schneebeli-Hermann, E.; Kürschner, W.M.; Hochuli, P.A.; Weissert, H.; Bernasconi, S.M.; Roohi, G.; ur-Rehman, K.; Goudemand, N.; Bucher, H. Evidence for atmospheric carbon injection during the end-Permian extinction. Geology 2013, 41, 579–582. [Google Scholar] [CrossRef]
- Hasegawa, T.; Pratt, L.M.; Maeda, H.; Shigeta, Y.; Okamoto, T.; Kase, T.; Uemura, K. Upper Cretaceous stable carbon isotope stratigraphy of terrestrial organic matter from Sakhalin, Russian Far East. A proxy for the isotopic composition of paleoatmospheric CO2. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 189, 97–115. [Google Scholar] [CrossRef]
- Suarez, M.B.; Ludvigson, G.A.; González, L.A.; Al-Suwaidi, A.H.; You, H. Stable isotope chemostratigraphy in lacustrine strata of the Xiagou Formation, Gansu Province, NW China. Geol. Soc. Spec. Publ. 2013, 382, 143–155. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, I.; Gale, A.S.; Jenkyns, H.C.; Pearce, M.A. Secular variation in Late Cretaceous carbon isotopes: A new δ13C carbonate reference curve for the late Cenomanian–Campanian (99.6–70.6 Ma). Geol. Mag. 2006, 143, 561–608. [Google Scholar] [CrossRef] [Green Version]
- Tsikos, H.; Jenkyns, H.C.; Walsworth-Bell, B.; Petrizzo, M.R.; Forster, A.; Kolonic, S.; Erba, E.; Premoli Silva, I.; Baas, M.; Wagner, T.; et al. Carbon-isotope stratigraphy recorded by the Cenomanian-Turonian oceanic anoxic event: Correlation and implications based on three key localities. J. Geol. Soc. 2004, 161, 711–719. [Google Scholar] [CrossRef]
- Jarvis, I.; Lignum, J.S.; Gröcke, D.R.; Jenkyns, H.C.; Pearce, M.A. Black shale deposition, atmospheric CO2 drawdown, and cooling during the Cenomanian-Turonian Oceanic Anoxic Event. Paleoceanography 2011, 26, 3. [Google Scholar] [CrossRef] [Green Version]
- Joo, Y.J.; Sageman, B.B. Cenomanian to Campanian Carbon Isotope Chemostratigraphy from the Western Interior Basin, U.S.A. J. Sediment. Res. 2014, 84, 529–542. [Google Scholar] [CrossRef]
- Navarro-Ramirez, J.P.; Bodin, S.; Immenhauser, A. Ongoing Cenomanian—Turonian heterozoan carbonate production in the neritic settings of Peru. Sediment. Geol. 2016, 331, 78–93. [Google Scholar] [CrossRef]
- Coccioni, R.; Galeotti, S. The mid-Cenomanian Event: Prelude to OAE 2. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2003, 190, 427–440. [Google Scholar] [CrossRef]
- Friedrich, O.; Erbacher, J.; Wilson, P.A.; Moriya, K.; Mutterlose, J. Paleoenvironmental changes across the Mid Cenomanian Event in the tropical Atlantic Ocean (Demerara rise, ODP Leg 207) inferred from bentonic foraminiferal assemblages. Mar. Micropaleontol. 2009, 71, 28–40. [Google Scholar] [CrossRef]
- Stoll, H.M.; Schrag, D.P. High-resolution stable isotope records from the Upper Cretaceous rocks of Italy and Spain: Glacial episodes in a greenhouse planet. Geol. Soc. Am. Bull. 2000, 112, 308–319. [Google Scholar] [CrossRef]
- Heimhofer, U.; Wucherpfennig, N.; Adatte, T.; Schouten, S.; Schneebeli-Hermann, E.; Gardin, S.; Keller, G.; Kentsch, S.; Kujau, A. Vegetation response to exceptional global warmth during Oceanic Anoxic Event 2. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Valle, B.; Dal’ Bó, P.F.; Mendes, M.; Favoreto, J.; Rigueti, A.L.; Borghi, L.; Oliveira Mendonça, J.; Silva, R., Jr. The expression of the Oceanic Anoxic Event 2 (OAE2) in the northeast of Brazil (Sergipe-Alagoas Basin). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 529, 12–23. [Google Scholar] [CrossRef]
- Kuypers, M.M.M.; Pancost, R.D.; Sinninghe Damsté, J.S. A large and abrupt fall in atmospheric CO2 concentration during Cretaceous times. Nature 1999, 399, 342–345. [Google Scholar] [CrossRef]
- Miall, A.D. Fluvial Depositional Systems; Springer International Publishing: Berlin, Germany, 2014; Volume 14, p. 316. [Google Scholar]
- Fosdick, J.C.; Romans, B.W.; Fildani, A.; Bernhardt, A.; Calderón, M.; Graham, S.A. Kinematic evolution of the Patagonian retroarc fold-and-thrust belt and Magallanes foreland basin, Chile and Argentina, 51°30′ S. Geol. Soc.Am. Bull. 2011, 123, 1679–1698. [Google Scholar] [CrossRef]
- Tosolini, A.M.P.; McLoughlin, S.; Wagstaff, B.E.; Cantrill, D.J.; Gallagher, S.J. Cheirolepideacean foliage and pollen from Cretaceous high-latitudes of southeastern Australia. Gondwana Res. 2015, 27, 960–977. [Google Scholar] [CrossRef] [Green Version]
- Coe, M.J.; Dilcher, D.L.; Farlow, J.O.; Jarzen, D.M.; Russell, D.A. Dinosaurs and land plants. In The Origins of Angiosperms and Their Biological Consequences; Friis, E.M., Chaloner, W.G., Crane, P.R., Eds.; Cambridge University Press: New York, NY, USA, 1987; pp. 225–258. [Google Scholar]
- Skog, J.E.; Dilcher, D.L. Lower vascular plants of the Dakota Formation in Kansas and Nebraska, USA. Rev. Palaeobot. Palynol. 1994, 80, 1–18. [Google Scholar] [CrossRef]
- Collinson, M.E. “What use are fossil fern?”—20 years on: With a review of the fossil history of extant Pteridophyte families and genera. In Pteridology in Perspective; Camus, J.M., Gibby, M., Jhons, R.J., Eds.; Kew, Royal Botanical Garden: Richmond, UK, 1996; pp. 349–394. [Google Scholar]
- Jaramillo, C.; Rueda, M.J.; Mora, G. Cenozoic plant diversity in the Neotropics. Science 2006, 311, 1893–1896. [Google Scholar] [CrossRef]
- Barreda, V.D.; Palazzesi, L.; Pujana, R.R.; Panti, C.; Tapia, M.J.; Fernández, D.A.; Noetinger, S. The Gondwanan heritage of the Eocene–Miocene Patagonian floras. J. S. Am. Earth Sci. 2021, 107, 103022. [Google Scholar] [CrossRef]
- Fernández, D.A.; Palazzesi, L.; Estebenet, M.S.G.; Tellería, M.C.; Barreda, V.D. Impact of mid Eocene greenhouse warming on America’s southernmost floras. Commun. Biol. 2021, 4, 1–9. [Google Scholar] [CrossRef]
- Prebble, J.G.; Kennedy, E.M.; Reichgelt, T.; Clowes, C.; Womack, T.; Mildenhall, D.C.; Raine, J.I.; Crouch, E.M. A 100 million year composite pollen record from New Zealand shows maximum angiosperm abundance delayed until Eocene. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2021, 566, 110207. [Google Scholar] [CrossRef]
- Archangelsky, A.; Llorens, M. Palinología de la formación Kachaike, Cretácico Inferior de la Cuenca Austral, provincia de Santa Cruz. I- Esporas lisas y cinguladas. Ameghiniana 2003, 40, 71–80. [Google Scholar]
- Archangelsky, A.; Llorens, M. Palinología de la Formación Kachaike, Cretácico Inferior de la Cuenca Austral, provincia de Santa Cruz. II. Esporas. Ameghiniana 2005, 42, 311–328. [Google Scholar]
- Archangelsky, A.; Llorens, M. Palinología de la Formación Kachaike, Cretácico Inferior de la Cuenca Austral, provincia de Santa Cruz: Granos de polen de Gimnospermas. Ameghiniana 2009, 46, 225–234. [Google Scholar]
- Barreda, V.; Archangelsky, S. The southernmost record of tropical pollen grains in the mid-Cretaceous of Patagonia, Argentina. Cretac. Res. 2006, 27, 778–787. [Google Scholar] [CrossRef]
- Archangelsky, A.; Archangelsky, S.; Poiré, D.; Canessa, N. Registros palinológicos en la Formación Piedra Clavada (Albiano) en su área tipo, provincia de Santa Cruz, Argentina. Rev. Mus. Argent. Cienc. Nat. Nueva Ser. 2008, 10, 185–198. [Google Scholar] [CrossRef] [Green Version]
- Archangelsky, S.; Archangelsky, A.; Cladera, G. Palinología y paleoambientes en el perfil de Bajo Comisión (Cretácico), provincia de Santa Cruz, Argentina. Rev. Mus. Argent. Cienc. Nat. 2012, 14, 23–39. [Google Scholar] [CrossRef] [Green Version]
- Villar de Seoane, L.M.; Archangelsky, S. Taxonomy and biostratigraphy of Cretaceous megaspores from Patagonia, Argentina. Cretac. Res. 2008, 29, 354–372. [Google Scholar] [CrossRef]
- Pérez Loinaze, V.S.; Archangelsky, S.; Cladera, G. Palynostratigraphic study of the Early Cretaceous Río Mayer and Kachaike formations at the Quebrada el Moro section, Austral Basin, southwestern Argentina. Cretac. Res. 2012, 34, 161–171. [Google Scholar] [CrossRef]
- Pérez Loinaze, V.S.; Barreda, V.D.; Archangelsky, S.; Archangelsky, A. Cretaceous angiosperm pollen from the Kachaike formation, south-western Santa Cruz Province, Argentina. Hist. Biol. 2015, 28, 941–951. [Google Scholar] [CrossRef]
- Dettmann, M.E.; Thomson, M.R.A. Cretaceous palynomorphs from the James Ross Island area, Antarctica—A pilot study. Br. Antarct. Surv. 1987, 77, 13–59. [Google Scholar]
- Keating, J.M.; Spencer-Jones, M.; Newham, S. The stratigraphical palynology of the Kotick Point and Whisky Bay formations, Gustav Group (Cretaceous), James Ross Island. Antarct. Sci. 1992, 4, 279–292. [Google Scholar] [CrossRef]
- Mays, C.; Stilwell, J.D. Pollen and spore biostratigraphy of the mid-Cretaceous Tupuangi Formation, Chatham Islands, New Zealand. Rev. Palaeobot. Palynol. 2013, 192, 79–102. [Google Scholar] [CrossRef]
- Mays, C. A Late Cretaceous (Cenomanian–Turonian) south polar palynoflora from the Chatham Islands, New Zealand. Mem. Assoc. Australas. Palaeontol. 2015, 47, 1–96. [Google Scholar]
- Archangelsky, S.; Bellosi, E.S.; Jalfin, G.A.; Perrot, C. Palynology and alluvial facies from the mid-Cretaceous of Patagonia, subsurface of San Jorge Basin, Argentina. Cretac. Res. 1994, 15, 127–142. [Google Scholar] [CrossRef]
- Vallati, P. Middle cretaceous microflora from the Huincul Formation (“dinosaurian beds”) in the Neuquén Basin, Patagonia, Argentina. Palynology 2001, 25, 179–197. [Google Scholar]
- Vallati, P. Las primeras angiospermas en el Cretácico de la Cuenca Neuquina (centro oeste de Argentina): Aspectos geológicos relacionados. Rev. Bras. Paleontol. 2006, 9, 83–92. [Google Scholar] [CrossRef]
- Vallati, P. Paleotropical pollen grains from the Neuquén Group, Patagonia, Argentina. Carnets Géologie 2013, 11, 273–279. [Google Scholar] [CrossRef] [Green Version]
- Burger, D. Palynological observations in the Carpentaria Basin, Queensland. Bur. Miner. Resour. Geol. Geophys. Aust. 1973, 140, 27–44. [Google Scholar]
- Burger, D. Early Cretaceous angiosperms from Queensland, Australia. Rev. Palaeobot. Palynol. 1990, 65, 153–163. [Google Scholar] [CrossRef]
- Burger, D. Early and middle Cretaceous angiosperm pollen grains from Australia. Rev. Palaeobot. Palynol 1993, 78, 183–234. [Google Scholar] [CrossRef]
- Norvick, M.; Burger, D. Palynology of the Cenomanian of Bathurst Island. North. Territ. Austral. Bur. Miner. Resour. Geol. Geophys. 1975, 151, 1–237. [Google Scholar]
- Herngreen, G.F.W.; Kedves, M.; Rovnina, L.V.; Smirnova, S.B. Cretaceous palynofloral provinces. A review. In Palynology, Principles and Applications; Jansonius, J., McGregor, D.C., Eds.; American Association of Stratigraphic Palynologists Foundation: Dallas, TX, USA, 1996; pp. 1157–1188. [Google Scholar]
- Groot, J.J.; Penny, J.S.; Groot, C.R. Plant microfossils and age of the Raritan, Tuscaloosa and Magothy formations of the eastern United States. Palaeontogr. Abt. B 1961, 121–140. [Google Scholar]
- Brenner, G.J. Early angiosperm pollen differentiation in the Albian to Cenomanian deposits of Delaware (USA). Rev. Palaeobot. Palynol. 1967, 1, 219–227. [Google Scholar] [CrossRef]
- Hedlund, R.W. Palynology of the Red Branch Member of the Woodbine Formation (Cenomanian), Bryan County, Oklahoma; Oklahoma Geological Survey; University of Oklahoma: Norman, OK, USA, 1966; Volume 113, pp. 1–69. [Google Scholar]
- Peyrot, D.; Barrón, E.; Comas Rengifo, M.J.; Barroso Barcenilla, F.; Feist-Burkhardt, S. Palinología del tránsito Cenomaniense/Turoniense en la sección de Puentedey (Burgos, España). Coloq. Paleontol. 2008, 58, 101–161. [Google Scholar]
- Pierce, R.L. Lower Upper Cretaceous plant microfossils from Minnesota. Minn. Geol. Surv. 1961, 42, 1–86. [Google Scholar]
- Galloway, J.M.; Sweet, A.R.; Pugh, A.; Schröder-Adams, C.J.; Swindles, G.T.; Haggart, J.W.; Embry, A.F. Correlating middle Cretaceous palynological records from the Canadian High Arctic based on a section from the Sverdrup Basin and samples from the Eclipse Trough. Palynology 2012, 36, 277–302. [Google Scholar] [CrossRef]
- Sulphur, K. Quantitative Palynological Analyses of Albian–Cenomanian (Lower to Upper Cretaceous) Strata in the Sverdrup Basin: Insights into Paleoecology, Paleoclimatology and Palynostratigraphy. Master’s Thesis, University of Calgary, Calgary, AB, Canada, 2015. [Google Scholar]
- Peyrot, D.; Barrón, E.; Polette, F.; Batten, D.J.; Néraudeau, D. Early Cenomanian palynofloras and inferred resiniferous forests and vegetation types in Charentes (southwestern France). Cretac. Res. 2019, 94, 168–189. [Google Scholar] [CrossRef] [Green Version]
- Žítt, J.; Nekvalisová, O.; Bosák, P.; Svobodová, M.; Stemproková-Jírová, D.; Stasstny, M. Rocky Coast facies of the Cenomanian-Turonian boundary interval at Velim (Bohemian Cretaceous Basin, Czech Republic). Second Part Věstník Státního Geol. Ust. 1997, 72, 141–155. [Google Scholar]
- Svobodová, M.; Méon, H.; Pacltová, B. Characteristics of palynospectra of the Upper Cenomanian-Lower Turonian (anoxic facies) of the Bohemian and Vocontian Basins. Bull. Czech Geol. Surv. 1998, 73, 229–251. [Google Scholar]
- Méon, H.; Guignard, G.; Pacltová, B.; Svobodová, M. Normapolles. Comparison between central and southwestern Europe during the Cenomanian and Turonian: Evolution of biodiversity and paleoenvironment. Bull. Soc. Géol. Fr. 2004, 175, 579–593. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Pedofeatures and Compositional Data of the Paleosols of the Mata Amarilla Formation | ||||
|---|---|---|---|---|
| Paleosol-Type | Macromorphological Pedofeatures | Micromorphological Pedofeatures | Clay-Mineral Assemblages (XRD Analysis) | Geochemical Data |
| Histosols | Stacks of thin Oe-A-Bg horizons, dark grey to black mottles of carbonaceous remains, orange and purplish-blue mottles, rhizoliths, fossil roots | Enaulic, close porphyric or double space to open and close porphyric c/f-related distribution. Abundant coarse organic components. Undifferentiated b-fabric disguised by Fe-oxides and organic matter. Simple-packing voids, chambers, channels, and plant residues. Fe-Mn nodules | S-P assemblage (palygorskite-rich association with dominant smectite), >S-K assemblage (kaolinite-rich association with smectite and other clays), >S assemblage (smectite-dominated association) | Average of Al/bases ratio: 0.54, Ba/Sr ratio: 1.05; Al/Si ratio: 0.16; CIA-K: 79; PWI: 32 |
| Vertisols | Thick well-developed profiles with A-Bss; Btss-Bssg-Cg horizons, greenish-gray matrix with low chromas, slickensides, angular peds, rhizoliths, mottles, Fe-Mn nodules | Double space to open porphyric c/f-related distribution. Undifferentiated b-fabric disguised by Fe-oxides and organic matter or cross-straited b-fabric. Compound-packing voids, chambers, channels, and rhizoliths. Laminated and non-laminated clay coatings; Fe-Mn nodules | S assemblage (smectite-dominated association), >S-K assemblage (kaolinite-rich association with smectite and other clays) | Average of Al/bases ratio: 0.54, Ba/Sr ratio: 1.43; Al/Si ratio: 0.17; CIA-K: 80; PWI: 35 |
| Inceptisols | Thin moderately to poorly developed and stacked profiles with A-Bw or B/C-C horizons, poorly developed horizonation, gley colors, rhizoliths, mottles, Fe-Mn nodules, massive or poor ped developments | Gefuric to chitonic and monic c/f-related distribution. Undifferentiated b-fabric. Simple-packing voids and chambers. Thin non-laminated clay coatings; Fe-Mn nodules | S assemblage (smectite-dominated association) | Average of Al/bases ratio: 0.65, Ba/Sr ratio: 1.67; Al/Si ratio: 0.14; CIA-K: 76; PWI: 39 |
| Vertic Alfisols | Thick well-developed profiles with A-A/E-Bt-Bss horizons, dark gray co-lors, well-structured Bt horizons with prismatic peds and Bss horizons with angular peds, abundant cutans, slickensides, mottles, Fe-Mn nodules | Monic, chitonic to double- or simple-space porphyric c/f-related distribution. Undifferentiated or cross-striated b-fabric. Compound-packing voids, channels, and rhizoliths. Laminated to microlaminated clay infillings | S assemblage (smectite-dominated association) | Average of Al/bases ratio: 0.51, Ba/Sr ratio: 1.13; Al/Si ratio: 0.20; CIA-K: 81; PWI: 37 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varela, A.N.; Raigemborn, M.S.; Santamarina, P.E.; Lizzoli, S.; Adatte, T.; Heimhofer, U. Carbon Isotopic Signature and Organic Matter Composition of Cenomanian High-Latitude Paleosols of Southern Patagonia. Geosciences 2021, 11, 378. https://doi.org/10.3390/geosciences11090378
Varela AN, Raigemborn MS, Santamarina PE, Lizzoli S, Adatte T, Heimhofer U. Carbon Isotopic Signature and Organic Matter Composition of Cenomanian High-Latitude Paleosols of Southern Patagonia. Geosciences. 2021; 11(9):378. https://doi.org/10.3390/geosciences11090378
Chicago/Turabian StyleVarela, Augusto Nicolás, María Sol Raigemborn, Patricio Emmanuel Santamarina, Sabrina Lizzoli, Thierry Adatte, and Ulrich Heimhofer. 2021. "Carbon Isotopic Signature and Organic Matter Composition of Cenomanian High-Latitude Paleosols of Southern Patagonia" Geosciences 11, no. 9: 378. https://doi.org/10.3390/geosciences11090378
APA StyleVarela, A. N., Raigemborn, M. S., Santamarina, P. E., Lizzoli, S., Adatte, T., & Heimhofer, U. (2021). Carbon Isotopic Signature and Organic Matter Composition of Cenomanian High-Latitude Paleosols of Southern Patagonia. Geosciences, 11(9), 378. https://doi.org/10.3390/geosciences11090378