
Are Past Sea-Ice Reconstructions Based on Planktonic Foraminifera Realistic? Study of the Last 50 ka as a Test to Validate Reconstructed Paleohydrography Derived from Transfer Functions Applied to Their Fossil Assemblages

 ,
,
Abstract
:1. Introduction
2. Past Hydrographical Quantifications Based on Planktonic Foraminifera
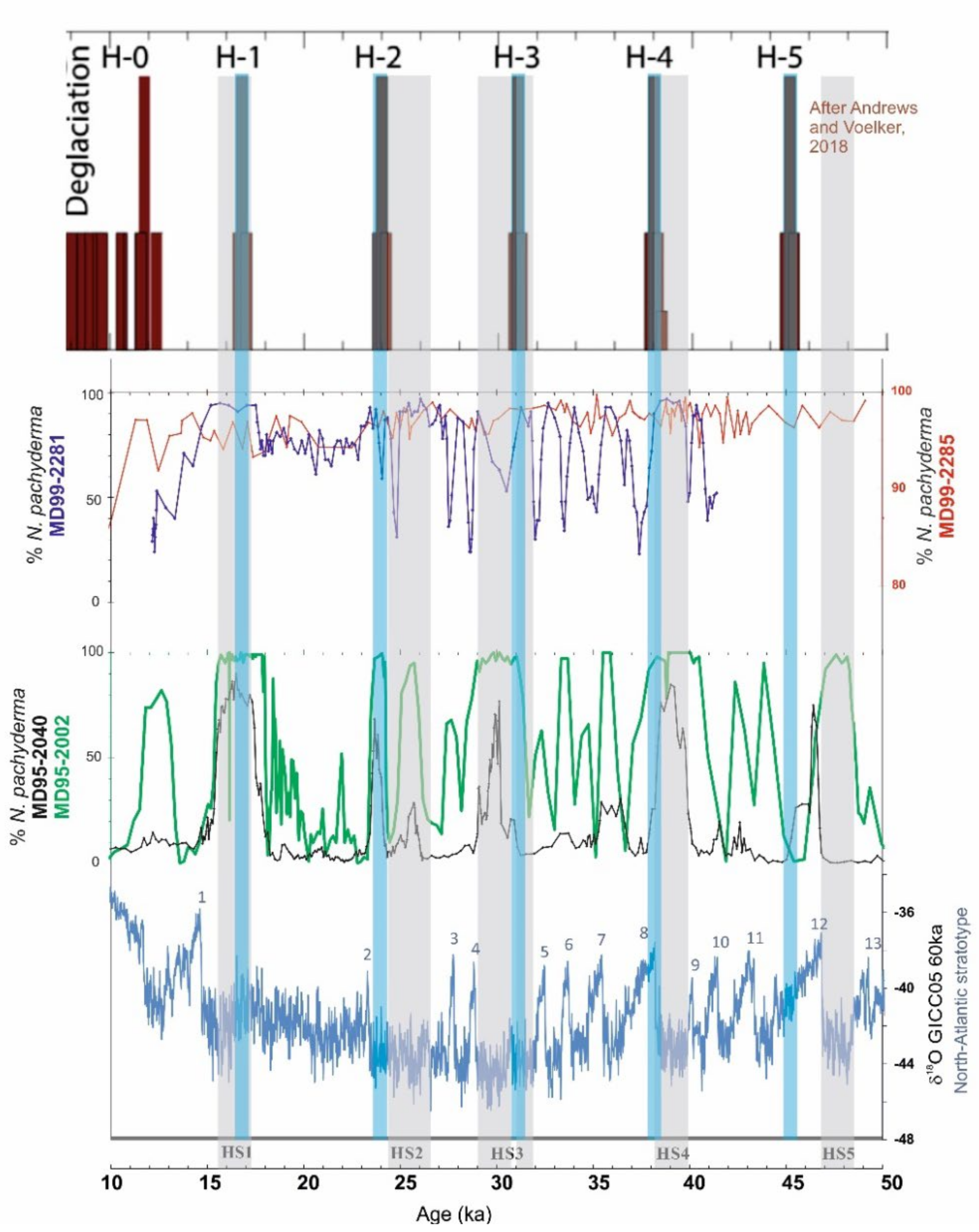
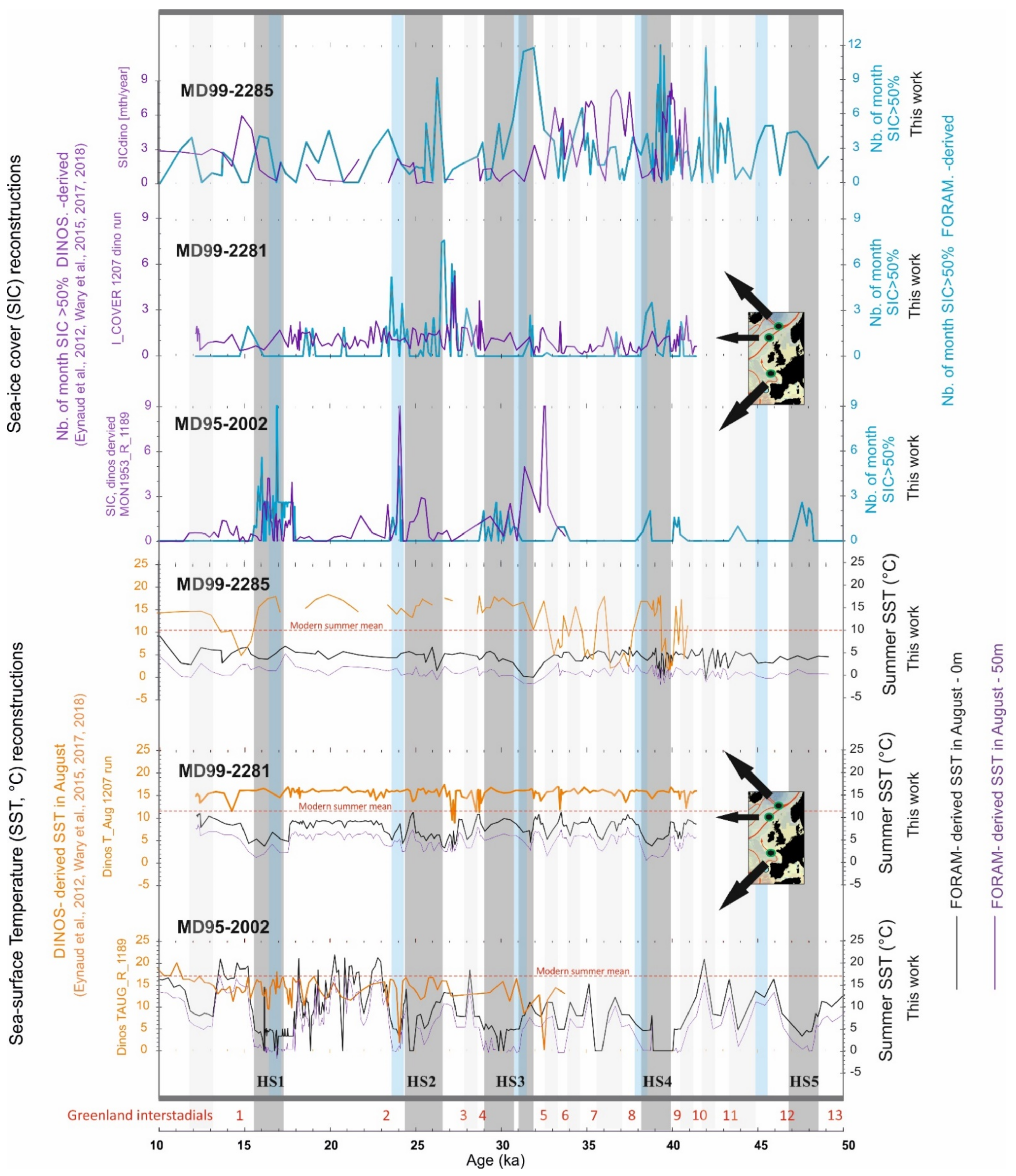
3. Heinrich Events and Their PF-Derived Signatures
3.1. Basic Features and Knowledge
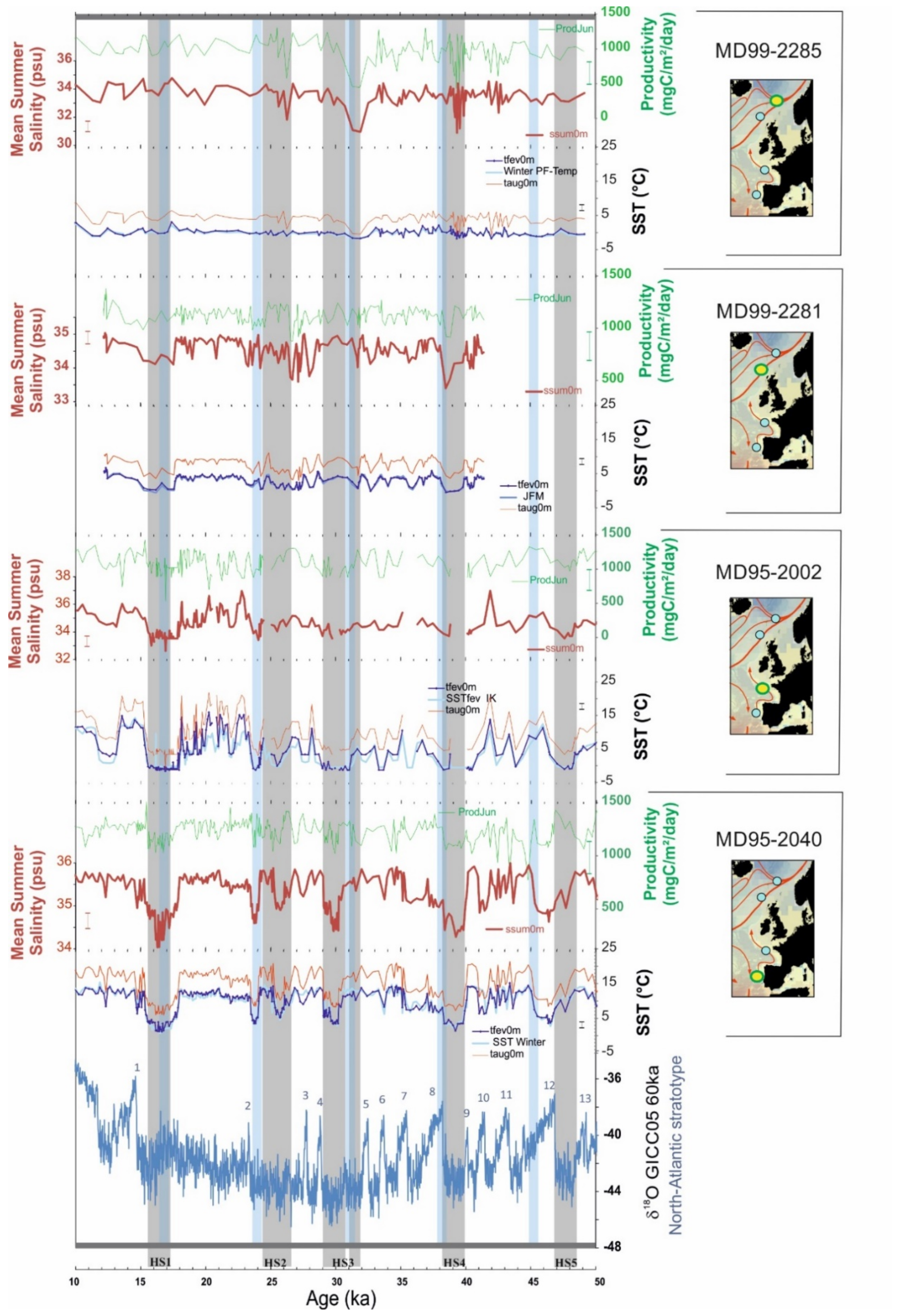
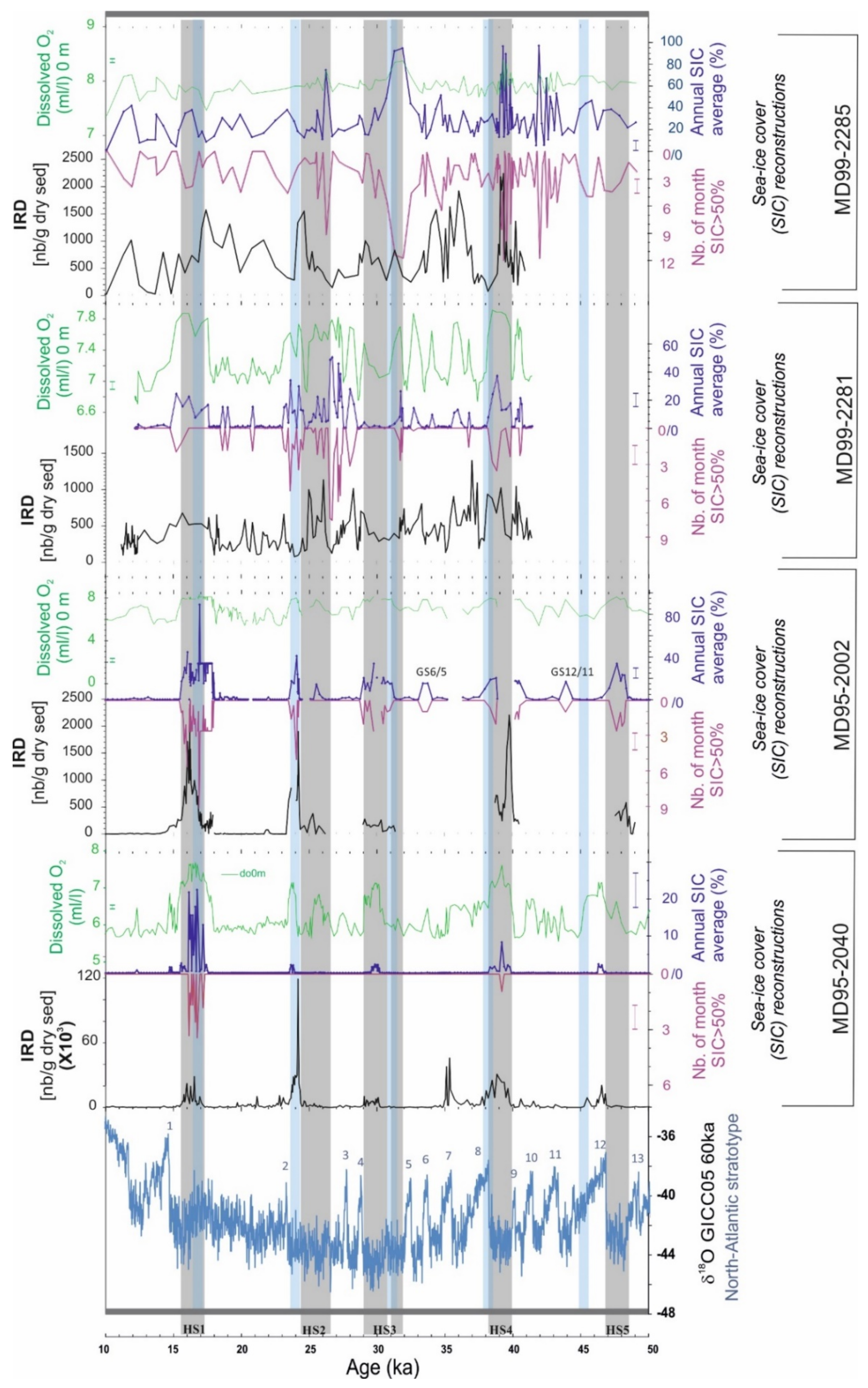
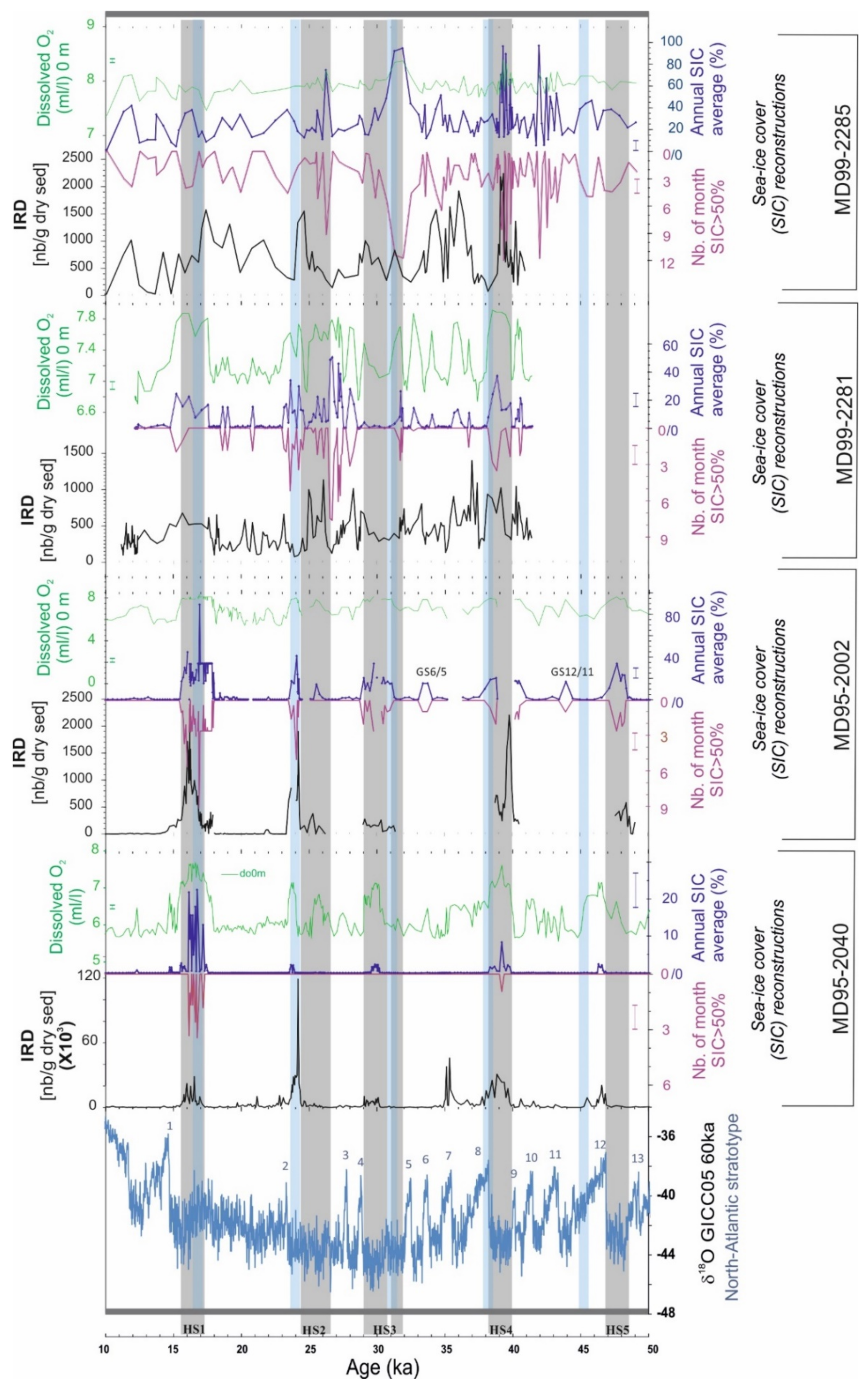
3.2. New Hydrographic Signals to Better Understand HEs
4. Discussion: Is the Reconstruction of New Hydrographical Parameters with PF Progress?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Phleger, F.B. Foraminifera of submarine cores from the continental slope. GSA Bull. 1939, 50, 1395–1422. [Google Scholar] [CrossRef]
- Schiebel, R.; Smart, S.M.; Jentzen, A.; Jonkers, L.; Morard, R.; Meilland, J.; Michel, E.; Coxall, H.K.; Hull, P.M.; de Garidel-Thoron, T.; et al. Advances in planktonic foraminifer research: New perspectives for paleoceanography. Rev. Micropaleontol. 2018, 61, 113–138. [Google Scholar] [CrossRef]
- CLIMAP Projectc Members. The Surface of the Ice-Age Earth. Science 1976, 191, 1131–1137. [Google Scholar] [CrossRef]
- Pflaumann, U.; Duprat, J.; Pujol, C.; Labeyrie, L.D. SIMMAX: A modern analog technique to deduce Atlantic sea surface temperatures from planktonic foraminifera in deep-sea sediments. Paleoceanography 1996, 11, 15–35. [Google Scholar] [CrossRef]
- Siccha, M.; Kucera, M. ForCenS, a curated database of planktonic foraminifera census counts in marine surface sediment samples. Sci. Data 2017, 4, 170109. [Google Scholar] [CrossRef] [PubMed]
- Kucera, M.; Rosell-Melé, A.; Schneider, R.; Waelbroeck, C.; Weinelt, M. Multiproxy approach for the reconstruction of the glacial ocean surface (MARGO). Quat. Sci. Rev. 2005, 24, 813–819. [Google Scholar] [CrossRef]
- Kucera, M.; Weinelt, M.; Kiefer, T.; Pflaumann, U.; Hayes, A.; Weinelt, M.; Chen, M.-T.; Mix, A.C.; Barrows, T.T.; Cortijo, E.; et al. Reconstruction of sea-surface temperatures from assemblages of planktonic foraminifera: Multi-technique approach based on geographically constrained calibration data sets and its application to glacial Atlantic and Pacific Oceans. Quat. Sci. Rev. 2005, 24, 951–998. [Google Scholar] [CrossRef]
- Nguyen, T.M.P.; Speijer, R.P. A new procedure to assess dissolution based on experiments on Pliocene–Quaternary foraminifera (ODP Leg 160, Eratosthenes Seamount, Eastern Mediterranean). Mar. Micropaleontol. 2014, 106, 22–39. [Google Scholar] [CrossRef]
- Greco, M.; Jonkers, L.; Kretschmer, K.; Bijma, J.; Kucera, M. Depth habitat of the planktonic foraminifera Neogloboquadrina pachyderma in the northern high latitudes explained by sea-ice and chlorophyll concentrations. Biogeosciences 2019, 16, 3425–3437. [Google Scholar] [CrossRef] [Green Version]
- Greco, M.; Morard, R.; Kucera, M. Single-cell metabarcoding reveals biotic interactions of the Arctic calcifier Neogloboquadrina pachyderma with the eukaryotic pelagic community. J. Plankton Res. 2021, 43, 113–125. [Google Scholar] [CrossRef]
- Telford, R.J.; Li, C.; Kucera, M. Mismatch between the depth habitat of planktonic foraminifera and the calibration depth of SST transfer functions may bias reconstructions. Clim. Past 2013, 9, 859–870. [Google Scholar] [CrossRef] [Green Version]
- Jonkers, L.; Kučera, M. Sensitivity to species selection indicates the effect of nuisance variables on marine microfossil transfer functions. Clim. Past 2019, 15, 881–891. [Google Scholar] [CrossRef] [Green Version]
- Ruddiman, W.F. Late Quaternary deposition of ice-rafted sand in the subpolar North Atlantic (lat 40° to 65°N). GSA Bull. 1977, 88, 1813–1827. [Google Scholar] [CrossRef]
- Eynaud, F.; De Abreu, L.; Voelker, A.; Schönfeld, J.; Salgueiro, E.; Turon, J.-L.; Penaud, A.; Toucanne, S.; Naughton, F.; Goñi, M.F.S.; et al. Position of the Polar Front along the western Iberian margin during key cold episodes of the last 45 ka. Geochem. Geophys. Geosyst. 2009, 10, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Guiot, J.; de Vernal, A. Chapter Thirteen: Transfer Functions: Methods for Quantitative Paleoceanography Based on Microfossils, In Developments in Marine Geology; Hillaire-Marcel, C., de Vernal, A., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2007; pp. 523–563. [Google Scholar]
- de Vernal, A.; Radi, T.; Zaragosi, S.; Van Nieuwenhove, N.; Rochon, A.; Allan, E.; De Schepper, S.; Eynaud, F.; Head, M.J.; Limoges, A.; et al. Distribution of common modern dinoflagellate cyst taxa in surface sediments of the Northern Hemisphere in relation to environmental parameters: The new n = 1968 database. Mar. Micropaleontol. 2019, 159, 101796. [Google Scholar] [CrossRef]
- Eynaud, F.; Mary, Y.; Zumaque, J.; Wary, M.; Gasparotto, M.-C.; Swingedouw, D.; Colin, C. Compiling multiproxy quantitative hydrographic data from Holocene marine archives in the North Atlantic: A way to decipher oceanic and climatic dynamics and natural modes? Glob. Planet. Chang. 2018, 170, 48–61. [Google Scholar] [CrossRef]
- Mary, Y.; Eynaud, F.; Colin, C.; Rossignol, L.; Brocheray, S.; Mojtahid, M.; Garcia, J.; Peral, M.; Howa, H.; Zaragosi, S.; et al. Changes in Holocene meridional circulation and poleward Atlantic flow: The Bay of Biscay as a nodal point. Clim. Past 2017, 13, 201–216. [Google Scholar] [CrossRef] [Green Version]
- Matsuzaki, K.M.R.; Eynaud, F.; Malaizé, B.; Grousset, F.E.; Tisserand, A.; Rossignol, L.; Charlier, K.; Jullien, E. Paleoceanography of the Mauritanian margin during the last two climatic cycles: From planktonic foraminifera to African climate dynamics. Mar. Micropaleontol. 2011, 79, 67–79. [Google Scholar] [CrossRef]
- Sánchez Goñi, M.F.; Bakker, P.; Desprat, S.; Carlson, A.E.; Van Meerbeeck, C.J.; Peyron, O.; Naughton, F.; Fletcher, W.J.; Eynaud, F.; Rossignol, L.; et al. European climate optimum and enhanced Greenland melt during the Last Interglacial. Geology 2012, 40, 627–630. [Google Scholar] [CrossRef]
- Wary, M.; Eynaud, F.; Sabine, M.; Zaragosi, S.; Rossignol, L.; Malaizé, B.; Palis, E.; Zumaque, J.; Caulle, C.; Penaud, A.; et al. Stratification of surface waters during the last glacial millennial climatic events: A key factor in subsurface and deep-water mass dynamics. Clim. Past 2015, 11, 1507–1525. [Google Scholar] [CrossRef] [Green Version]
- Eynaud, F.; Malaizé, B.; Zaragosi, S.; de Vernal, A.; Scourse, J.; Pujol, C.; Cortijo, E.; Grousset, F.E.; Penaud, A.; Toucanne, S.; et al. New constraints on European glacial freshwater releases to the North Atlantic Ocean. Geophys. Res. Lett. 2012, 39, L15601. [Google Scholar] [CrossRef] [Green Version]
- Eynaud, F.; Rossignol, L.; Gasparotto, M.-C. Planktic foraminifera throughout the Pleistocene: From cell to populations to past marine hydrology. In Foraminifera: Classification, Biology, and Evolutionary Significance; Georgescu, M.D., Ed.; Nova Science Publishers: New York, NY, USA, 2013. [Google Scholar]
- MARGO Program Members; Waelbroeck, C.; Paul, A.; Kucera, , M.; Rosell-Melé, A.; Weinelt, M.; Schneider, R.; Mix, A.C.; Abelmann, A.; Armand, L.; et al. Constraints on the magnitude and patterns of ocean cooling at the Last Glacial Maximum. Nat. Geosci. 2009, 2, 127–132. [Google Scholar] [CrossRef]
- Morey, A.E.; Mix, A.C.; Pisias, N.G. Planktonic foraminiferal assemblages preserved in surface sediments correspond to multiple environment variables. Quat. Sci. Rev. 2005, 24, 925–950. [Google Scholar] [CrossRef]
- Trenberth, K.E. What are the Seasons? Bull. Am. Meteorol. Soc. 1983, 64, 1276–1282. [Google Scholar] [CrossRef]
- Bé, A.W.H.; Tolderlund, D.S. Chapter 6. Distribution and ecology of living planktonic foraminifera in surface waters of the Atlantic and Indian oceans, In Micropaleontology of Oceans; Funnell, B.M., Riedel, W.R., Eds.; Cambridge University Press: London, UK, 1971; pp. 105–149. [Google Scholar]
- Lombard, F.; Labeyrie, L.D.; Michel, E.; Bopp, L.; Cortijo, E.; Retailleau, S.; Howa, H.; Jorissen, F. Modelling planktic foraminifer growth and distribution using an ecophysiological multi-species approach. Biogeosciences 2011, 8, 853–873. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, M.; Braconnot, P.; Bopp, L.; Mariotti, V.; Roy, T.; Woillez, M.-N.; Caubel, A.; Foujols, M.-A.; Guilyardi, E.; Khodri, M.; et al. Mid-Holocene and last glacial maximum climate simulations with the IPSL model: Part II: Model-data comparisons. Clim. Dyn. 2012, 40, 2469–2495. [Google Scholar] [CrossRef]
- Locarnini, R.A.; Mishonov, A.V.; Antonov, J.I.; Boyer, T.P.; Garcia, H.E.; Baranova, O.K.; Zweng, M.M.; Paver, C.R.; Reagan, J.R.; Johnson, D.R.; et al. World Ocean Atlas 2013, Volume 1: Temperature; NOAA Atlas NESDIS 73. 2012; p. 40. Available online: https://www.nodc.noaa.gov/OC5/woa13/pubwoa13.html (accessed on 20 July 2021).
- de Abreu, L.; Shackleton, N.J.; Schönfeld, J.; Hall, M.; Chapman, M. Millennial-scale oceanic climate variability off the Western Iberian margin during the last two glacial periods. Mar. Geol. 2003, 196, 1–20. [Google Scholar] [CrossRef]
- Voelker, A.H.L.; de Abreu, L. A.H.L.; de Abreu, L. A Review of Abrupt Climate Change Events in the Northeastern Atlantic Ocean (Iberian Margin): Latitudinal, Longitudinal, and Vertical Gradients. In Geophysical Monograph Series; Rashid, H., Polyak, L., Mosley-Thompson, E., Eds.; American Geophysical Union: Washington, DC, USA, 2011; pp. 15–37. [Google Scholar]
- Auffret, G.; Zaragosi, S.; Dennielou, B.; Cortijo, E.; Van Rooij, D.; Grousset, F.; Pujol, C.; Eynaud, F.; Siegert, M. Terrigenous fluxes at the Celtic margin during the last glacial cycle. Mar. Geol. 2002, 188, 79–108. [Google Scholar] [CrossRef]
- Grousset, F.E.; Pujol, C.; Labeyrie, L.; Auffret, G.; Boelaert, A. Were the North Atlantic Heinrich events triggered by the behavior of the European ice sheets? Geology 2000, 28, 123–126. [Google Scholar] [CrossRef]
- Grousset, F.E.; Cortijo, E.; Huon, S.; Herve, L.; Burdloff, D.; Duprat, J.; Weber, O.; Richter, T. Zooming in on Heinrich layers. Paleoceanography 2001, 16, 240–259. [Google Scholar] [CrossRef]
- Grousset, F.E.; Cortijo, E.; Huon, S.; Herve, L.; Richter, T.; Burdloff, D.; Duprat, J.; Weber, O. Abundance of planktonic foraminifera, Neogloboquadrina pachyderma δ¹⁸O values and summer SST for sediment core SU90-09. PANGAEA 2002. [Google Scholar] [CrossRef]
- Zaragosi, S.; Eynaud, F.; Pujol, C.; Auffret, G.A.; Turon, J.-L.; Garlan, T. Initiation of the European deglaciation as recorded in the northwestern Bay of Biscay slope environments (Meriadzek Terrace and Trevelyan Escarpment): A multi-proxy approach. Earth Planet. Sci. Lett. 2001, 188, 493–507. [Google Scholar] [CrossRef]
- Ménot, G.; Bard, E.; Rostek, F.; Weijers, J.W.H.; Hopmans, E.C.; Schouten, S.; Damsté, J.S.S. Early Reactivation of European Rivers During the Last Deglaciation. Science 2006, 313, 1623–1625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eynaud, F.; Zaragosi, S.; Scourse, J.D.; Mojtahid, M.; Bourillet, J.F.; Hall, I.R.; Penaud, A.; Locascio, M.; Reijonen, A. Deglacial laminated facies on the NW Eu-ropean continental margin: The hydrographic significance of British–Irish Ice Sheet deglaciation and Fleuve Manche paleoriver discharges. Geochem. Geophys. Geosyst. 2007, 8, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Zumaque, J.; Eynaud, F.; Zaragosi, S.; Marret, F.; Matsuzaki, K.M.; Kissel, C.; Roche, D.M.; Malaizé, B.; Michel, E.; Billy, I.; et al. An ocean-ice coupled response during the last glacial: Aview from a marine isotopic stage 3 record South of the Faeroe Shetland Gateway. Clim. Past 2012, 8, 1997–2017. [Google Scholar] [CrossRef] [Green Version]
- Caulle, C.; Penaud, A.; Eynaud, F.; Zaragosi, S.; Roche, D.M.; Michel, E.; Boulay, S.; Richter, T. Sea-surface hydrographical conditions off South Faeroes and within the North-Eastern North Atlantic through MIS 2: The response of dinocysts. J. Quat. Sci. 2013, 28, 217–228. [Google Scholar] [CrossRef]
- Wary, M.; Eynaud, F.; Rossignol, L.; Lapuyade, J.; Gasparotto, M.-C.; Londeix, L.; Malaizé, B.; Castéra, M.-H.; Charlier, K. Norwegian Sea warm pulses during Dansgaard-Oeschger stadials: Zooming in on these anomalies over the 35–41 ka cal BP interval and their impacts on proximal European ice-sheet dynamics. Quat. Sci. Rev. 2016, 151, 255–272. [Google Scholar] [CrossRef]
- Wary, M.; Eynaud, F.; Rossignol, L.; Zaragosi, S.; Sabine, M.; Castera, M.-H.; Billy, I. The southern Norwegian Sea during the last 45 ka: Hydrographical reorganizations under changing ice-sheet dynamics. J. Quat. Sci. 2017, 32, 908–922. [Google Scholar] [CrossRef]
- Wary, M.; Eynaud, F.; Swingedouw, D.; Masson-Delmotte, V.; Matthiessen, J.; Kissel, C.; Zumaque, J.; Rossignol, L.; Jouzel, J. Regional seesaw between the North Atlantic and Nordic Seas during the last glacial abrupt climate events. Clim. Past 2017, 13, 729–739. [Google Scholar] [CrossRef] [Green Version]
- Andrews, J.T.; Voelker, A.H. “Heinrich events” (& sediments): A history of terminology and recommendations for future usage. Quat. Sci. Rev. 2018, 187, 31–40. [Google Scholar] [CrossRef]
- Svensson, A.; Andersen, K.K.; Bigler, M.; Clausen, H.B.; Dahl-Jensen, D.; Davies, S.M.; Johnsen, S.J.; Muscheler, R.; Parrenin, F.; Rasmussen, S.O.; et al. A 60 000 year Greenland stratigraphic ice core chronology. Clim. Past 2008, 4, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Wolff, E.; Chappellaz, J.; Blunier, T.; Rasmussen, S.O.; Svensson, A. Millennial-scale variability during the last glacial: The ice core record. Quat. Sci. Rev. 2010, 29, 2828–2838. [Google Scholar] [CrossRef]
- Waelbroeck, C.; Lougheed, B.C.; Riveiros, N.V.; Missiaen, L.; Pedro, J.; Dokken, T.; Hajdas, I.; Wacker, L.; Abbott, P.; Dumoulin, J.-P.; et al. Consistently dated Atlantic sediment cores over the last 40 thousand years. Sci. Data 2019, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Heinrich, H. Origin and Consequences of Cyclic Ice Rafting in the Northeast Atlantic Ocean During the Past 130,000 Years. Quat. Res. 1988, 29, 142–152. [Google Scholar] [CrossRef]
- Andrews, J.T.; Jennings, A.E.; Kerwin, M.; Kirby, M.; Manley, W.; Miller, G.H.; Bond, G.; MacLean, B. A Heinrich-like event, H-0 (DC-0): Source(s) for detrital carbonate in the North Atlantic during the Younger Dryas chronozone. Paleoceanography 1995, 10, 943–952. [Google Scholar] [CrossRef]
- Raymo, M.E.; Ganley, K.; Carter, S.D.; Oppo, D.W.; McManus, J. Millennial-scale climate instability during the early Pleistocene epoch. Nature 1998, 392, 699–702. [Google Scholar] [CrossRef]
- Channell, J.; Hodell, D. Magnetic signatures of Heinrich-like detrital layers in the Quaternary of the North Atlantic. Earth Planet. Sci. Lett. 2013, 369–370, 260–270. [Google Scholar] [CrossRef]
- McManus, J.F.; Bond, G.C.; Broecker, W.S.; Johnsen, S.; Labeyrie, L.; Higgins, S. High-resolution climate records from the North Atlantic during the last interglacial. Nature 1994, 371, 326–329. [Google Scholar] [CrossRef]
- Eynaud, F.; Turon, J.; Sánchez-Goñi, M.; Gendreau, S. Dinoflagellate cyst evidence of ‘Heinrich-like events’ off Portugal during the Marine Isotopic Stage 5. Mar. Micropaleontol. 2000, 40, 9–21. [Google Scholar] [CrossRef]
- Pérez-Mejías, C.; Moreno, A.; Sancho, C.; Martín-García, R.; Spötl, C.; Cacho, I.; Cheng, H.; Edwards, R.L. Orbital-to-millennial scale climate variability during Marine Isotope Stages 5 to 3 in northeast Iberia. Quat. Sci. Rev. 2019, 224, 105946. [Google Scholar] [CrossRef]
- Dansgaard, W.; Johnsen, S.J.; Clausen, H.B.; Dahl-Jensen, D.; Gundestrup, N.S.; Hammer, C.U.; Hvidberg, C.S.; Steffensen, J.P.; Sveinbjörnsdottir, A.E.; Jouzel, J.; et al. Evidence for general instability of past climate from a 250-kyr ice-core record. Nature 1993, 364, 218–220. [Google Scholar] [CrossRef]
- Kageyama, M.; Roche, D.M.; Combourieu Nebout, N.; Alvarez-Solas, J. Chap. 29: Rapid Climate Variability: Description and Mechanisms. In Paleoclimatology, Frontiers in Earth Sciences; Ramstein, G., Ed.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Grousset, F.E.; Labeyrie, L.; Sinko, J.A.; Cremer, M.; Bond, G.; Duprat, J.; Cortijo, E.; Huon, S. Patterns of Ice-Rafted Detritus in the Glacial North Atlantic (40–55°N). Paleoceanography 1993, 8, 175–192. [Google Scholar] [CrossRef]
- Nave, S.; Labeyrie, L.; Gherardi, J.; Caillon, N.; Cortijo, E.; Kissel, C.; Abrantes, F. Primary productivity response to Heinrich events in the North Atlantic Ocean and Norwegian Sea. Paleoceanography 2007, 22, PA3216. [Google Scholar] [CrossRef]
- Mariotti, V.; Bopp, L.; Tagliabue, A.; Kageyama, M.; Swingedouw, D. Marine productivity response to Heinrich events: A model-data comparison. Clim. Past 2012, 8, 1581–1598. [Google Scholar] [CrossRef] [Green Version]
- Ausín, B.; Hodell, D.A.; Cutmore, A.; Eglinton, T.I. The impact of abrupt deglacial climate variability on productivity and upwelling on the southwestern Iberian margin. Quat. Sci. Rev. 2020, 230, 106139. [Google Scholar] [CrossRef]
- Bard, E.; Rostek, F.; Turon, J.-L.; Gendreau, S. Hydrological Impact of Heinrich Events in the Subtropical Northeast Atlantic. Science 2000, 289, 1321–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turon, J.-L.; Lezine, A.-M.; Denèfle, M. Land–sea correlations for the last glaciation inferred from a pollen and dinocyst record from the Portuguese margin. Quat. Res. 2003, 59, 88–96. [Google Scholar] [CrossRef]
- Dowdeswell, J.A.; Whittington, R.J.; Jennings, A.E.; Andrews, J.T.; Mackensen, A.; Marienfeld, P. An origin for laminated glacimarine sediments through sea-ice build-up and suppressed iceberg rafting. Sedimentology 2000, 47, 557–576. [Google Scholar] [CrossRef]
- Lichey, C.; Hellmer, H.H. Modeling giant-iceberg drift under the influence of sea ice in the Weddell Sea, Antarctica. J. Glaciol. 2001, 47, 452–460. [Google Scholar] [CrossRef] [Green Version]
- Sancetta, C. Primary production in the glacial North Atlantic and North Pacific oceans. Nature 1992, 360, 249–251. [Google Scholar] [CrossRef]
- Stern, A.A.; Johnson, E.; Holland, D.M.; Wagner, T.J.; Wadhams, P.; Bates, R.; Abrahamsen, E.P.; Nicholls, K.W.; Crawford, A.; Gagnon, J.; et al. Wind-driven upwelling around grounded tabular icebergs. J. Geophys. Res. Ocean. 2015, 120, 5820–5835. [Google Scholar] [CrossRef] [Green Version]
- Jongma, J.I.; Driesschaert, E.; Fichefet, T.; Goosse, H.; Renssen, H. The effect of dynamic–thermodynamic icebergs on the Southern Ocean climate in a three-dimensional model. Ocean Model. 2009, 26, 104–113. [Google Scholar] [CrossRef]
- Romero, O.E.; LeVay, L.J.; McClymont EL, L.; Müller, J.; Cowan, E.A. Orbital and Suborbital Variations of Productivity and Sea Surface Conditions in the Gulf of Alaska during the Past 54,000 years: Impact of Iron Fertilization by Icebergs and Meltwater. Earth and Space Science Open Archive. 2021. Available online: https://www.essoar.org/doi/10.1002/essoar.10507545.1 (accessed on 20 August 2021).
- Garcia, H.E.; Boyer, T.P.; Locarnini, R.A.; Antonov, J.I.; Mishonov, A.V.; Baranova, O.K.; Zweng, M.M.; Reagan, J.; Johnson, D.R. World Ocean Atlas 2013. Volume 3, Dissolved Oxygen, Apparent Oxygen Utilization, and Oxygen Saturation. Available online: https://www.nodc.noaa.gov/OC5/woa13/pubwoa13.html (accessed on 20 August 2021).
- Ziemen, F.A.; Kapsch, M.-L.; Klockmann, M.; Mikolajewicz, U. Heinrich events show two-stage climate response in transient glacial simulations. Clim. Past 2019, 15, 153–168. [Google Scholar] [CrossRef] [Green Version]
- Marchal, O.; Cacho, I.; Stocker, T.F.; Grimalt, J.O.; Calvo, E.; Martrat, B.; Shackleton, N.; Vautravers, M.; Cortijo, E.; van Kreveld, S.; et al. Apparent long-term cooling of the sea surface in the northeast Atlantic and Mediterranean during the Holocene. Quat. Sci. Rev. 2002, 21, 455–483. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Core | Latitude | Longitude | Water Depth (m) | Mean Modern SST (°C) ** | Relevant References, with Those Using Transfer Functions Derived from PF in Bold | Transfer Function Method | ||
|---|---|---|---|---|---|---|---|---|
| Annual | Winter-JFM | Summer-JAS | ||||||
| MD95-2040 | 40.580 | –9.860 | 2465 | 16.2 | 14.0 | 18.6 | de Abreu et al., 2003 [31]; Eynaud et al., 2009 [14]; Voelker and de Abreu, 2011 [32] | MAT (number of spectra in the database n = 1066), SIMMAX28, distance-weighted, data available at: http://doi.pangaea.de/10.1594/PANGAEA.737449, 20 September 2021 |
| MD95-2002 | 47.450 | –8.530 | 2174 | 13.9 | 11.4 | 17.2 | Auffret et al., 2002 [33]; Grousset et al., 2000 [34], 2001 [35], 2002 [36]; Zaragosi et al., 2001 [37]; Menot et al., 2005 [38]; Eynaud et al., 2007 [39]; 2012 [22]; 2018 [17] | MAT (number of spectra in the database: n = 556, n = 1007 for publication after 2005) and Imbrie & Kipp method * (number of spectra in the database: n = 556) |
| MD99-2281 | 60.342 | –9.456 | 1197 | 9.7 | 8.3 | 11.5 | Zumaque et al., 2012 [40]; Caulle et al., 2013 [41]; Wary et al., 2015 [21] | MAT (number of spectra in the database: n = 1007) |
| MD99-2285 | 62.694 | –3.572 | 885 | 8.0 | 6.2 | 10.4 | Wary et al., 2015 [21], 2016 [42], 2017 [43], 2018 [44] | MAT (number of spectra in the database: n = 1007) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eynaud, F.; Zaragosi, S.; Wary, M.; Woussen, E.; Rossignol, L.; Voisin, A. Are Past Sea-Ice Reconstructions Based on Planktonic Foraminifera Realistic? Study of the Last 50 ka as a Test to Validate Reconstructed Paleohydrography Derived from Transfer Functions Applied to Their Fossil Assemblages. Geosciences 2021, 11, 409. https://doi.org/10.3390/geosciences11100409
Eynaud F, Zaragosi S, Wary M, Woussen E, Rossignol L, Voisin A. Are Past Sea-Ice Reconstructions Based on Planktonic Foraminifera Realistic? Study of the Last 50 ka as a Test to Validate Reconstructed Paleohydrography Derived from Transfer Functions Applied to Their Fossil Assemblages. Geosciences. 2021; 11(10):409. https://doi.org/10.3390/geosciences11100409
Chicago/Turabian StyleEynaud, Frédérique, Sébastien Zaragosi, Mélanie Wary, Emilie Woussen, Linda Rossignol, and Adrien Voisin. 2021. "Are Past Sea-Ice Reconstructions Based on Planktonic Foraminifera Realistic? Study of the Last 50 ka as a Test to Validate Reconstructed Paleohydrography Derived from Transfer Functions Applied to Their Fossil Assemblages" Geosciences 11, no. 10: 409. https://doi.org/10.3390/geosciences11100409
APA StyleEynaud, F., Zaragosi, S., Wary, M., Woussen, E., Rossignol, L., & Voisin, A. (2021). Are Past Sea-Ice Reconstructions Based on Planktonic Foraminifera Realistic? Study of the Last 50 ka as a Test to Validate Reconstructed Paleohydrography Derived from Transfer Functions Applied to Their Fossil Assemblages. Geosciences, 11(10), 409. https://doi.org/10.3390/geosciences11100409