Bacterial Communities of Novaya Zemlya Archipelago Ice and Permafrost

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
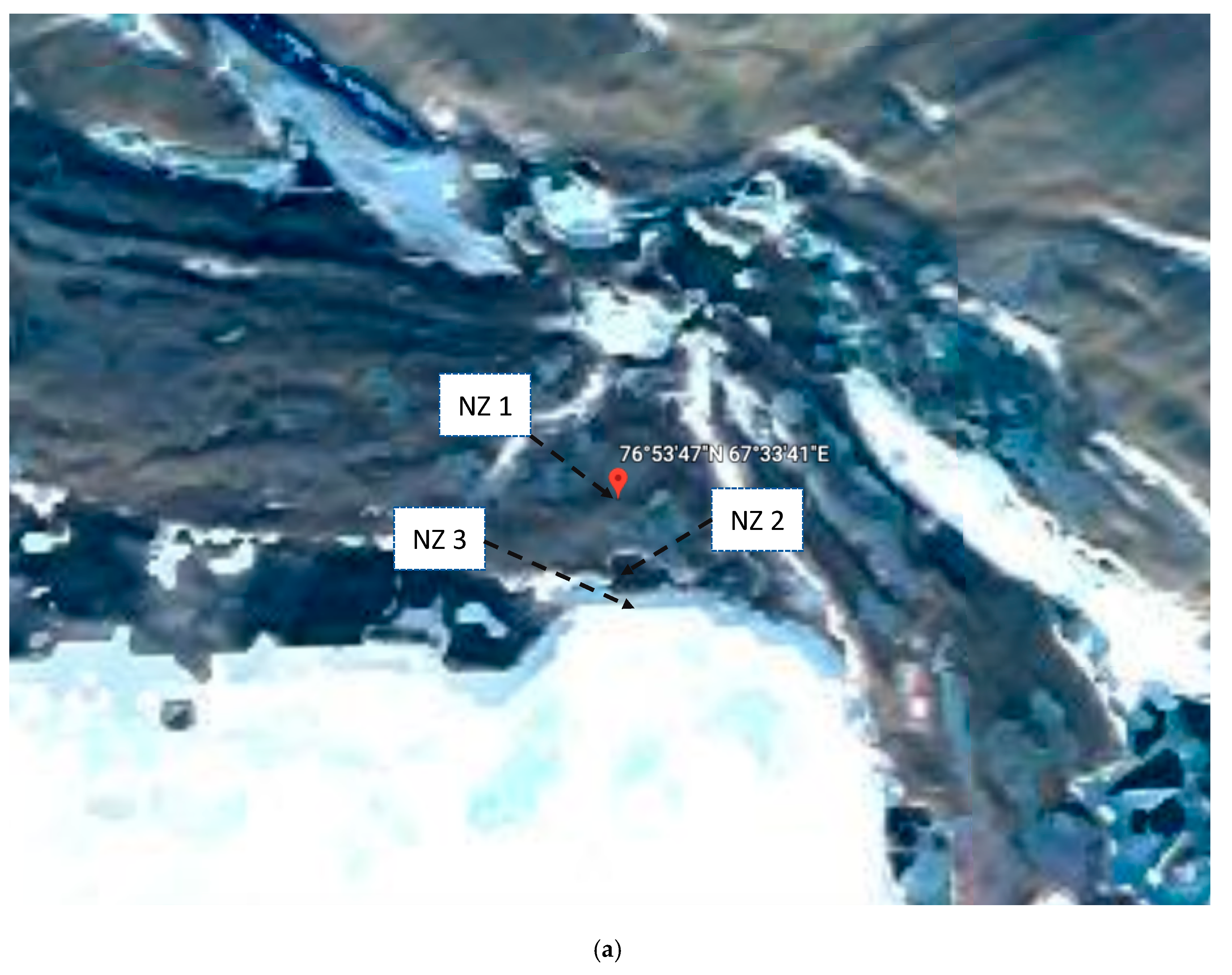
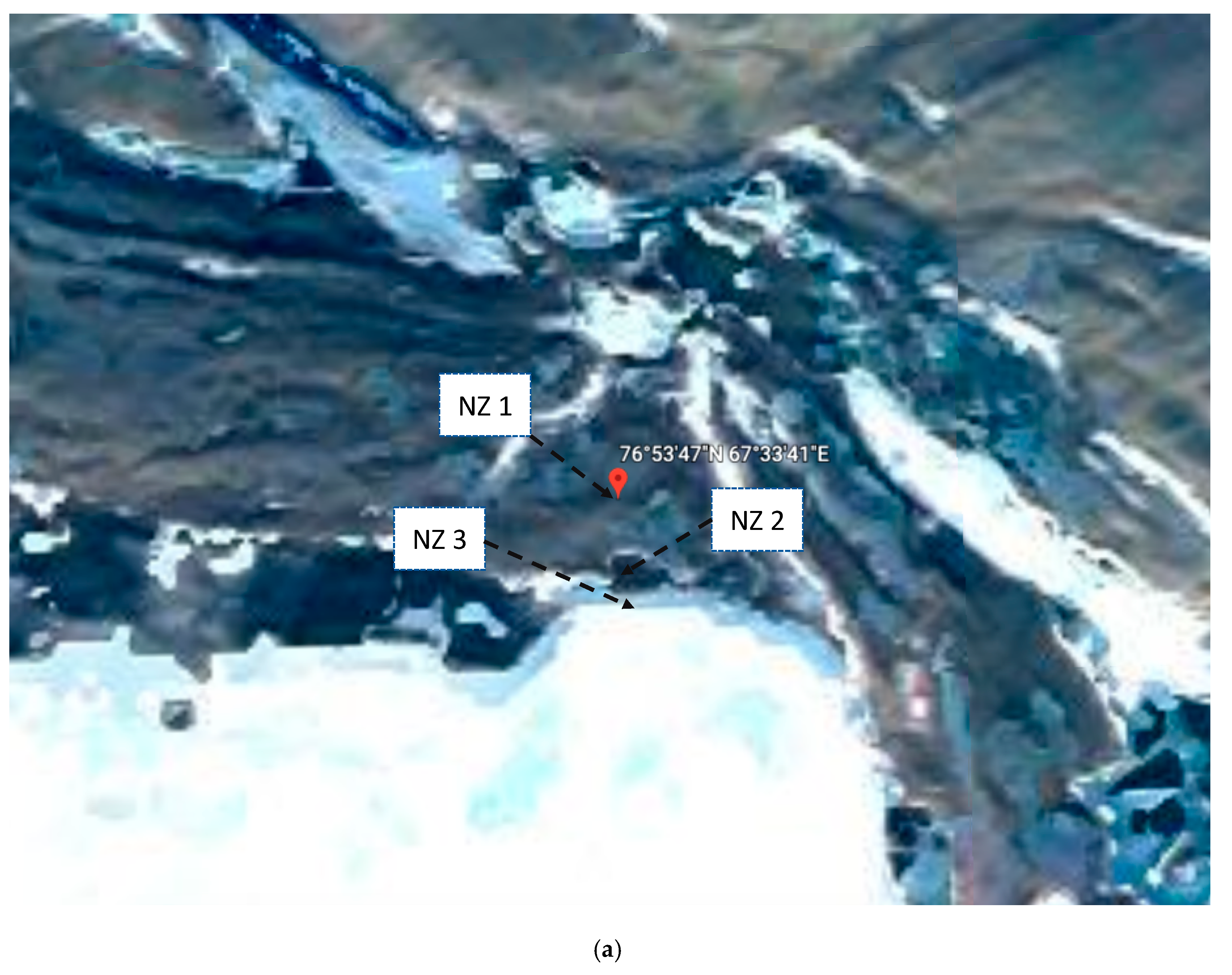
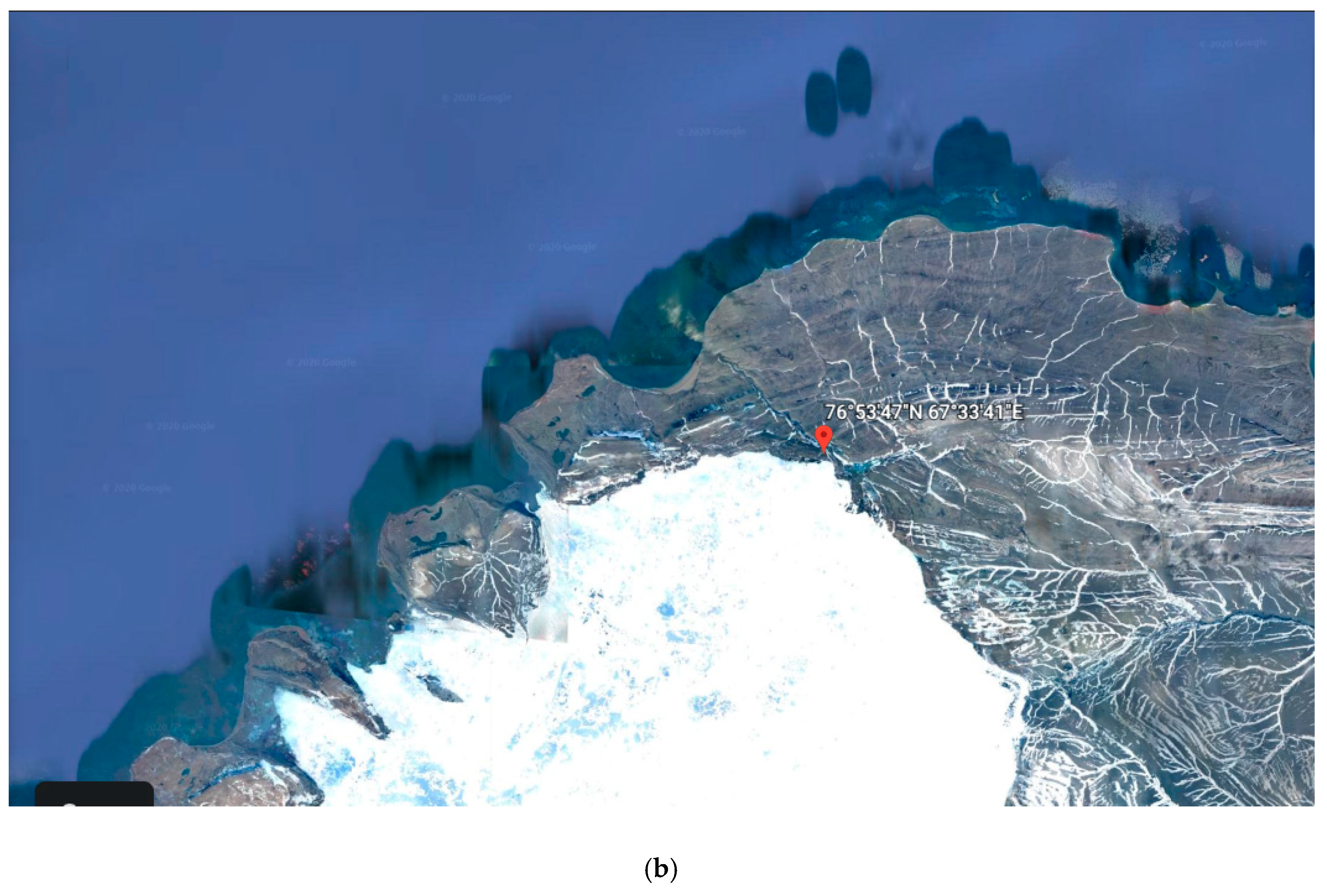
2.1. Sampling
2.2. Epifluorescence Microscopy
2.3. Total DNA Extraction and 16S rRNA Genes Illumina Sequencing
2.4. Fluorescence in situ Hybridization
2.5. Multisubstrate Testing
2.6. Isolation of Culturable Bacterial Communities
2.7. Physiological Assays of Isolated Strains
2.8. Pure Cultures Identification
2.9. Data Analysis
3. Results
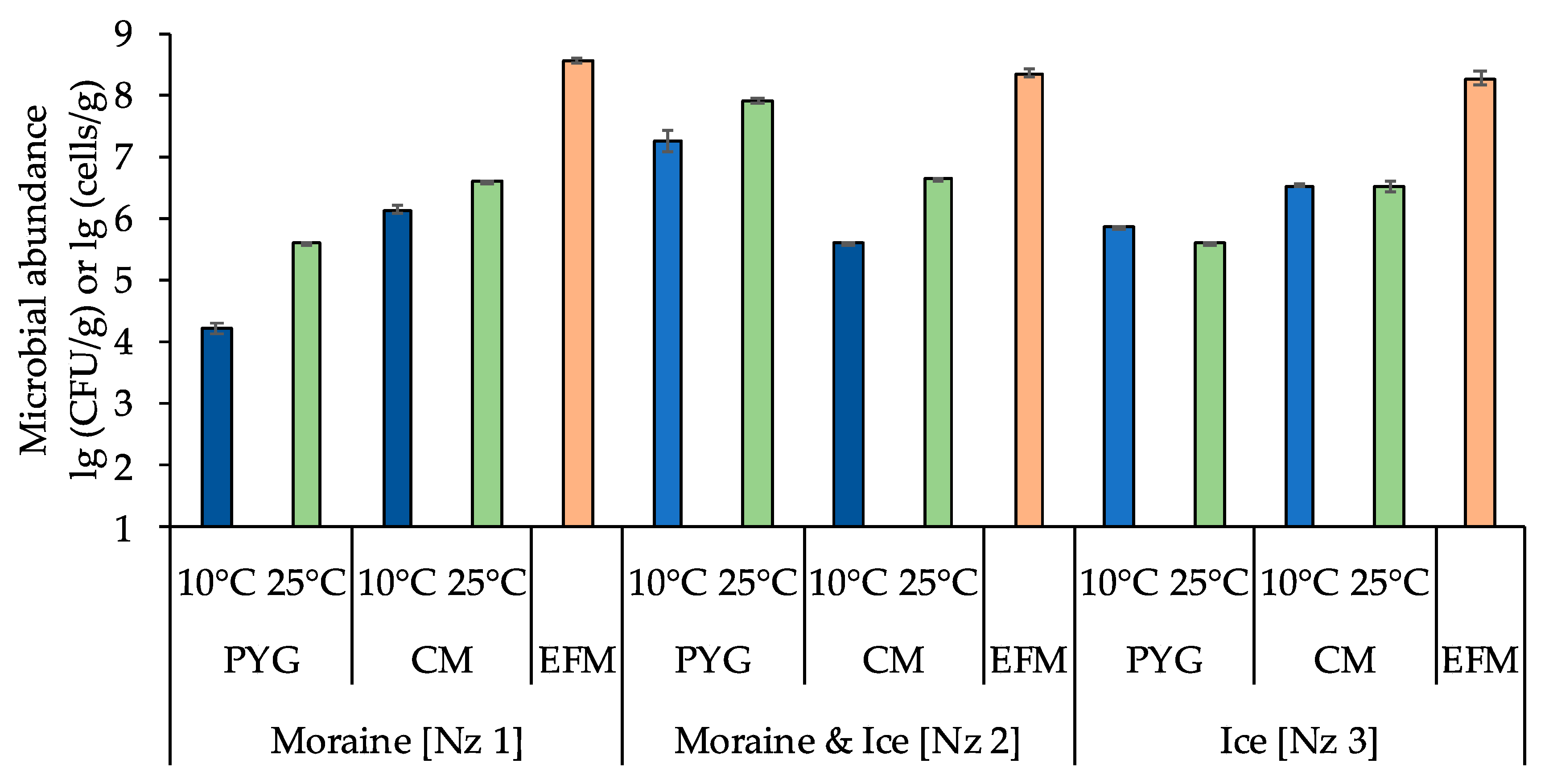
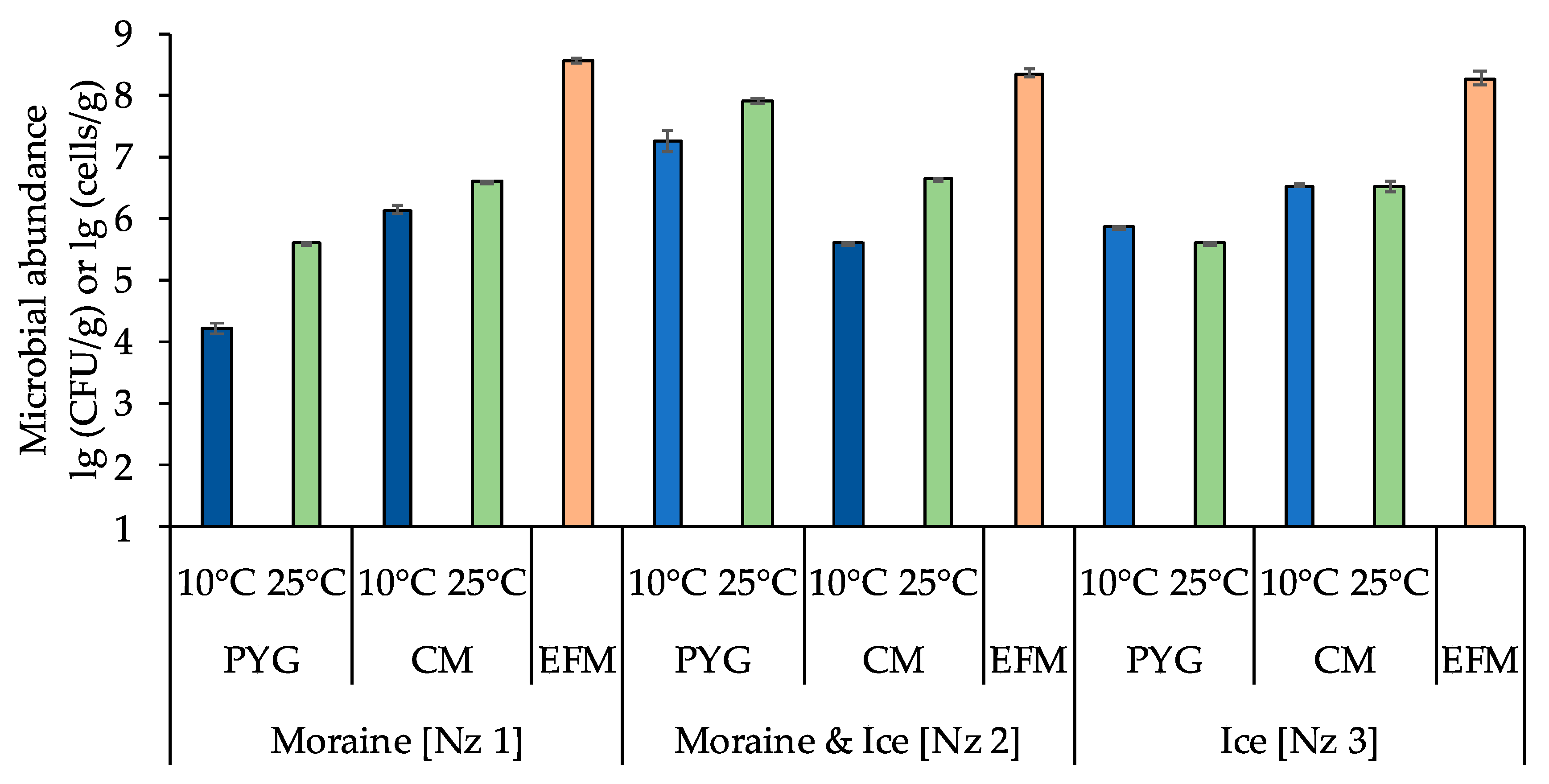
3.1. Total Numbers of Prokaryotic Cells and Numbers of Culturable Bacteria
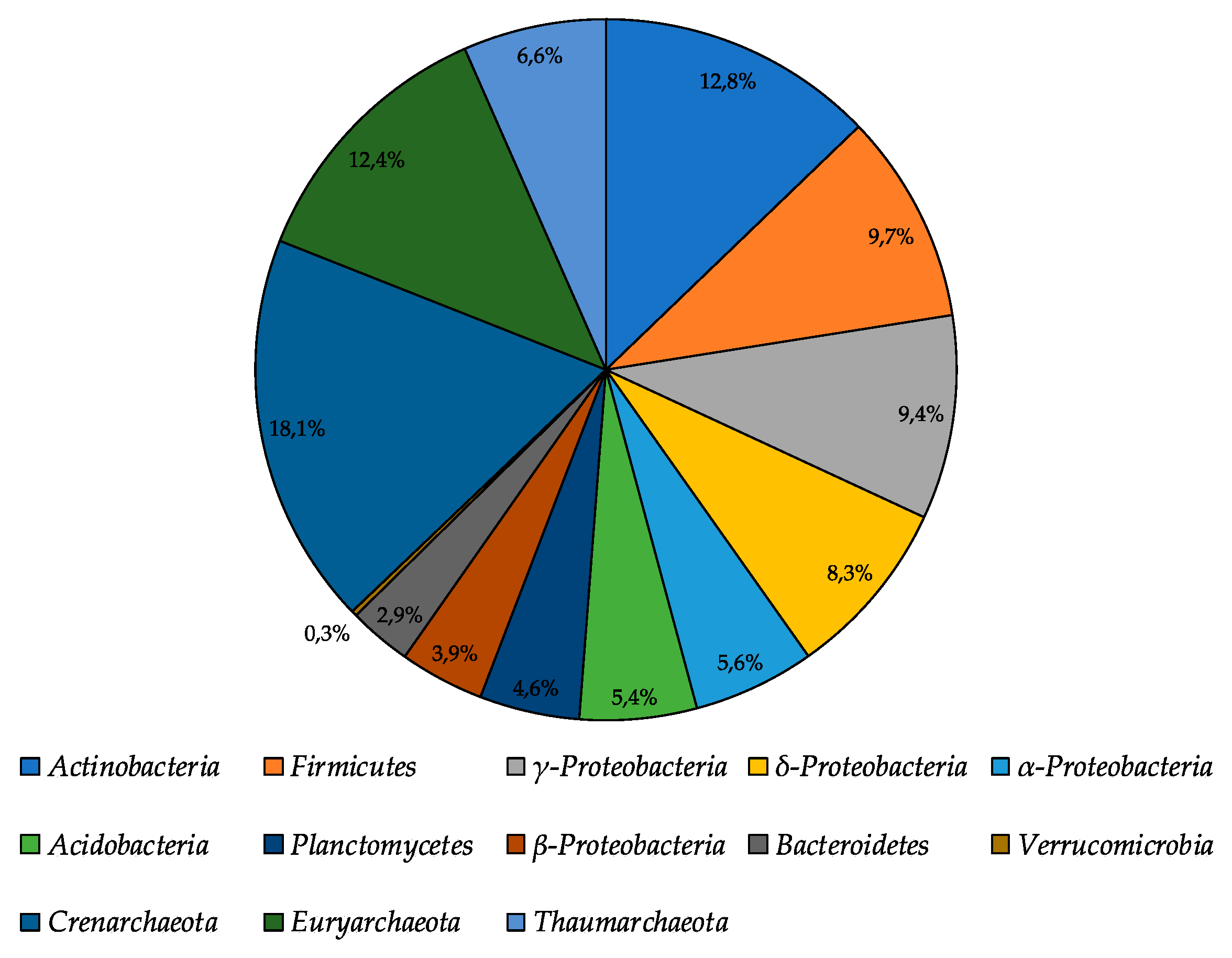
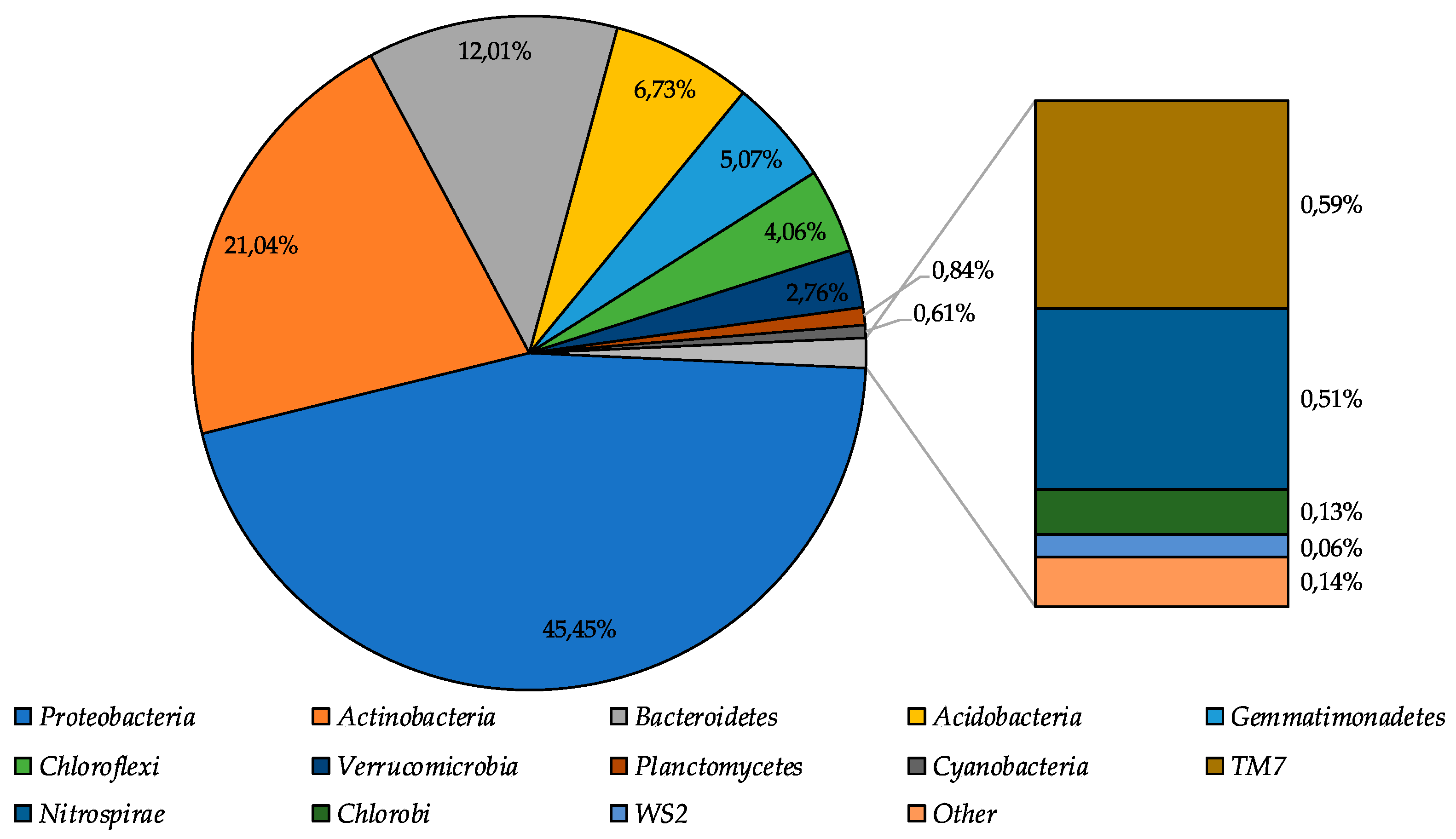
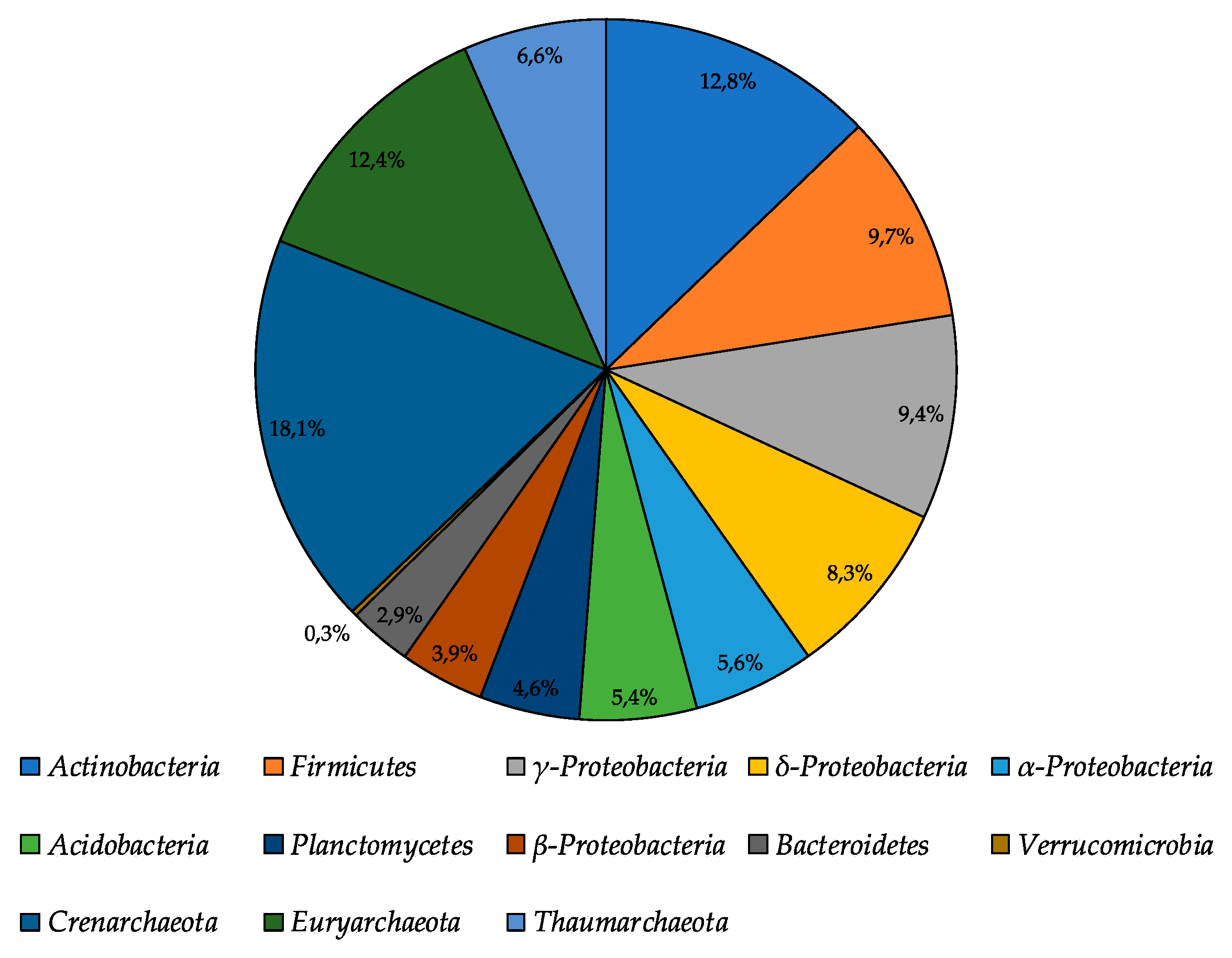
3.2. 16S rRNA Illumina Sequencing Based Bacterial Community Structure
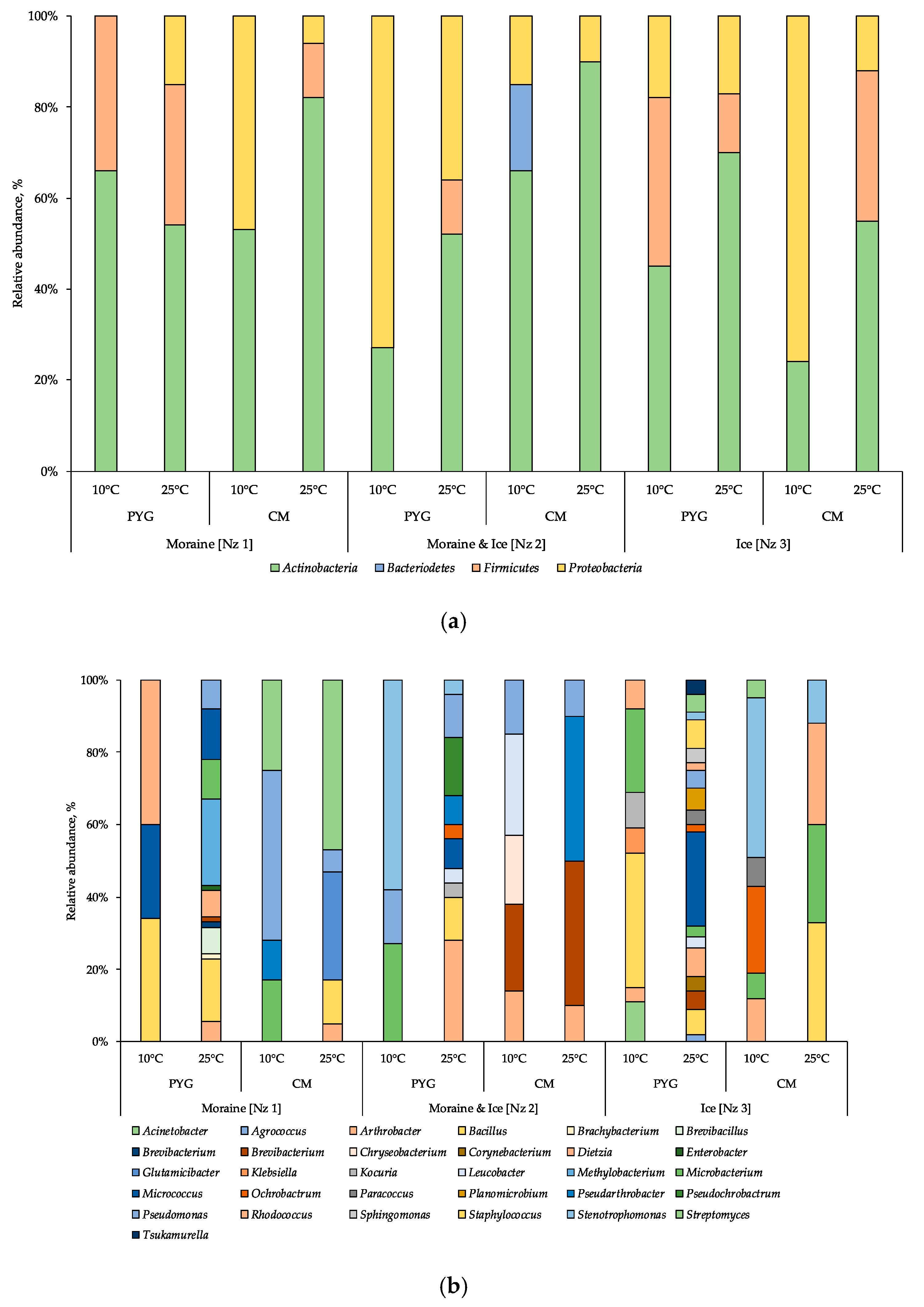
3.3. Culturable Bacterial Communities’ Structure
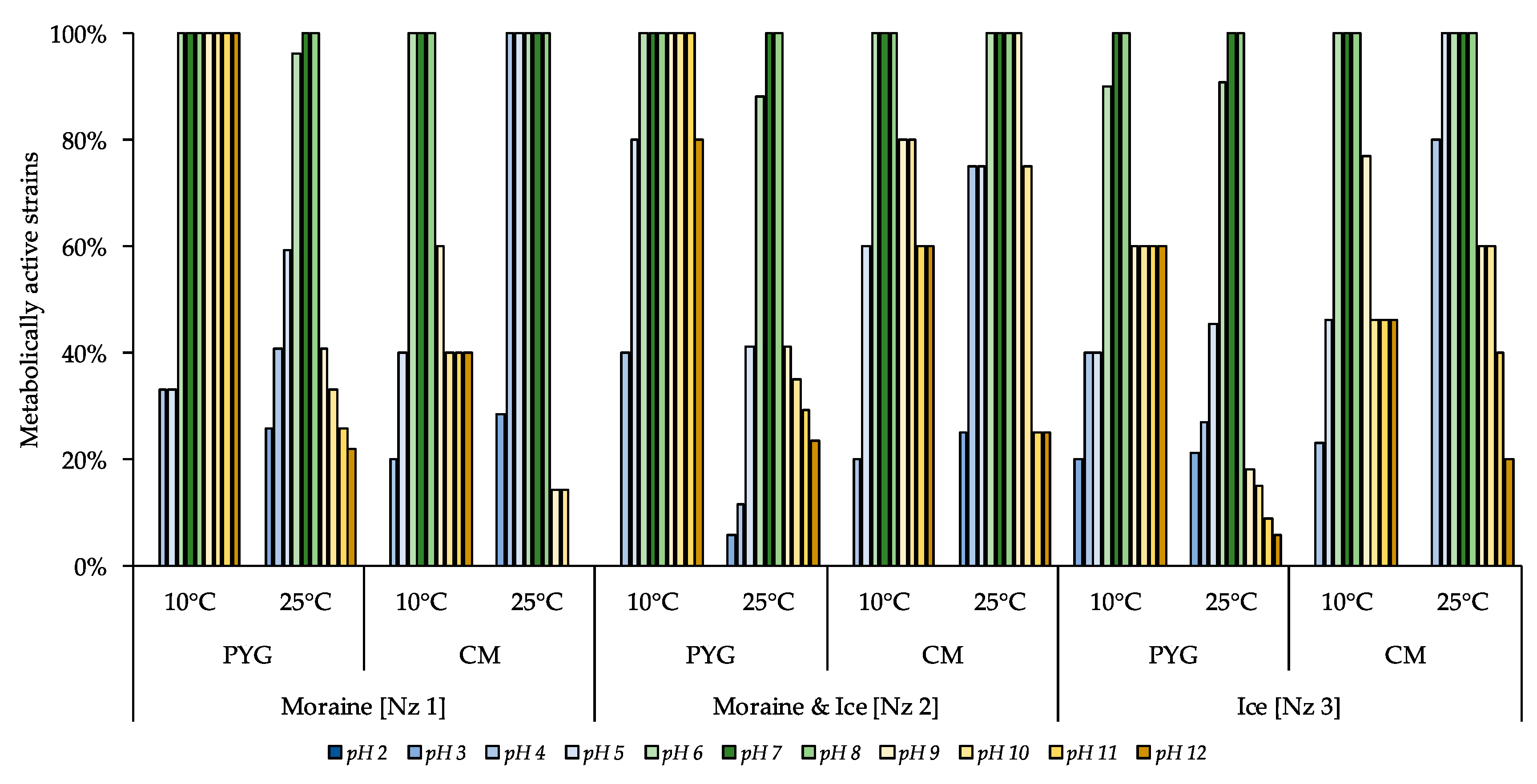
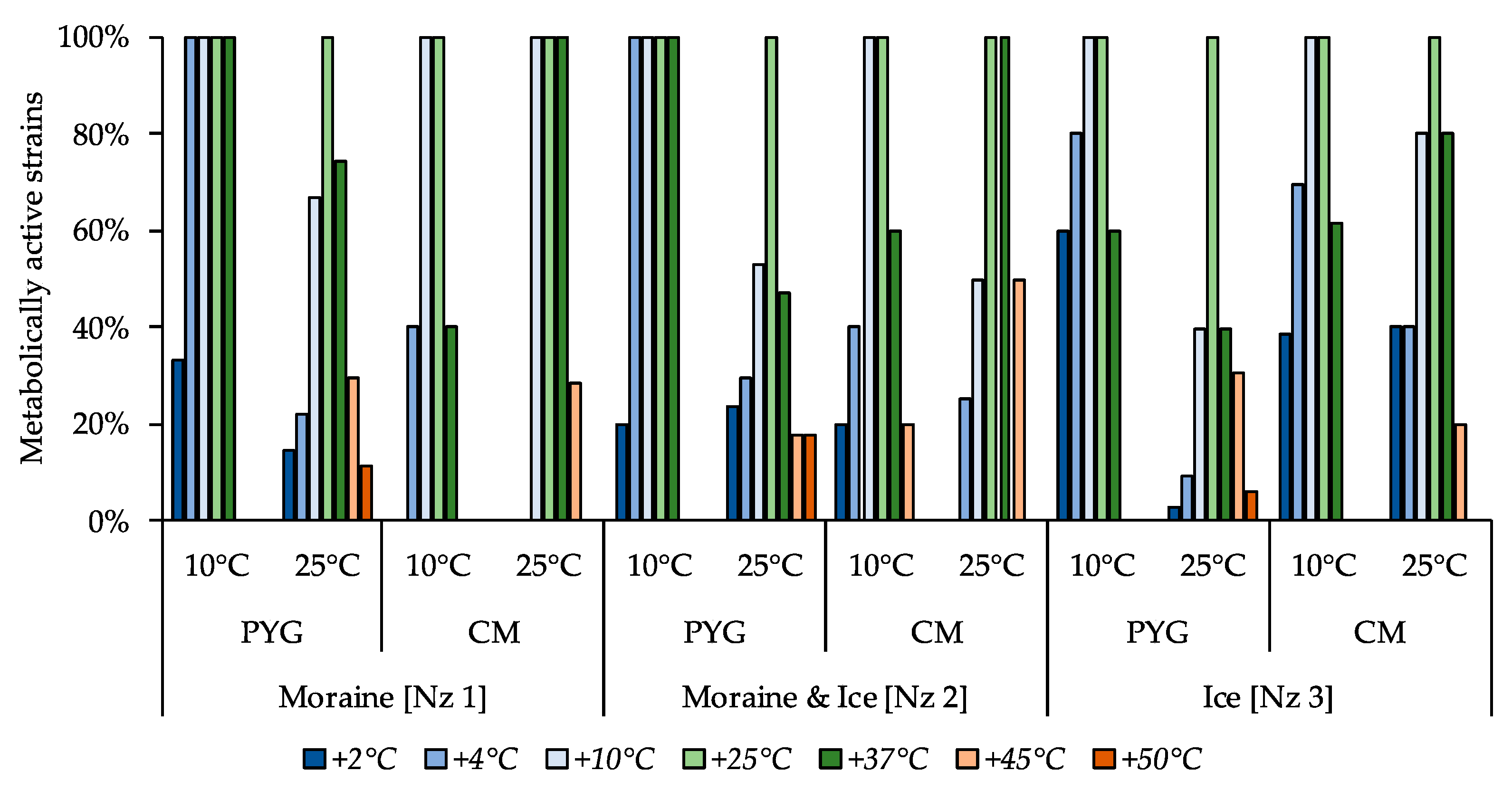
3.4. Metabolically Active Communities’ Structure
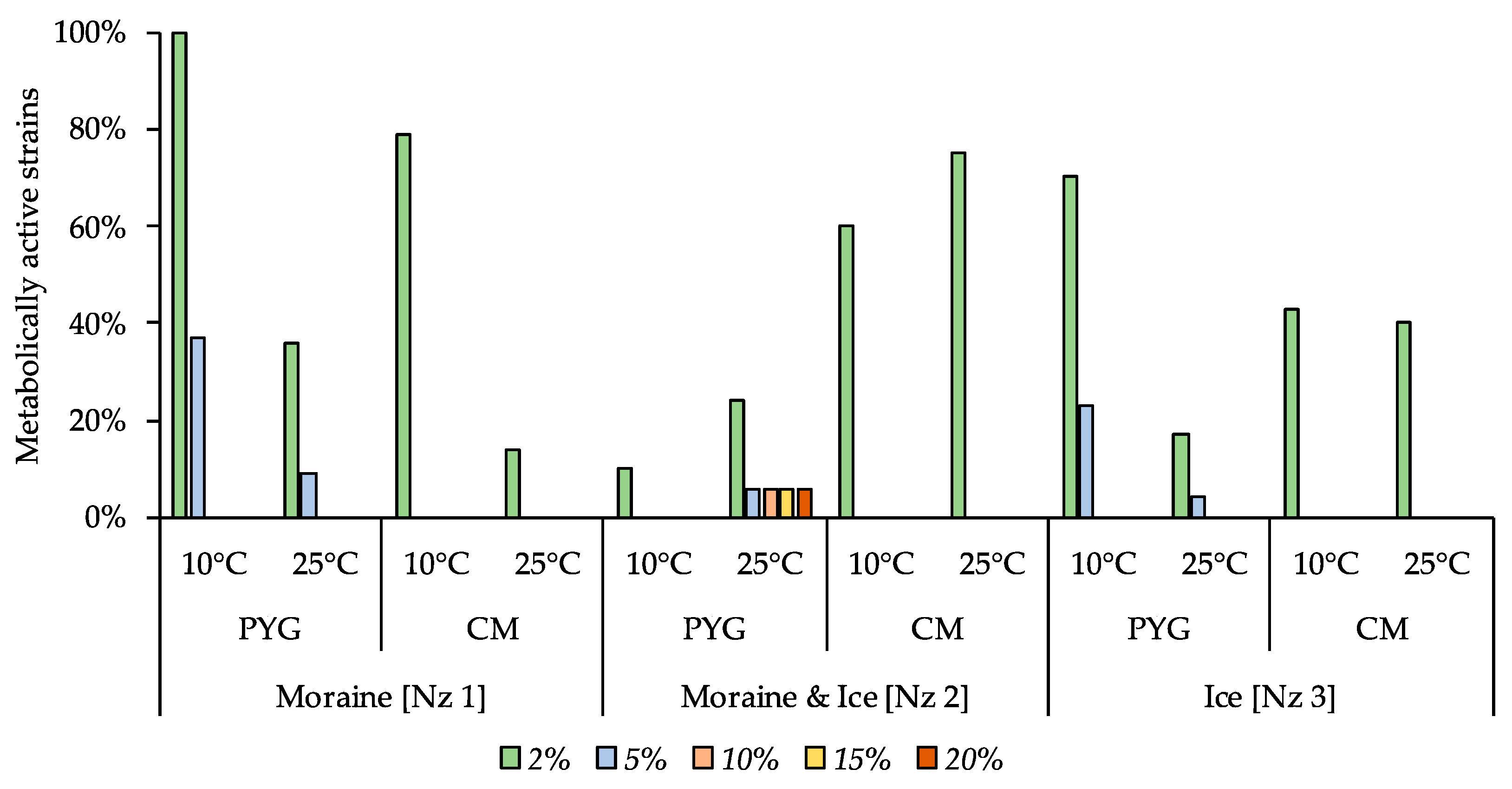
3.5. Multisubstrate Testing
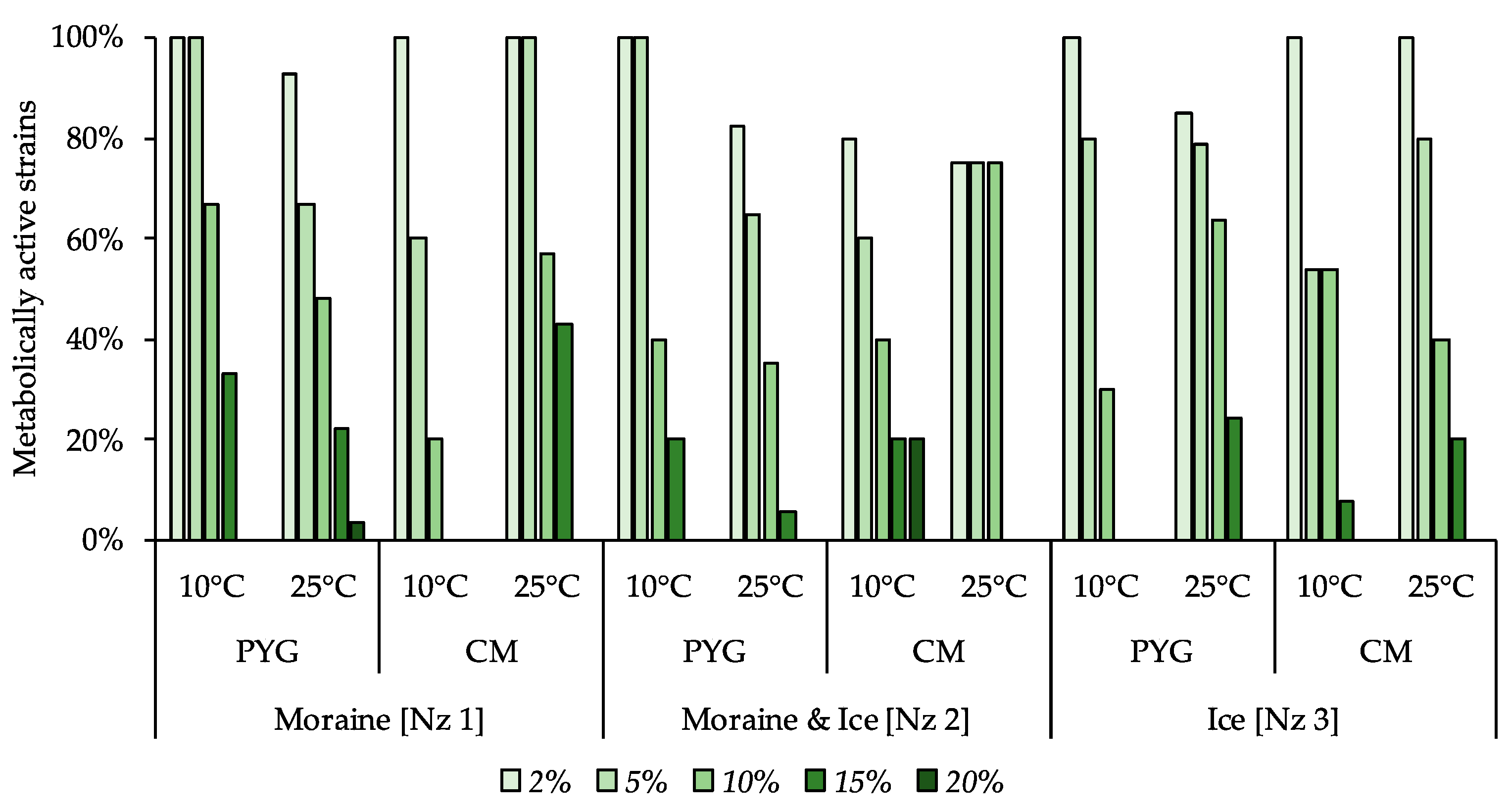
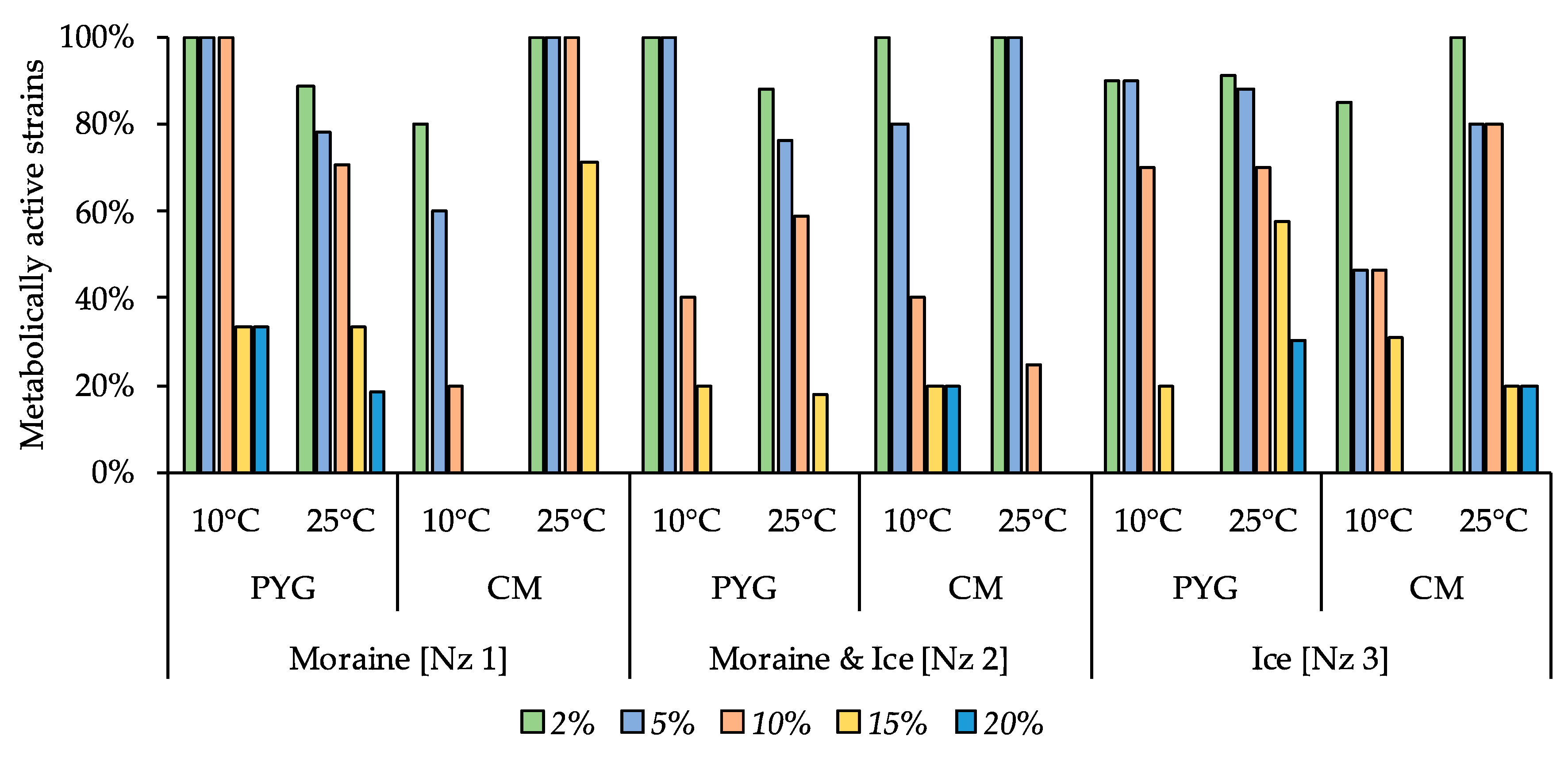
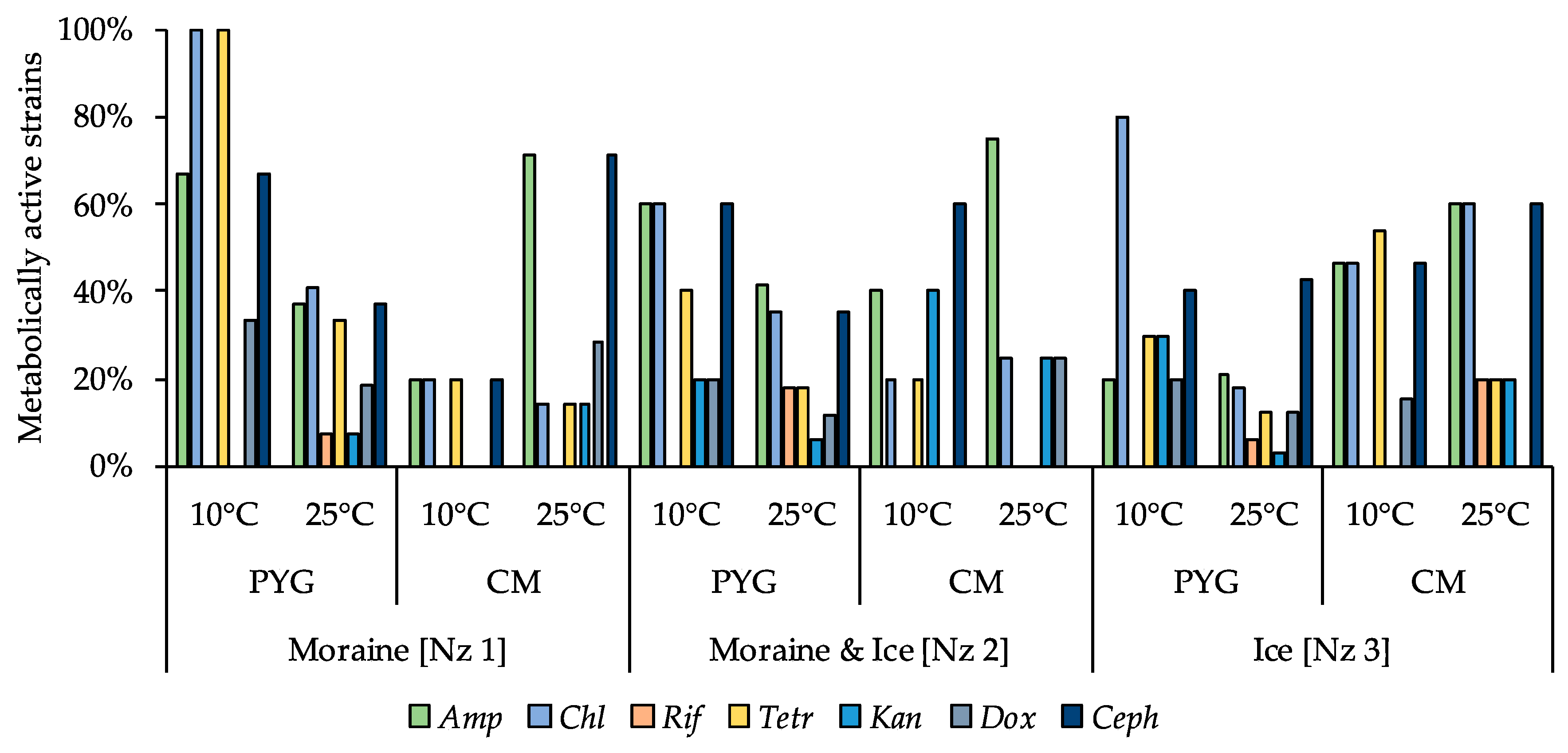
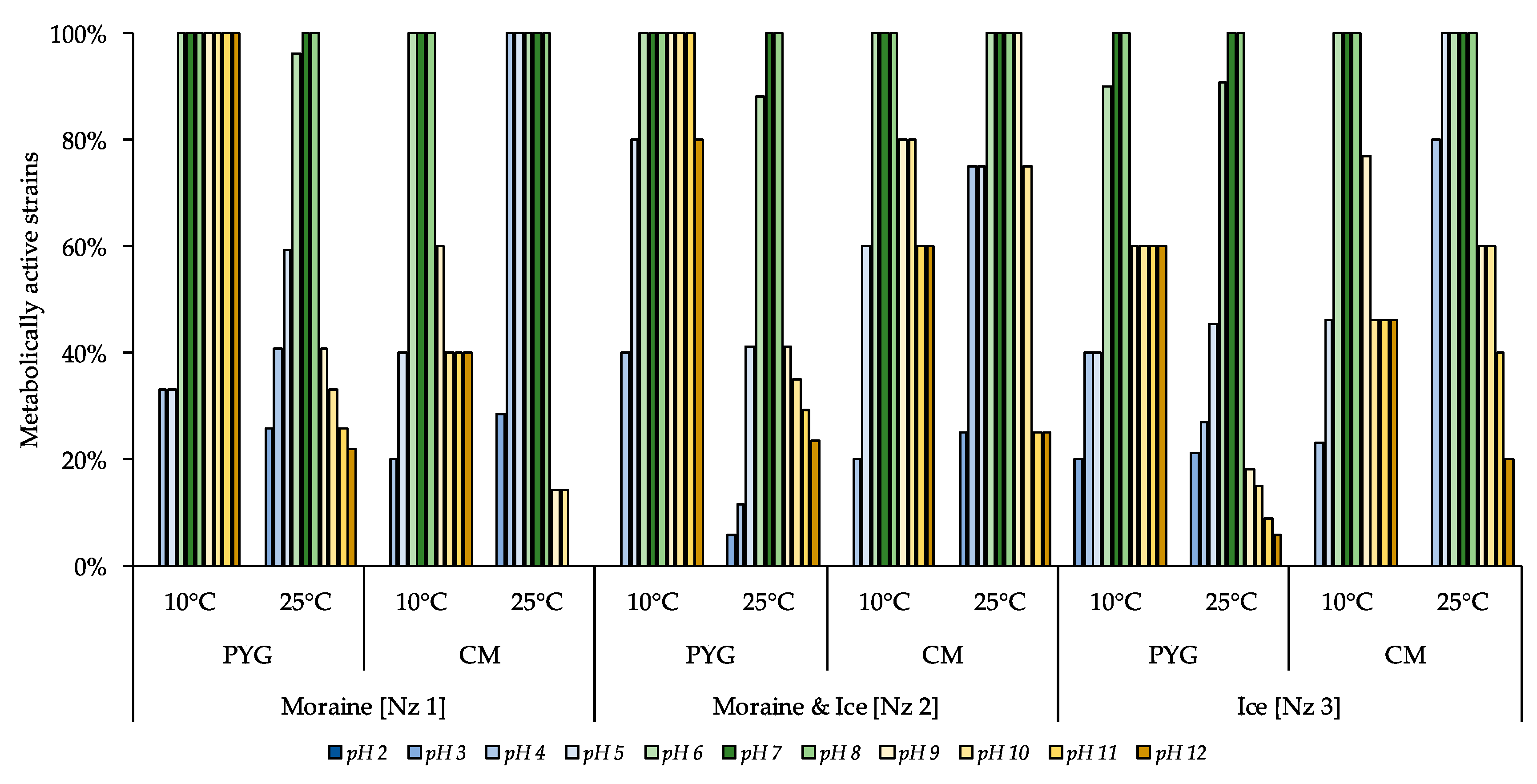
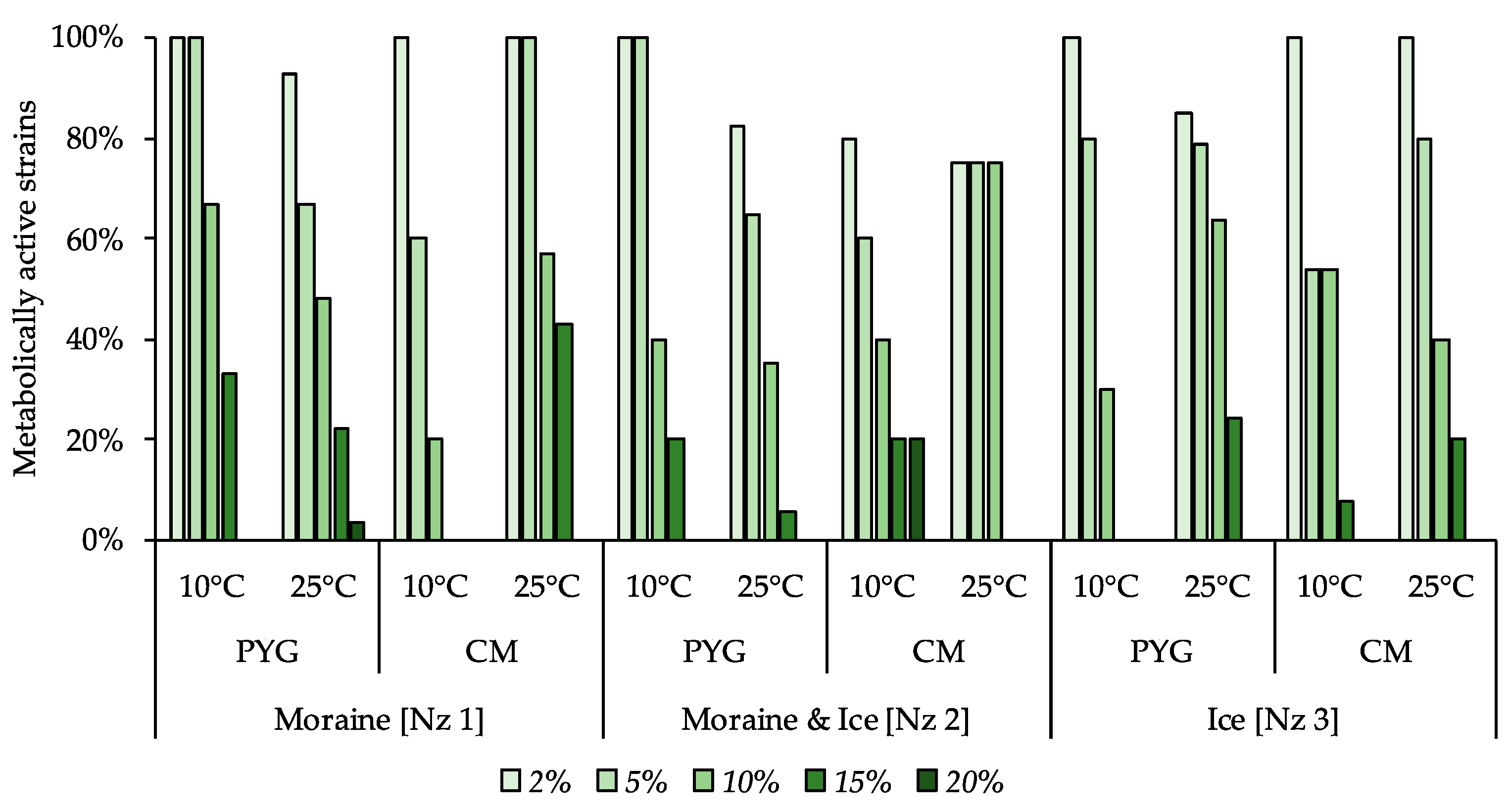
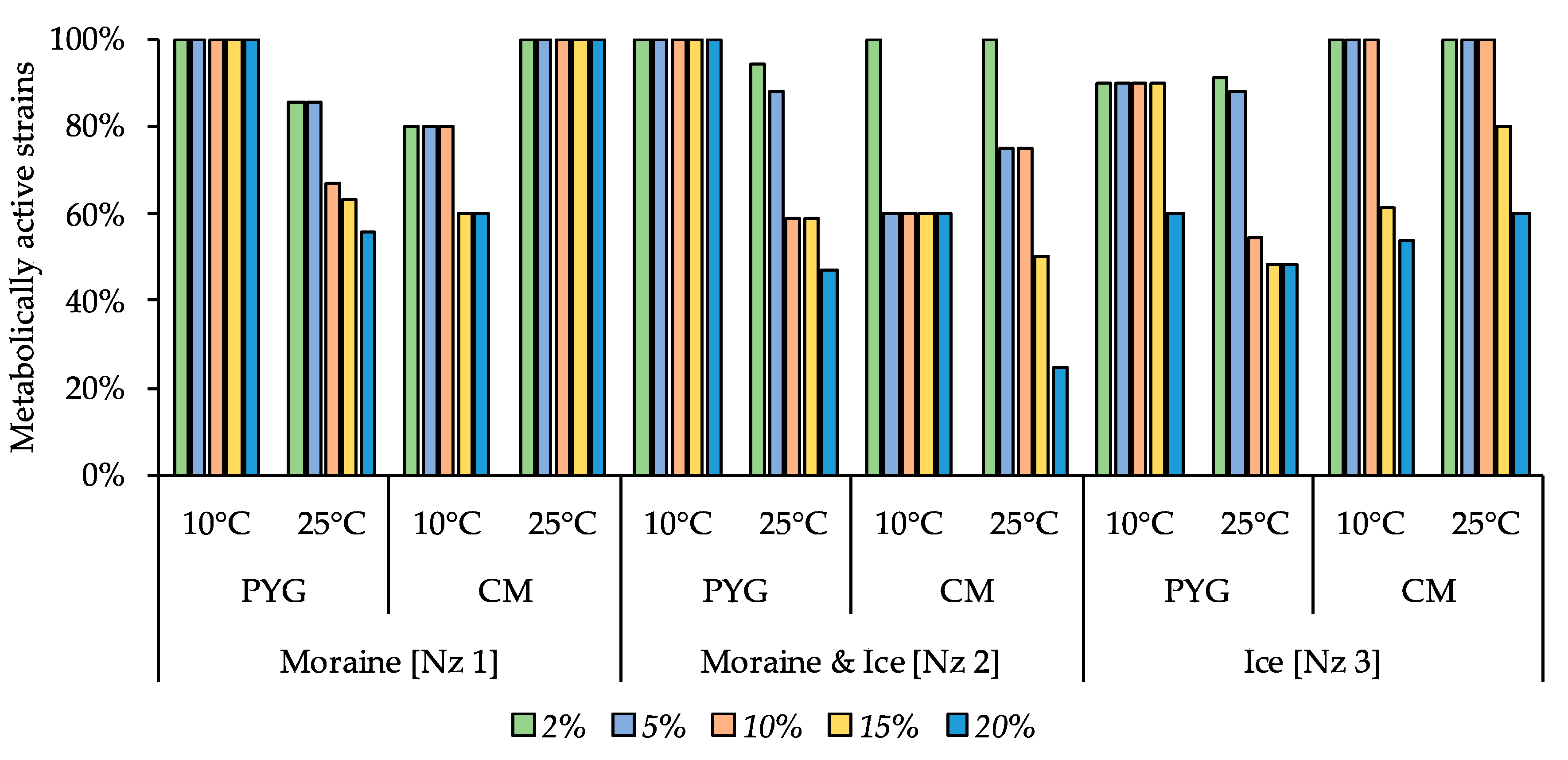
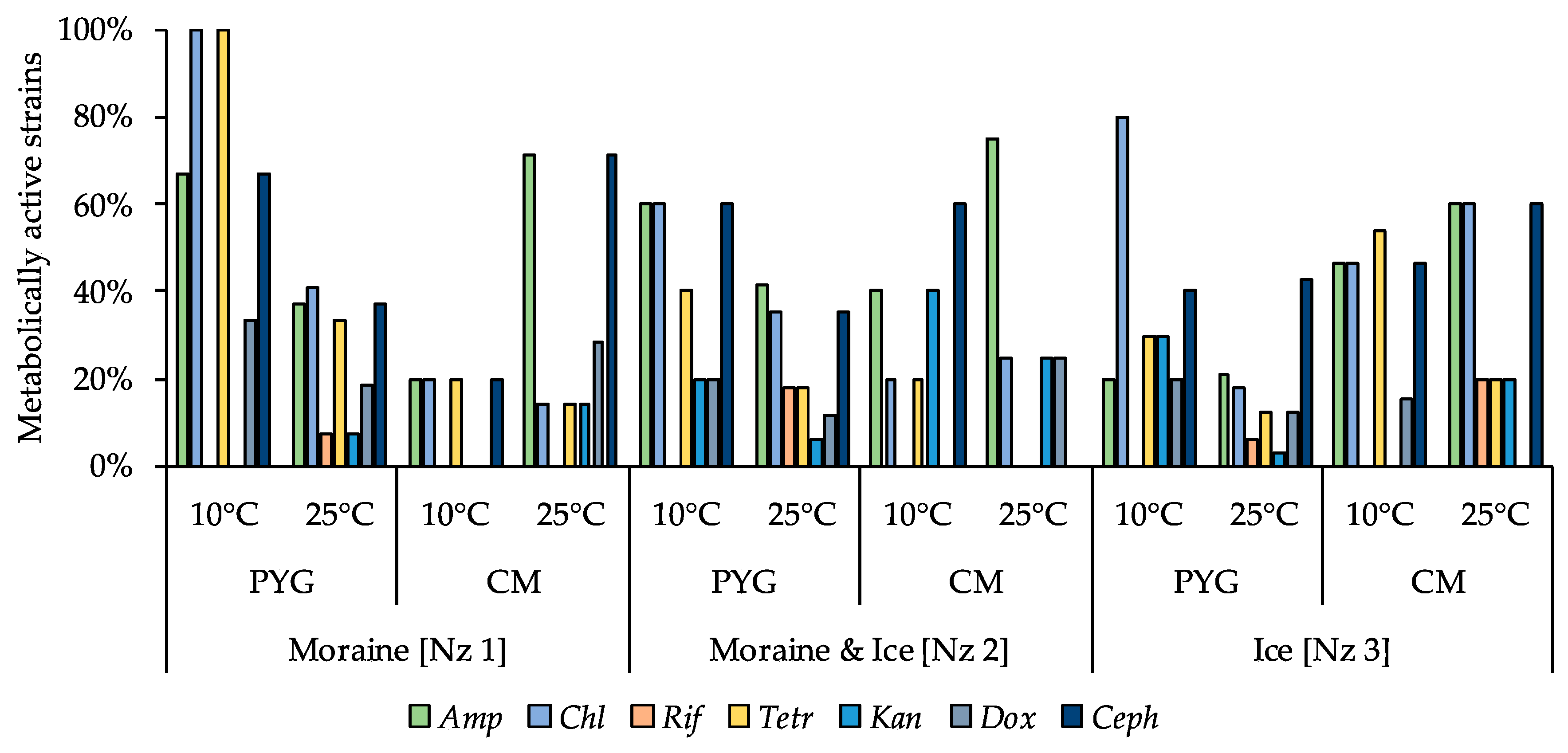
3.6. Physiological Assays of Isolated Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
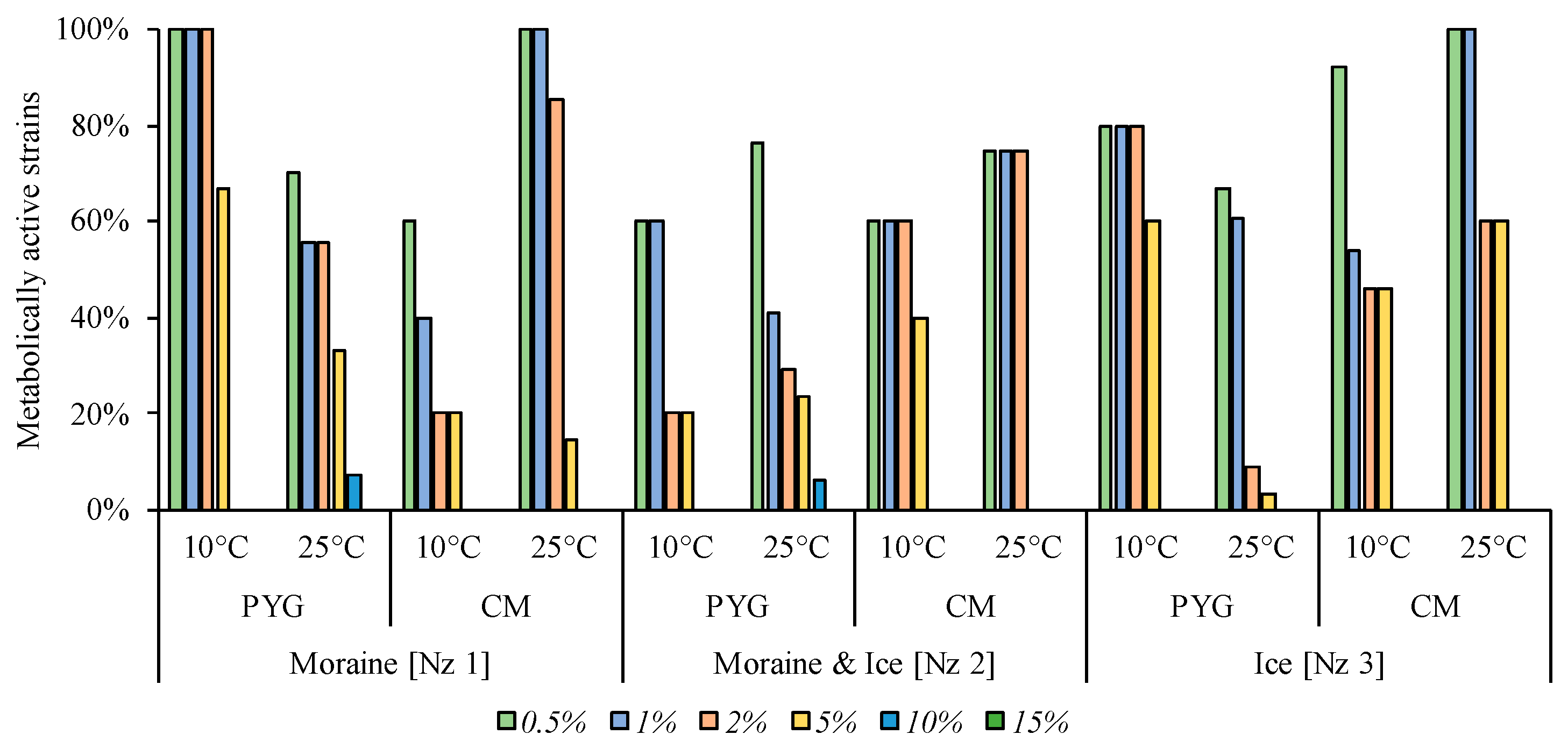{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Media | Temperature of Culturing | Cells/g or CFU/g |
|---|---|---|---|
| Moraine [Nz 1] | PYG | 10 °C | (1.60 ± 0.35) × 104 |
| 25 °C | (3.90 ± 0.15) × 105 | ||
| CM | 10 °C | (1.40 ± 0.17) × 106 | |
| 25 °C | (3.90 ± 0.14) × 106 | ||
| EFM | (3.51± 0.44)×108 | ||
| Moraine & Ice [Nz 2] | PYG | 10 °C | (1.77 ± 0.85) × 107 |
| 25 °C | (7.81 ± 0.99) × 107 | ||
| CM | 10 °C | (4.00 ± 0.16) × 105 | |
| 25 °C | (4.27 ± 0.39)×106 | ||
| EFM | (2.25 ± 0.36) × 108 | ||
| Ice [Nz 3] | PYG | 10 °C | (7.33 ± 0.21) × 105 |
| 25 °C | (3.90 ± 0.15) × 105 | ||
| CM | 10 °C | (3.35 ± 0.17) × 106 | |
| 25 °C | (3.28 ± 0.68) × 106 | ||
| EFM | (1.85 ± 0.46) × 108 | ||
Appendix B
| Taxonomic Affiliation | Moraine [Nz 1] | Moraine & Ice [Nz 2] | Ice [Nz 3] | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PYG | CM | PYG | CM | PYG | CM | |||||||
| 10 °C | 25 °C | 10 °C | 25 °C | 10 °C | 25 °C | 10 °C | 25 °C | 10 °C | 25 °C | 10 °C | 25 °C | |
| Acinetobacter pittii | - | - | - | - | - | - | - | - | 11% | - | - | - |
| Agrococcus jenensis | - | - | - | - | - | - | - | - | - | 2% | - | - |
| Arthrobacter agilis | - | - | - | - | - | 4% | - | - | - | - | - | - |
| Arthrobacter crystallopoietes | - | - | - | 5% | - | - | - | - | - | - | - | - |
| Arthrobacter ginsengisoli | - | 5% | - | - | - | 24% | - | 10% | - | - | 12% | - |
| Arthrobacter spp. | - | 2% | - | - | - | - | 14% | - | - | - | - | - |
| Arthrobacter sulfonivorans | - | - | - | - | - | - | - | - | 4% | - | - | - |
| Bacillus infantis | - | - | - | - | - | - | - | - | - | 1% | - | - |
| Bacillus megaterium | - | - | - | - | - | - | - | - | - | - | - | 21% |
| Bacillus pumilus | - | - | - | - | - | 4% | - | - | - | - | - | - |
| Bacillus simplex | - | 2% | - | - | - | - | - | - | 31% | - | - | - |
| Bacillus spp. | 34% | 20% | - | 12% | - | 8% | - | - | 6% | 5% | - | 12% |
| Bacillus subtilis | - | - | - | - | - | - | - | - | - | 1% | - | - |
| Brachybacterium spp. | - | 2% | - | - | - | - | - | - | - | - | - | - |
| Brevibacillus brevis | - | 9% | - | - | - | - | - | - | - | - | - | - |
| Brevibacterium aureum | - | 2% | - | - | - | - | - | - | - | - | - | - |
| Brevibacterium epidermidis | - | - | - | - | - | - | - | 40% | - | - | - | - |
| Brevibacterium linens | - | - | - | - | - | - | - | - | - | 2% | - | - |
| Brevibacterium luteolum | - | 2% | - | - | - | - | - | - | - | - | - | - |
| Brevibacterium spp. | - | - | - | - | - | - | 24% | - | - | 3% | - | - |
| Chryseobacterium kwangjuense | - | - | - | - | - | - | 19% | - | - | - | - | - |
| Corynebacterium mucifaciens | - | - | - | - | - | - | - | - | - | 4% | - | - |
| Dietzia cinnamea | - | 9% | - | - | - | - | - | - | - | 8% | - | - |
| Enterobacter spp. | - | 2% | - | - | - | - | - | - | - | - | - | - |
| Glutamicibacter spp. | - | - | - | 30% | - | - | - | - | - | - | - | - |
| Klebsiella spp. | - | - | - | - | - | - | - | - | 7% | - | - | - |
| Kocuria rosea | - | - | - | - | - | 4% | - | - | 10% | - | - | - |
| Leucobacter aridicollis | - | - | - | - | - | 4% | - | - | - | 3% | - | - |
| Leucobacter spp. | - | - | - | - | - | - | 28% | - | - | - | - | - |
| Methylobacterium spp. | - | 3% | - | - | - | - | - | - | - | - | - | - |
| Microbacterium flavescens | - | - | - | - | - | - | - | - | 9% | - | - | - |
| Microbacterium oxydans | - | 9% | 17% | - | - | - | - | - | 9% | - | - | - |
| Microbacterium paraoxydans | - | - | - | - | - | - | - | - | - | - | 7% | 27% |
| Microbacterium pumilum | - | 5% | - | - | - | - | - | - | - | - | - | - |
| Microbacterium spp. | - | - | - | - | 27% | - | - | - | 5% | 3% | - | - |
| Micrococcus luteus | - | 2% | - | - | - | - | - | - | - | - | - | - |
| Micrococcus spp. | 26% | 16% | - | - | - | 8% | - | - | - | 15% | - | - |
| Micrococcus terreus | - | - | - | - | - | - | - | - | - | 11% | - | - |
| Ochrobactrum spp. | - | - | - | - | - | - | - | - | - | 2% | - | - |
| Ochrobactrum thiophenivorans | - | - | - | - | - | 4% | - | - | - | - | 24% | - |
| Paracoccus marcusii | - | - | - | - | - | - | - | - | - | 4% | - | - |
| Paracoccus spp. | - | - | - | - | - | - | - | - | - | - | 8% | - |
| Planomicrobium okeanokoites | - | - | - | - | - | - | - | - | - | 2% | - | - |
| Planomicrobium spp. | - | - | - | - | - | - | - | - | - | 4% | - | - |
| Pseudarthrobacter scleromae | - | - | - | - | - | - | - | 40% | - | - | - | - |
| Pseudarthrobacter siccitolerans | - | - | 11% | - | - | - | - | - | - | - | - | - |
| Pseudarthrobacter spp. | - | - | - | - | - | 8% | - | - | - | - | - | - |
| Pseudochrobactrum spp. | - | - | - | - | - | 16% | - | - | - | - | - | - |
| Pseudomonas frederiksbergensis | - | 4% | - | 6% | - | - | - | - | - | - | - | - |
| Pseudomonas oryzihabitans | - | - | 47% | - | - | - | - | - | - | - | - | - |
| Pseudomonas putida | - | - | - | - | 15% | - | - | - | - | - | - | - |
| Pseudomonas spp. | - | 6% | - | - | - | 12% | 15% | 10% | - | 5% | - | - |
| Rhodococcus spp. | 40% | - | - | - | - | - | - | - | 8% | 2% | - | 28% |
| Sphingomonas spp. | - | - | - | - | - | - | - | - | - | 4% | - | - |
| Staphylococcus epidermidis | - | - | - | - | - | - | - | - | - | 8% | - | - |
| Stenotrophomonas maltophilia | - | - | - | - | 43% | - | - | - | - | 2% | 39% | 12% |
| Stenotrophomonas rhizophila | - | - | - | - | 15% | - | - | - | - | - | - | - |
| Stenotrophomonas spp. | - | - | - | - | - | 4% | - | - | - | - | 5% | - |
| Streptomyces spp. | - | - | 25% | 47% | - | - | - | - | - | 5% | 5% | - |
| Tsukamurella tyrosinosolvens | - | - | - | - | - | - | - | - | - | 4% | - | - |
References
- Makhalanyane, T.P.; Valverde, A.; Gunnigle, E.; Frossard, A.; Ramond, J.B.; Cowan, D.A. Microbial ecology of hot desert edaphic systems. FEMS Microbiol. Rev. 2015, 39, 203–221. [Google Scholar] [CrossRef] [PubMed]
- Jansson, J.K.; Taş, N. The microbial ecology of permafrost. Nat. Rev. Microbiol. 2014, 12, 414. [Google Scholar] [CrossRef] [PubMed]
- Mikucki, J.A.; Han, S.K.; Lanoil, B.D. Ecology of psychrophiles: Subglacial and permafrost environments. In Extremophiles Handbook; Horikoshi, K., Antranikian, G., Bull, A.T., Robb, F.T., Stetter, K.O., Eds.; Springer: Berlin, Germany, 2011; pp. 755–775. ISBN 978-4-431-53899-8. [Google Scholar]
- Goodfellow, M.; Nouioui, I.; Sanderson, R.; Xie, F.; Bull, A.T. Rare taxa and dark microbial matter: Novel bioactive actinobacteria abound in Atacama Desert soils. Anton. Leeuw. Int. J. G. 2018, 111, 1315–1332. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, M.; Golyshina, O.; Beloqui, A.; Golyshin, P.N. Mining enzymes from extreme environments. Curr. Opin. Microbiol. 2007, 10, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Belov, A.A.; Cheptsov, V.S.; Vorobyova, E.A.; Manucharova, N.A.; Ezhelev, Z.S. Stress-Tolerance and Taxonomy of Culturable Bacterial Communities Isolated from a Central Mojave Desert Soil Sample. Geosciences 2019, 9, 166. [Google Scholar] [CrossRef] [Green Version]
- Musilova, M.; Wright, G.; Ward, J.M.; Dartnell, L.R. Isolation of radiation-resistant bacteria from Mars analog Antarctic Dry Valleys by preselection, and the correlation between radiation and desiccation resistance. Astrobiology 2015, 15, 1076–1090. [Google Scholar] [CrossRef] [Green Version]
- Chyba, C.F.; Hand, K.P. Astrobiology: The study of the living universe. Annu. Rev. Astron. Astrophys. 2005, 43, 31–74. [Google Scholar] [CrossRef] [Green Version]
- Rampelotto, P. Extremophiles and extreme environments. Life 2013, 3, 482–485. [Google Scholar] [CrossRef]
- Gilichinsky, D.; Vishnivetskaya, T.; Petrova, M.; Spirina, E.; Mamykin, V.; Rivkina, E. Bacteria in permafrost. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Schinner, F., Marx, J.C., Eds.; Springer: Berlin, Heidelberg, 2008; ISBN 978-3-319-57056-3. [Google Scholar]
- Lindgren, A.R.; Buckley, B.A.; Eppley, S.M.; Reysenbach, A.L.; Stedman, K.M.; Wagner, J.T. Life on the edge—the biology of organisms inhabiting extreme environments: An introduction to the symposium. Integr. Comp. Biol. 2016, 56, 493–499. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.J.; Tow, L.A.; Stafford, W.; Cary, C.; Cowan, D.A. Bacterial diversity in three different Antarctic cold desert mineral soils. Microb. Ecol. 2006, 51, 413–421. [Google Scholar] [CrossRef]
- Yadav, A.N.; Sachan, S.G.; Verma, P.; Tyagi, S.P.; Kaushik, R.; Saxena, A.K. Culturable diversity and functional annotation of psychrotrophic bacteria from cold desert of Leh Ladakh (India). World J. Microb. Biot. 2015, 31, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.; Van Nostrand, J.D.; Zhou, J.; Pointing, S.B.; Farrell, R.L. Functional ecology of an Antarctic dry valley. Proc. Natl. Acad. Sci. USA 2013, 110, 8990–8995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belov, A.A.; Cheptsov, V.S.; Vorobyova, E.A. Soil bacterial communities of Sahara and Gibson deserts: Physiological and taxonomical characteristics. AIMS Microbiol. 2018, 4, 685–710. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. Molecular ecology of extremely halophilic Archaea and Bacteria. FEMS Microbiol. Ecol. 2002, 39, 1–7. [Google Scholar] [CrossRef]
- Aanderud, Z.T.; Vert, J.C.; Lennon, J.T.; Magnusson, T.W.; Breakwell, D.P.; Harker, A.R. Bacterial dormancy is more prevalent in freshwater than hypersaline lakes. Front. Microbiol. 2016, 7, 853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, D.B. Biodiversity and ecology of acidophilic microorganisms. FEMS Microbiol. Ecol. 1998, 27, 307–317. [Google Scholar] [CrossRef]
- Zhang, G.; Ma, X.; Niu, F.; Dong, M.; Feng, H.; An, L.; Cheng, G. Diversity and distribution of alkaliphilic psychrotolerant bacteria in the Qinghai–Tibet Plateau permafrost region. Extremophiles 2007, 11, 415–424. [Google Scholar] [CrossRef]
- Miteva, V. Bacteria in snow and glacier ice. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R.., Schinner, F., Marx, J.C., Eds.; Springer: Berlin, Germany, 2008; pp. 31–50. ISBN 978-3-319-57057-0. [Google Scholar]
- Anesio, A.M.; Laybourn-Parry, J. Glaciers and ice sheets as a biome. Trends. Ecol. Evol. 2012, 27, 219–225. [Google Scholar] [CrossRef]
- Gaboyer, F.; Le Milbeau, C.; Bohmeier, M.; Schwendner, P.; Vannier, P.; Beblo-Vranesevic, K.; Rabbow, E.; Foucher, F.; Gautret, P.; Guégan, R.; et al. Mineralization and preservation of an extremotolerant bacterium isolated from an early Mars analog environment. Sci. Rep. 2017, 7, 8775. [Google Scholar] [CrossRef] [Green Version]
- Takai, K. Limits of Terrestrial Life and Biosphere. In Astrobiology: From the Origins of Life to the Search for Extraterrestrial Intelligence; Yamagishi, A., Kakegawa, T., Usui, T., Eds.; Springer: Singapore, 2019; pp. 323–344. ISBN 978-981-13-3639-3. [Google Scholar]
- Tse, C.; Ma, K. Growth and Metabolism of Extremophilic Microorganisms. In Biotechnology of Extremophiles; Rampelotto, P.H., Ed.; Springer: Cham, Germany, 2016; pp. 1–46. ISBN 978-3-319-13521-2. [Google Scholar]
- Gilichinsky, D.A.; Wagener, S.; Vishnevetskaya, T.A. Permafrost microbiology. Permafrost. Periglac. 1995, 6, 281–291. [Google Scholar] [CrossRef]
- Margesin, R.; Collins, T. Microbial ecology of the cryosphere (glacial and permafrost habitats): Current knowledge. Appl. Microbiol. Biotechnol. 2019, 103, 2537–2549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorobyova, E.; Soina, V.; Gorlenko, M.; Minkovskaya, N.; Zalinova, N.; Mamukelashvili, A.; Gilichinsky, D.; Rivkina, E.; Vishnivetskaya, T. The deep cold biosphere: Facts and hypothesis. FEMS Microbiol. Rev. 1997, 20, 277–290. [Google Scholar] [CrossRef]
- Gilichinsky, D.A.; Wilson, G.S.; Friedmann, E.I.; McKay, C.P.; Sletten, R.S.; Rivkina, E.M.; Vishnivetskaya, T.A.; Erokhina, L.G.; Ivanushkina, N.E.; Kochkina, G.A.; et al. Microbial populations in Antarctic permafrost: Biodiversity, state, age, and implication for astrobiology. Astrobiology 2007, 7, 275–311. [Google Scholar] [CrossRef] [PubMed]
- Dartnell, L.R.; Hunter, S.J.; Lovell, K.V.; Coates, A.J.; Ward, J.M. Low-temperature ionizing radiation resistance of Deinococcus radiodurans and Antarctic Dry Valley bacteria. Astrobiology 2010, 10, 717–732. [Google Scholar] [CrossRef] [Green Version]
- Vishnivetskaya, T.; Kathariou, S.; McGrath, J.; Gilichinsky, D.; Tiedje, J.M. Low-temperature recovery strategies for the isolation of bacteria from ancient permafrost sediments. Extremophiles 2000, 4, 165–173. [Google Scholar] [CrossRef]
- Rivkina, E.M.; Laurinavichus, K.S.; Gilichinsky, D.A.; Shcherbakova, V.A. Methane generation in permafrost sediments. In Doklady Biological Sciences; MAIK Nauka/Interperiodica: Moscow, Russia, 2002; Volume 383, pp. 179–181. ISSN 1608-3105. [Google Scholar] [CrossRef]
- Rivkina, E.M.; Friedmann, E.I.; McKay, C.P.; Gilichinsky, D.A. Metabolic activity of permafrost bacteria below the freezing point. Appl. Environ. Microbiol. 2000, 66, 3230–3233. [Google Scholar] [CrossRef] [Green Version]
- Mykytczuk, N.C.; Foote, S.J.; Omelon, C.R.; Southam, G.; Greer, C.W.; Whyte, L.G. Bacterial growth at −15 °C; molecular insights from the permafrost bacterium Planococcus halocryophilus Or1. ISME J. 2013, 7, 1211–1226. [Google Scholar] [CrossRef] [Green Version]
- Panikov, N.S.; Sizova, M.V. Growth kinetics of microorganisms isolated from Alaskan soil and permafrost in solid media frozen down to− 35 C. FEMS Microbiol. Ecol. 2007, 59, 500–512. [Google Scholar] [CrossRef]
- Antony, R.; Krishnan, K.P.; Laluraj, C.M.; Thamban, M.; Dhakephalkar, P.K.; Engineer, A.S.; Shivaji, S. Diversity and physiology of culturable bacteria associated with a coastal Antarctic ice core. Microbiol. Res. 2012, 167, 372–380. [Google Scholar] [CrossRef]
- Cheptsov, V.S.; Vorobyova, E.A.; Manucharova, N.A.; Gorlenko, M.V.; Pavlov, A.K.; Vdovina, M.A.; Lomasov, V.N.; Bulat, S.A. 100 kGy gamma-affected microbial communities within the ancient Arctic permafrost under simulated Martian conditions. Extremophiles 2017, 21, 1057–1067. [Google Scholar] [CrossRef] [PubMed]
- Vishniac, H.S. The microbiology of Antarctic soils. In Antarctic Microbiology; Friedmann, E.I., Thistle, A.B., Eds.; Wiley-Liss: New York, NY, USA, 1993; pp. 297–341. ISBN 0-471-50776-8. [Google Scholar]
- Soina, V.S.; Mulyukin, A.L.; Demkina, E.V.; Vorobyova, E.A.; El-Registan, G.I. The structure of resting bacterial populations in soil and subsoil permafrost. Astrobiology 2004, 4, 345–358. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, R.C.; Radtke, K.J.; Mykytczuk, N.C.; Greer, C.W.; Whyte, L.G. Life at the wedge: The activity and diversity of Arctic ice wedge microbial communities. Astrobiology 2012, 12, 347–360. [Google Scholar] [CrossRef] [PubMed]
- Yergeau, E.; Hogues, H.; Whyte, L.G.; Greer, C.W. The functional potential of high Arctic permafrost revealed by metagenomic sequencing, qPCR and microarray analyses. ISME J. 2010, 4, 1206. [Google Scholar] [CrossRef]
- Steven, B.; Briggs, G.; McKay, C.P.; Pollard, W.H.; Greer, C.W.; Whyte, L.G. Characterization of the microbial diversity in a permafrost sample from the Canadian high Arctic using culture-dependent and culture-independent methods. FEMS Microbiol. Ecol. 2007, 59, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Frank-Fahle, B.A.; Yergeau, E.; Greer, C.W.; Lantuit, H.; Wagner, D. Microbial functional potential and community composition in permafrost-affected soils of the NW Canadian Arctic. PLoS ONE 2014, 9, e84761. [Google Scholar] [CrossRef] [Green Version]
- Perron, G.G.; Whyte, L.; Turnbaugh, P.J.; Goordial, J.; Hanage, W.P.; Dantas, G.; Desai, M.M. Functional characterization of bacteria isolated from ancient arctic soil exposes diverse resistance mechanisms to modern antibiotics. PLoS ONE 2015, 10, e0069533. [Google Scholar] [CrossRef]
- Zhou, J.; Davey, M.E.; Figueras, J.B.; Rivkina, E.; Gilichinsky, D.; Tiedje, J.M. Phylogenetic diversity of a bacterial community determined from Siberian tundra soil DNA. Microbiology 1997, 143, 3913–3919. [Google Scholar] [CrossRef] [Green Version]
- Neufeld, J.D.; Mohn, W.W. Unexpectedly high bacterial diversity in arctic tundra relative to boreal forest soils, revealed by serial analysis of ribosomal sequence tags. Appl. Environ. Microbiol. 2005, 71, 5710–5718. [Google Scholar] [CrossRef] [Green Version]
- Vishnivetskaya, T.A.; Petrova, M.A.; Urbance, J.; Ponder, M.; Moyer, C.L.; Gilichinsky, D.A.; Tiedje, J.M. Bacterial community in ancient Siberian permafrost as characterized by culture and culture-independent methods. Astrobiology 2006, 6, 400–414. [Google Scholar] [CrossRef]
- Rivkina, E.; Petrovskaya, L.; Vishnivetskaya, T.; Krivushin, K.; Shmakova, L.; Tutukina, M.; Meyers, A.; Kondrashov, F. Metagenomic analyses of the late Pleistocene permafrost–additional tools for reconstruction of environmental conditions. Biogeosciences 2016, 13, 2207–2219. [Google Scholar] [CrossRef] [Green Version]
- Pradeep Kumar, P. Cryopeg. In Encyclopedia of Snow, Ice and Glaciers. Encyclopedia of Earth Sciences Series; Singh, V.P., Singh, P., Haritashya, U.K., Eds.; Springer: Dordrecht, The Netherlands, 2011; ISBN 978-90-481-2642-2_90. [Google Scholar]
- Bakermans, C.; Tsapin, A.I.; Souza-Egipsy, V.; Gilichinsky, D.A.; Nealson, K.H. Reproduction and metabolism at −10 C of bacteria isolated from Siberian permafrost. Environ. Microbiol. 2003, 5, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.C.; Brouchkov, A.; Griva, G.; Schinner, F.; Margesin, R. Isolation and characterization of bacteria from ancient Siberian permafrost sediment. Biology 2013, 2, 85–106. [Google Scholar] [CrossRef]
- Goryachkin, S.V. The Soils of the North (Structure, Genesis, Ecology, Evolution); Russian Academy of Science: Moscow, Russia, 2010; p. 414, [Book in Russian]; ISBN 978-5-89118-508-1. [Google Scholar]
- MSU-Lab Research Laboratory. Available online: https://www.msulab.ru/ (accessed on 27 September 2019).
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Amann, R.I.; Ludwig, W. Ribosomal RNA-Targeted Nucleic Acid Probes for Studies in Microbial Ecology. FEMS Microbiol. Rev. 2000, 24, 555–565. [Google Scholar] [CrossRef]
- Manucharova, N.A.; Vlasenko, A.N.; Men’ko, E.V.; Zvyagintsev, D.G. Specificity of the chitinolytic microbial complex of soils incubated at different temperatures. Microbiology 2011, 80, 205–215. [Google Scholar] [CrossRef]
- Cheptsov, V.S.; Vorobyova, E.A.; Osipov, G.A.; Manucharova, N.A.; Polyanskaya, L.M.; Gorlenko, M.V.; Pavlov, A.K.; Rosanova, M.S.; Lomasov, V.N. Microbial activity in Martian analog soils after ionizing radiation: Implications for the preservation of subsurface life on Mars. AIMS Microbiol. 2018, 4, 541–562. [Google Scholar] [CrossRef]
- Rampelotto, P.H. Resistance of microorganisms to extreme environmental conditions and its contribution to astrobiology. Sustainability 2010, 2, 1602–1623. [Google Scholar] [CrossRef] [Green Version]
- Depository of Live Systems. Available online: https://depo.msu.ru (accessed on 27 September 2019).
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Wade, W.G. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [CrossRef] [Green Version]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- DeLong, E.F. Archaea in coastal marine environments. Proc. Natl. Acad. Sci. USA 1992, 89, 5685–5689. [Google Scholar] [CrossRef] [Green Version]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 1991; pp. 115–175. [Google Scholar]
- Chromas and ChromasPro DNA Sequencing Software—Technelysium Pty Ltd. Available online: http: //www.technelysium.com.au (accessed on 27 September 2019).
- Clustal Omega <Multiple Sequence Alignment <EMBL-EBI. Available online: http://www.ebi.ac.uk/ Tools/msa/clustalo/ (accessed on 27 September 2019).
- BLAST: Basic Local Alignment Search Tool. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 27 September 2019).
- Mega Home. Available online: https://www.megasoftware.net/ (accessed on 27 September 2019).
- Cottrell, M.T.; Wood, D.N.; Yu, L.; Kirchman, D.L. Selected chitinase genes in cultured and uncultured marine bacteria in the α-and γ-subclasses of the proteobacteria. Appl. Environ. Microbiol. 2000, 66, 1195–1201. [Google Scholar] [CrossRef] [Green Version]
- Miteva, V.; Rinehold, K.; Sowers, T.; Sebastian, A.; Brenchley, J. Abundance, viability and diversity of the indigenous microbial populations at different depths of the NEEM Greenland ice core. Polar Res. 2015, 34, 25057. [Google Scholar] [CrossRef] [Green Version]
- Muller, O.; Bang-Andreasen, T.; White III, R.A.; Elberling, B.; Tas, N.; Kneafsey, T.; Jansson, J.K.; Ovreas, L. Disentangling the complexity of permafrost soil by using high resolution profiling of microbial community composition, key functions and respiration rates. Environ. Microbiol. 2018, 20, 4328–4342. [Google Scholar] [CrossRef] [Green Version]
- Behrendt, U.; Schumann, P.; Ulrich, A. Agrococcus versicolor sp. nov., an actinobacterium associated with the phyllosphere of potato plants. Int. J. Syst. Evol. Microbiol. 2008, 58, 2833–2838. [Google Scholar] [CrossRef]
- Manucharova, N.A.; Trosheva, E.V.; Kol’tsova, E.M.; Demkina, E.V.; Karaevskaya, E.V.; Rivkina, E.M.; Mardanov, A.V. Characterization of the structure of the prokaryotic complex of Antarctic permafrost by molecular genetic techniques. Microbiology 2016, 85, 102–108. [Google Scholar] [CrossRef]
- Bajerski, F.; Ganzert, L.; Mangelsdorf, K.; Padur, L.; Lipski, A.; Wagner, D. Chryseobacterium frigidisoli sp. nov., a psychrotolerant species of the family Flavobacteriaceae isolated from sandy permafrost from a glacier forefield. Int. J. Syst. Evol. Microbiol. 2013, 63, 2666–2671. [Google Scholar] [CrossRef]
- Ponder, M.; Vishnivetskaya, T.; McGrath, J.; Tiedje, J. Microbial life in permafrost: Extended times in extreme conditions. In Life in the Frozen State; Fuller, B.J., Lane, N., Benson, E.E., Eds.; CRC Press: Boca Raton, FL, USA, 2004; ISBN 0-203-67332-8. [Google Scholar] [CrossRef]
- Steven, B.; Pollard, W.H.; Greer, C.W.; Whyte, L.G. Microbial diversity and activity through a permafrost/ground ice core profile from the Canadian high Arctic. Environ. Microbiol. 2008, 10, 3388–3403. [Google Scholar] [CrossRef] [Green Version]
- Singh, P.; Singh, S.M.; Singh, R.N.; Naik, S.; Roy, U.; Srivastava, A.; Bolter, M. Bacterial communities in ancient permafrost profiles of Svalbard, Arctic. J. Basic Microbiol. 2017, 57, 1018–1036. [Google Scholar] [CrossRef]
- Balan, B.M.; Binish, S.; Sruthy, K.; Kottekkattu, P.; Mohan, M. Mercury tolerance and biosorption in bacteria isolated from Ny-Alesund, Svalbard, Arctic. J. Basic Microbiol. 2018, 58, 286–295. [Google Scholar] [CrossRef]
- Liu, Y.; Yao, T.; Zhu, L.; Jiao, N.; Liu, X.; Zeng, Y.; Jiang, H. Bacterial diversity of freshwater alpine lake Puma Yumco on the Tibetan Plateau. Geomicrobiol. J. 2009, 26, 131–145. [Google Scholar] [CrossRef]
- Raymond-Bouchard, I.; Goordial, J.; Zolotarov, Y.; Ronholm, J.; Stromvik, M.; Bakermans, C.; Whyte, L.G. Conserved genomic and amino acid traits of cold adaptation in subzero-growing Arctic permafrost bacteria. FEMS Microbiol. Ecol. 2018, 94, fiy023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steven, B.; Leveille, R.; Pollard, W.H.; Whyte, L.G. Microbial ecology and biodiversity in permafrost. Extremophiles 2006, 10, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Zhao, Q.; Zhang, G.; Jiang, Z.; Sheng, H.; Feng, H.; An, L. Paracoccus tibetensis sp. nov.; isolated from Qinghai-Tibet Plateau permafrost. Int. J. Syst. Evol. Microbiol. 2013, 63, 1902–1905. [Google Scholar] [CrossRef] [Green Version]
- Romanenko, L.A.; Tanaka, N.; Frolova, G.M.; Mikhailov, V.V. Pseudochrobactrum glaciei sp. nov.; isolated from sea ice collected from Peter the Great Bay of the Sea of Japan. Int. J. Syst. Evol. Microbiol. 2008, 58, 2454–2458. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.C.; Busse, H.J.; Liu, H.C.; Zhou, Y.G.; Schinner, F.; Margesin, R. Sphingomonas glacialis sp. nov.; a psychrophilic bacterium isolated from alpine glacier cryoconite. Int. J. Syst. Evol. Microbiol. 2011, 61, 587–591. [Google Scholar] [CrossRef] [Green Version]
- D’Amico, S.; Collins, T.; Marx, J.C.; Feller, G.; Gerday, C. Psychrophilic microorganisms: Challenges for life. EMBO Rep. 2006, 7, 385–389. [Google Scholar] [CrossRef]
- Yi Pan, S.; Tan, G.Y.; Convey, P.; Pearce, D.A.; Tan, I.K. Diversity and bioactivity of actinomycetes from Signy Island terrestrial soils, maritime Antarctic. Adv. Polar Sci. 2013, 24, 208–212. [Google Scholar] [CrossRef]
- Gunde-Cimerman, N.; Sonjak, S.; Zalar, P.; Frisvad, J.C.; Diderichsen, B.; Plemenitas, A. Extremophilic fungi in arctic ice: A relationship between adaptation to low temperature and water activity. Phys. Chem. Earth. 2003, 28, 1273–1278. [Google Scholar] [CrossRef]
- Rivkina, E.; Laurinavichius, K.; McGrath, J.; Tiedje, J.; Shcherbakova, V.; Gilichinsky, D. Microbial life in permafrost. Adv. Space Res. 2004, 33, 1215–1221. [Google Scholar] [CrossRef]
- Pedros-Alio, C. The rare bacterial biosphere. Annu. Rev. Mar. Sci. 2012, 4, 449–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mindlin, S.Z.; Soina, V.S.; Petrova, M.A.; Gorlenko, Z.M. Isolation of antibiotic resistance bacterial strains from Eastern Siberia permafrost sediments. Russ. J. Genet. 2008, 44, 27–34. [Google Scholar] [CrossRef]
- Bjedov, I.; Tenaillon, O.; Gerard, B.; Souza, V.; Denamur, E.; Radman, M.; Taddei, F.; Matic, I. Stress-induced mutagenesis in bacteria. Science 2003, 300, 1404–1409. [Google Scholar] [CrossRef] [PubMed]

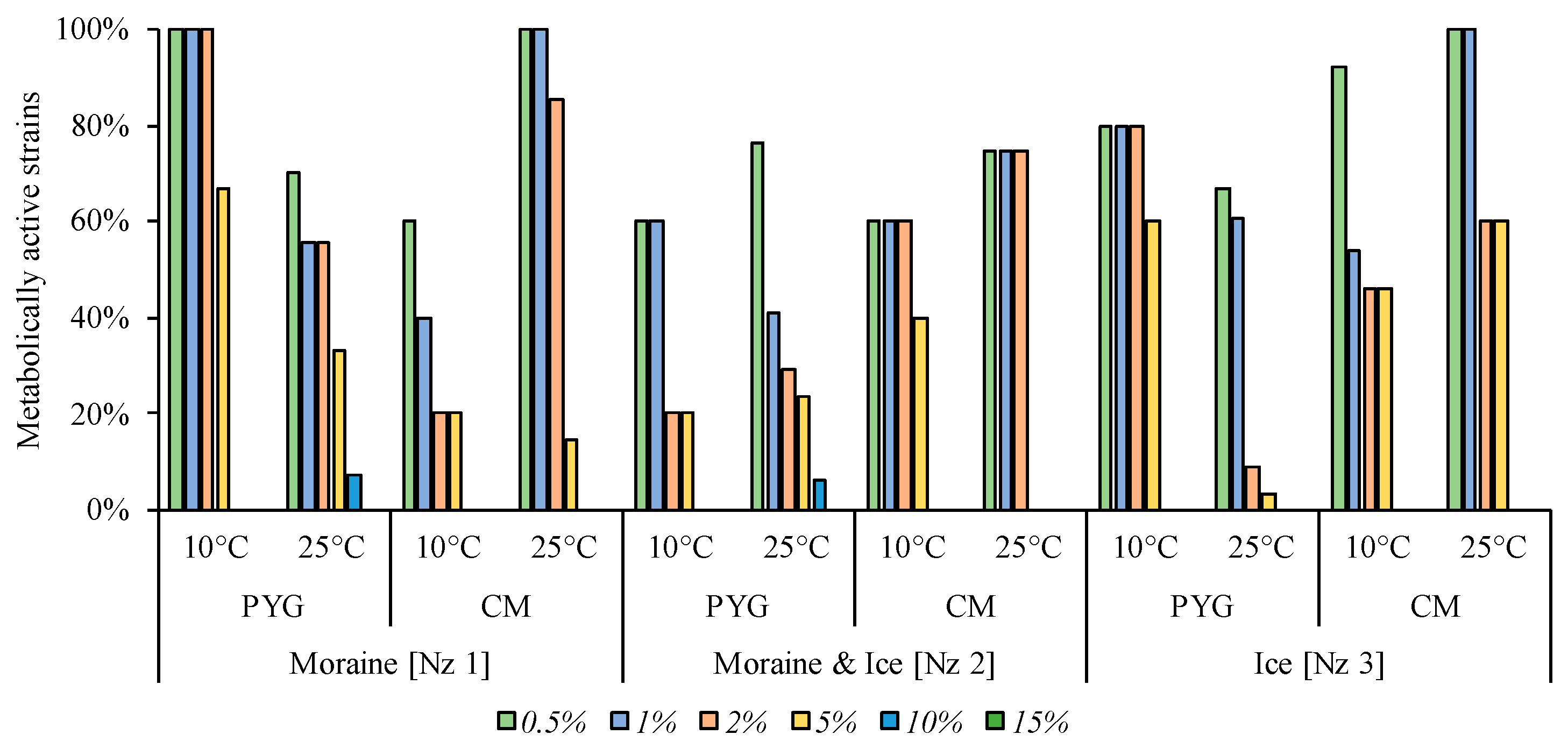
| Element | ppm | Element | ppm | Anions | mg/L |
|---|---|---|---|---|---|
| Ag | 0.0003 | Mg | 1.96 | F– | 0.3182 |
| Al | 0.1667 | Mn | 0.0182 | Cl– | 6.405 |
| As | 0.0124 | Mo | 0.0065 | NO2– | 0.3067 |
| Ba | 0.0075 | Na | 8.38 | NO3– | 43.1043 |
| Ca | 5.83 | Ni | 0.0231 | SO42– | 2.6359 |
| Cd | 0.0003 | P | 0.456 | PO43– | not detected |
| Co | 0.0004 | S | 12.29 | ||
| Cr | 0.0011 | Si | 0.9007 | ||
| Cu | 0.0174 | Sr | 0.0437 | ||
| Fe | 0.189 | V | 0.0027 | ||
| K | 8.34 | Zn | 0.0103 | ||
| Li | 0.0011 |
| Sample | Number of Isolated Strains | |||
|---|---|---|---|---|
| PYG | CM | |||
| 10 °C | 25 °C | 10 °C | 25 °C | |
| Moraine [Nz 1] | 3 | 26 | 5 | 7 |
| Moraine & Ice [Nz 2] | 5 | 17 | 5 | 4 |
| Ice [Nz 3] | 10 | 32 | 7 | 13 |
| Incubation Temperature | Community Indices | Substrates Assimilation, Relative Units | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| The Number of Assimilated substrates, N | Shannon Index, H | Evenness, E | Specific Metabolic Work, W | Pentoses | Hexoses | Oligosaccharides | Alcohols | Amino Acids | Salts of Organic Acids | Polymers | Amines Amidesnucleosides | |
| 10 °C | 17 | 3.64 | 0.91 | 0.45 | 0.27 | 0.11 | 0.6 | bdl | 4.2 | 0.56 | 1.73 | 0.24 |
| 25 °C | 7 | 2.33 | 0.83 | 0.25 | bdl | bdl | 0.08 | 0.09 | 1.55 | bdl | bdl | bdl |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belov, A.A.; Cheptsov, V.S.; Manucharova, N.A.; Ezhelev, Z.S. Bacterial Communities of Novaya Zemlya Archipelago Ice and Permafrost. Geosciences 2020, 10, 67. https://doi.org/10.3390/geosciences10020067
Belov AA, Cheptsov VS, Manucharova NA, Ezhelev ZS. Bacterial Communities of Novaya Zemlya Archipelago Ice and Permafrost. Geosciences. 2020; 10(2):67. https://doi.org/10.3390/geosciences10020067
Chicago/Turabian StyleBelov, Andrey A., Vladimir S. Cheptsov, Natalia A. Manucharova, and Zakhar S. Ezhelev. 2020. "Bacterial Communities of Novaya Zemlya Archipelago Ice and Permafrost" Geosciences 10, no. 2: 67. https://doi.org/10.3390/geosciences10020067
APA StyleBelov, A. A., Cheptsov, V. S., Manucharova, N. A., & Ezhelev, Z. S. (2020). Bacterial Communities of Novaya Zemlya Archipelago Ice and Permafrost. Geosciences, 10(2), 67. https://doi.org/10.3390/geosciences10020067