Spinal Reactive Oxygen Species and Oxidative Damage Mediate Chronic Pain in Lame Dairy Cows

{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioethics Statement
2.2. Animals
2.3. Lameness Assessment
2.4. Spinal Cord Processing, Protein Extraction, and Quantification
2.5. Laboratory Analysis
2.6. Statistical Analysis
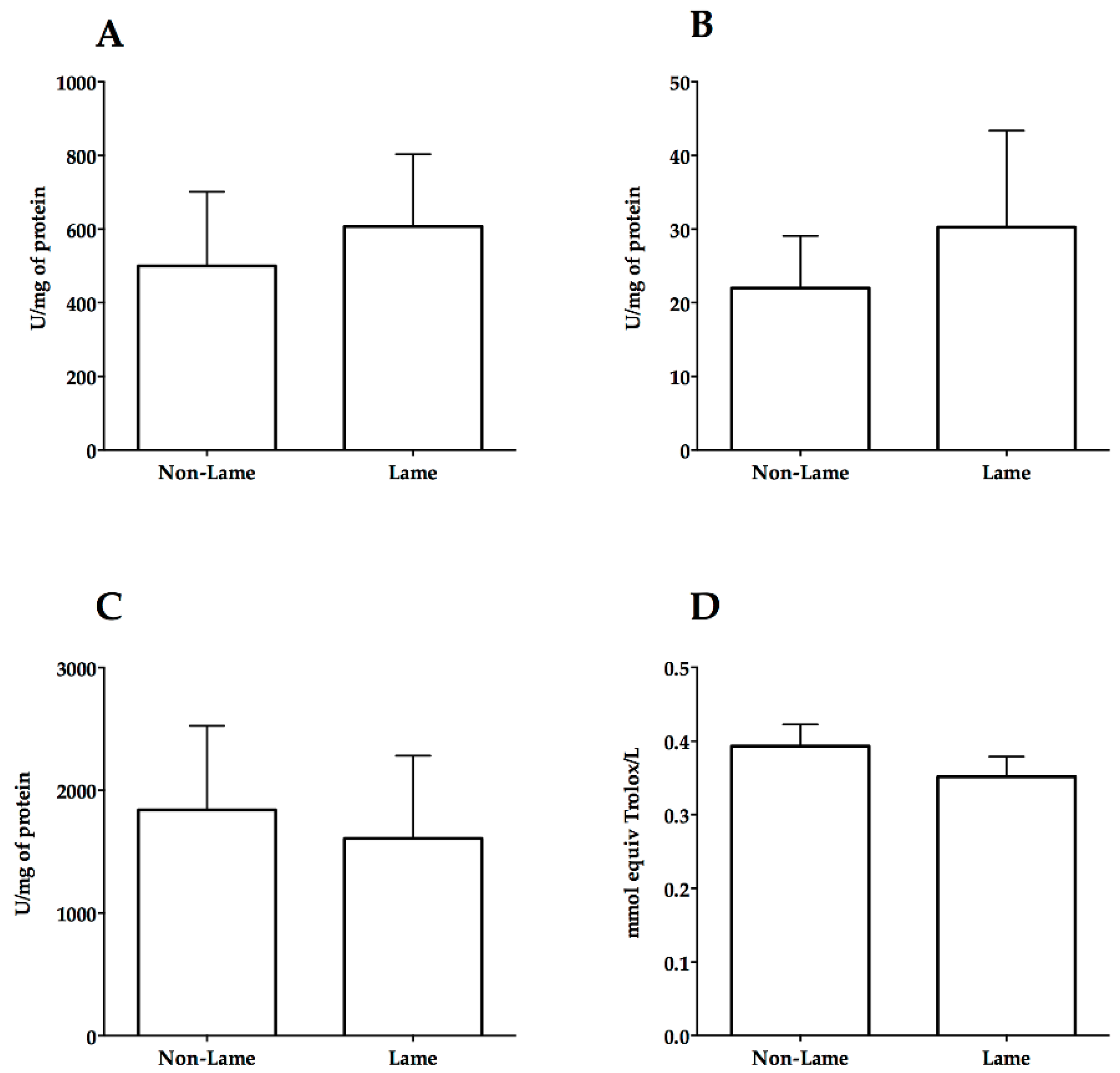
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Latremoliere, A.; Woolf, C.J. Central sensitization: A generator of pain hypersensitivity by central neural plasticity. J. Pain 2009, 10, 895–926. [Google Scholar] [CrossRef] [PubMed]
- Kuner, R. Spinal excitatory mechanisms of pathological pain. Pain 2015, 156, S11–S17. [Google Scholar] [CrossRef] [PubMed]
- Park, E.S.; Gao, X.; Chung, J.M.; Chung, K. Levels of mitochondrial reactive oxygen species increase in rat neuropathic spinal dorsal horn neurons. Neurosci. Lett. 2006, 391, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, D.; Little, J.W.; Doyle, T.; Neumann, W.L. Roles of reactive oxygen and nitrogen species in pain. Free. Radic. Biol. Med. 2011, 51, 951–966. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Kim, H.K.; Chung, J.M.; Chung, K. Reactive oxygen species (ROS) are involved in enhancement of NMDA-receptor phosphorylation in animal models of pain. Pain 2007, 131, 262–271. [Google Scholar] [CrossRef]
- Yowtak, J.; Lee, K.Y.; Kim, H.Y.; Wang, J.; Kim, H.K.; Chung, K.; Chung, J.M. Reactive oxygen species contribute to neuropathic pain by reducing spinal GABA release. Pain 2011, 152, 844–852. [Google Scholar] [CrossRef]
- Nishio, N.; Taniguchi, W.; Sugimura, Y.K.; Takiguchi, N.; Yamanaka, M.; Kiyoyuki, Y.; Yamada, H.; Miyazaki, N.; Yoshida, M.; Nakatsuka, T. Reactive oxygen species enhance excitatory synaptic transmission in rat spinal dorsal horn neurons by activating TRPA1 and TRPV1 channels. Neuroscience 2013, 247, 201–212. [Google Scholar] [CrossRef]
- Grace, P.M.; Gaudet, A.D.; Staikopoulos, V.; Maier, S.F.; Hutchinson, M.R.; Salvemini, D.; Watkins, L.R. Nitroxidative signaling mechanisms in pathological pain. Trends. Neurosci. 2016, 39, 862–879. [Google Scholar] [CrossRef]
- Haddad, J.J.; Land, S.C. Redox/ROS regulation of lipopolysaccharide-induced mitogen-activated protein kinase (MAPK) activation and MAPK-mediated TNF-alpha biosynthesis. Br. J. Pharm. 2012, 135, 520–536. [Google Scholar] [CrossRef]
- Kaur, U.; Banerjee, P.; Bir, A.; Sinha, M.; Biswas, A.; Chakrabarti, S. Reactive oxygen species, redox signaling and neuroinflammation in Alzheimer’s disease: The NF-κB connection. Curr. Top. Med. Chem. 2015, 15, 446–457. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Porreca, F.; Cuzzocrea, S.; Galen, K.; Lightfoot, R.; Masini, E.; Muscoli, C.; Mollace, V.; Ndengele, M.; Ischiropoulos, H.; et al. A newly identified role for superoxide in inflammatory pain. J. Pharm. Exp. Ther. 2004, 309, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Kim, H.K.; Kim, J.H.; Chung, K.; Chung, J.M. The role of reactive oxygen species in capsaicin-induced mechanical hyperalgesia and in the activities of dorsal horn neurons. Pain 2007, 133, 9–17. [Google Scholar] [CrossRef]
- Brieger, K.; Schiavone, S.; Miller, F.J., Jr.; Krause, K.H. Reactive oxygen species: From health to disease. Swiss Med. Wkly. 2012, 142, w13659. [Google Scholar] [CrossRef] [PubMed]
- Santo, A.; Zhu, H.; Robert Li, Y. Free radicals: From health to disease. React. Oxyg. Species 2016, 2, 245–263. [Google Scholar] [CrossRef]
- Klinck, M.P.; Mogil, J.S.; Moreau, M.; Lascelles, B.D.X.; Flecknell, P.A.; Poitte, T.; Troncy, E. Translational pain assessment: Could natural animal models be the missing link? Pain 2017, 158, 1633–1646. [Google Scholar] [CrossRef] [PubMed]
- Lascelles, B.D.X.; Brown, D.C.; Maixner, W.; Mogil, J.S. Spontaneous painful disease in companion animals can facilitate the development of chronic pain therapies for humans. Osteoarthr. Cartil. 2018, 26, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Lascelles, B.D.X.; Knazovicky, D.; Case, B.; Freire, M.; Innes, J.F.; Drew, A.C.; Gearing, D.P. A canine-specific anti-nerve growth factor antibody alleviates pain and improves mobility and function in dogs with degenerative joint disease-associated pain. BMC Vet. Res. 2015, 11, 101. [Google Scholar] [CrossRef] [PubMed]
- Gruen, M.E.; Thomson, A.E.; Griffith, E.H.; Paradise, H.; Gearing, D.P.; Lascelles, B.D.X. A feline-specific anti-nerve growth factor antibody improves mobility in cats with degenerative joint disease-associated pain: A pilot proof of concept study. J. Vet. Intern. Med. 2016, 30, 1138–1148. [Google Scholar] [CrossRef] [PubMed]
- Sadiq, B.M.; Ramanoon, S.Z.; Shaik Mossadeq, W.M.; Mansor, R.; Syed-Hussain, S.S. Association between lameness and indicators of dairy cow welfare based on locomotion scoring, body and hock condition, leg hygiene and lying behavior. Animals 2017, 7, 79. [Google Scholar] [CrossRef]
- Grandin, T. Welfare problems in cattle, pigs, and sheep that persist even though scientific research clearly shows how to prevent them. Animals 2018, 8, 124. [Google Scholar] [CrossRef]
- Green, L.E.; Hedges, V.J.; Schukken, Y.H.; Blowey, R.W.; Packington, A.J. The impact of clinical lameness on the milk yield of dairy cows. J. Dairy Sci. 2002, 85, 2250–2256. [Google Scholar] [CrossRef]
- Sogstad, A.M.; Østerås, O.; Fjeldaas, T. Bovine claw and limb disorders related to reproductive performance and production diseases. J. Dairy Sci. 2006, 89, 2519–2528. [Google Scholar] [CrossRef]
- Bustamante, H.A.; Rodríguez, A.R.; Herzberg, D.E.; Werner, M.P. Stress and pain response after oligofructose induced-lameness in dairy heifers. J. Vet. Sci. 2015, 16, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Reinecke, H.; Weber, C.; Lange, K.; Simon, M.; Stein, C.; Sorgatz, H. Analgesic efficacy of opioids in chronic pain: Recent meta-analyses. Br. J. Pharm. 2015, 172, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Wongrakpanich, S.; Wongrakpanich, A.; Melhado, K.; Rangaswami, J.A. Comprehensive review of non-steroidal anti-inflammatory drug use in the elderly. Aging Dis. 2018, 9, 143–150. [Google Scholar] [CrossRef]
- Fiore, E.; Perillo, G.; Piccione, G.; Giudice, E.; Zumbo, A.; Armato, L.; Fabbri, G.; Gianesella, M. Effect of claw horn lesions in Holstein dairy cows: Clinical and radiological study. Ann. Anim. Sci. 2019, 19, 147–158. [Google Scholar] [CrossRef]
- Novotna, I.; Langova, L.; Havlicek, Z. Risk factors and detection of lameness using infrared thermography in dairy cows – A review. Ann. Anim. Sci. 2019, 19, 563–578. [Google Scholar] [CrossRef]
- Rodriguez, A.R.; Olivares, F.J.; Descouvieres, P.T.; Werner, M.P.; Tadich, N.A.; Bustamante, H.A. Thermographic assessment of hoof temperature in dairy cows with different mobility scores. Livest. Sci. 2016, 184, 92–96. [Google Scholar] [CrossRef]
- Al-Qudah, K.M.; Ismail, Z.B. The relationship between serum biotin and oxidant/antioxidant activities in bovine lameness. Res. Vet. Sci. 2012, 92, 138–141. [Google Scholar] [CrossRef]
- Zhao, X.J.; Wang, X.Y.; Wang, J.H.; Wang, Z.Y.; Wang, L.; Wang, Z.H. Oxidative stress and imbalance of mineral metabolism contribute to lameness in dairy cows. Biol. Trace Elem. Res. 2015, 164, 43–49. [Google Scholar] [CrossRef]
- Reader, J.D.; Green, M.J.; Kaler, J.; Mason, S.A.; Green, L.E. Effect of mobility score on milk yield and activity in dairy cattle. J. Dairy Sci. 2011, 94, 5045–5052. [Google Scholar] [CrossRef]
- Gao, L.; Zhang, Z.; Xu, W.; Li, T.; Ying, G.; Qin, B.; Li, J.; Zheng, J.; Zhao, T.; Yan, F.; et al. Natrium benzoate alleviates neuronal apoptosis via the dj-1-related anti-oxidative stress pathway involving akt phosphorylation in a rat model of traumatic spinal cord injury. Front. Mol. Neurosci. 2019, 12, 42. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Mesquita, C.S.; Oliveira, R.; Bento, F.; Geraldo, D.; Rodrigues, J.V.; Marcos, J.C. Simplified 2,4-dinitrophenylhydrazine spectrophotometric assay for quantification of carbonyls in oxidized proteins. Anal. Biochem. 2014, 458, 69–71. [Google Scholar] [CrossRef]
- Erel, O. A novel automated method to measure total antioxidant response against potent free radical reactions. Clin. Biochem. 2004, 37, 112–119. [Google Scholar] [CrossRef]
- Hadwan, M.H.; Ali, S.K. New spectrophotometric assay for assessments of catalase activity in biological samples. Anal. Biochem. 2018, 542, 29–33. [Google Scholar] [CrossRef]
- Paglia, D.E.; Valentine, W.N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 1967, 70, 158–169. [Google Scholar]
- Schadrack, J.; Neto, F.L.; Ableitner, A.; Castro-Lopes, J.M.; Willoch, F.; Bartenstein, P.; Zieglgänsberger, W.; Tölle, T.R. Metabolic activity changes in the rat spinal cord during adjuvant monoarthritis. Neuroscience 1999, 94, 595–605. [Google Scholar] [CrossRef]
- Kunz, S.; Tegeder, I.; Coste, O.; Marian, C.; Pfenninger, A.; Corvey, C.; Karas, M.; Geisslinger, G.; Niederberger, E. Comparative proteomic analysis of the rat spinal cord in inflammatory and neuropathic pain models. Neurosci. Lett. 2005, 381, 289–293. [Google Scholar] [CrossRef]
- Komori, N.; Takemori, N.; Kim, H.K.; Singh, A.; Hwang, S.-H.; Foreman, R.D.; Chung, K.; Chung, J.M.; Matsumoto, H. Proteomics study of neuropathic and nonneuropathic dorsal root ganglia: Altered protein regulation following segmental spinal nerve ligation injury. Physiol. Genom. 2007, 29, 215–230. [Google Scholar] [CrossRef]
- Huang, H.L.; Cendan, C.M.; Roza, C.; Okuse, K.; Cramer, R.; Timms, J.F.; Wood, J.N. Proteomic profiling of neuromas reveals alterations in protein composition and local protein synthesis in hyper-excitable nerves. Mol. Pain 2008, 4, 33. [Google Scholar] [CrossRef]
- Gomez-Varela, D.; Barry, A.M.; Schmidt, M. Proteome-based systems biology in chronic pain. J. Proteom. 2019, 190, 1–11. [Google Scholar] [CrossRef]
- Cao, S.S.; Kaufman, R.J. Endoplasmic reticulum stress and oxidative stress in cell fate decision and human disease. Antioxid. Redox Signal. 2014, 21, 396–413. [Google Scholar] [CrossRef]
- Chakrabarti, A.; Chen, A.W.; Varner, J.D. A review of the mammalian unfolded protein response. Biotechnol. Bioeng. 2011, 108, 2777–2793. [Google Scholar] [CrossRef]
- Zeeshan, H.M.A.; Lee, G.H.; Kim, H.R.; Chae, H.J. Endoplasmic reticulum stress and associated ROS. Int. J. Mol. Sci. 2016, 17, 327. [Google Scholar] [CrossRef]
- Yang, E.S.; Bae, J.Y.; Kim, T.H.; Kim, Y.S.; Suk, K.; Bae, Y.C. Involvement of endoplasmic reticulum stress response in orofacial inflammatory pain. Exp. Neurobiol. 2014, 23, 372–380. [Google Scholar] [CrossRef]
- Zhang, E.; Yi, M.H.; Shin, N.; Baek, H.; Kim, S.; Kim, E.; Kwon, K.; Lee, S.; Kim, H.W.; Chul Bae, Y.; et al. Endoplasmic reticulum stress impairment in the spinal dorsal horn of a neuropathic pain model. Sci. Rep. 2015, 5, 11555. [Google Scholar] [CrossRef]
- Zhou, F.; Zhang, W.; Zhou, J.; Li, M.; Zhong, F.; Zhang, Y.; Liu, Y.; Wang, Y. Involvement of endoplasmic reticulum stress in formalin-induced pain is attenuated by 4-phenylbutyric acid. J. Pain Res. 2017, 10, 653–662. [Google Scholar] [CrossRef]
- Hermans, N.; Cos, P.; Maes, L.; De Bruyne, T.; Vanden Berghe, D.; Vlietinck, A.J.; Pieters, L. Challenges and pitfalls in antioxidant research. Curr. Med. Chem. 2007, 14, 417–430. [Google Scholar] [CrossRef]
- Wardman, P. Fluorescent and luminescent probes for measurement of oxidative and nitrosative species in cells and tissues: Progress, pitfalls, and prospects. Free Radic. Biol. Med. 2007, 43, 995–1022. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, W.; Zheng, B.; Tian, N. Sinomenine Attenuates traumatic spinal cord injury by suppressing oxidative stress and inflammation via Nrf2 pathway. Neurochem. Res. 2019, 44, 763–775. [Google Scholar] [CrossRef]
- Abuelo, A.; Gandy, J.C.; Neuder, L.; Brester, J.; Sordillo, L.M. Short communication: Markers of oxidant status and inflammation relative to the development of claw lesions associated with lameness in early lactation cows. J. Dairy Sci. 2016, 99, 5640–5648. [Google Scholar] [CrossRef]
- Ayala, A.; Muñoz, M.F.; Argüelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell. Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef]
- Komirishetty, P.; Areti, A.; Yerra, V.G.; Ruby, P.K.; Sharma, S.S.; Gogoi, R.; Sistla, R.; Kumar, A. PARP inhibition attenuates neuroinflammation and oxidative stress in chronic constriction injury induced peripheral neuropathy. Life Sci. 2016, 150, 50–60. [Google Scholar] [CrossRef]
- Hassler, S.N.; Johnson, K.M.; Hulsebosch, C.E. Reactive oxygen species and lipid peroxidation inhibitors reduce mechanical sensitivity in a chronic neuropathic pain model of spinal cord injury in rats. J. Neurochem. 2014, 131, 413–417. [Google Scholar] [CrossRef]
- Komirishetty, P.; Areti, A.; Gogoi, R.; Sistla, R.; Kumar, A. Combination strategy of PARP inhibitor with antioxidant prevent bioenergetic deficits and inflammatory changes in CCI-induced neuropathy. Neuropharmacology 2017, 113, 137–147. [Google Scholar] [CrossRef]
- Siems, W.; Bresgen, N.; Brenke, R.; Siems, R.; Kitzing, M.; Harting, H.; Eckl, P.M. Pain and mobility improvement and MDA plasma levels in degenerative osteoarthritis, low back pain, and rheumatoid arthritis after infrared A-irradiation. Acta Biochim. Pol. 2010, 57, 313–319. [Google Scholar] [CrossRef]
- Cheng, J.; Wang, F.; Yu, D.F.; Wu, P.-F.; Chen, J.G. The cytotoxic mechanism of malondialdehyde and protective effect of carnosine via protein cross-linking/mitochondrial dysfunction/reactive oxygen species/MAPK pathway in neurons. Eur. J. Pharm. 2011, 650, 184–194. [Google Scholar] [CrossRef]
- Thiele, G.M.; Duryee, M.J.; Anderson, D.R.; Klassen, L.W.; Mohring, S.M.; Young, K.A.; Benissan-Messan, D.; Sayles, H.; Dusad, A.; Hunter, C.D.; et al. Malondialdehyde-acetaldehyde adducts and anti-malondialdehyde-acetaldehyde antibodies in rheumatoid arthritis. Arthritis Rheumatol. 2015, 67, 645–655. [Google Scholar] [CrossRef]
- Fedorova, M.; Bollineni, R.C.; Hoffmann, R. Protein carbonylation as a major hallmark of oxidative damage: Update of analytical strategies. Mass Spectrom. Rev. 2014, 33, 79–97. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Giustarini, D.; Colombo, R.; Rossi, R.; Milzani, A. Protein carbonylation in human diseases. Trends Mol. Med. 2003, 9, 169–176. [Google Scholar] [CrossRef]
- Dalle-Donne, I.; Aldini, G.; Carini, M.; Colombo, R.; Rossi, R.; Milzani, A. Protein carbonylation, cellular dysfunction, and disease progression. J. Cell Mol. Med. 2006, 10, 389–406. [Google Scholar] [CrossRef]
- Petrov, D.; Zagrovic, B. Microscopic analysis of protein oxidative damage: Effect of carbonylation on structure, dynamics, and aggregability of villin headpiece. J. Am. Chem. Soc. 2011, 133, 7016–7024. [Google Scholar] [CrossRef]
- Curtis, J.M.; Hahn, W.S.; Long, E.K.; Burrill, J.S.; Arriaga, E.A.; Bernlohr, D.A. Protein carbonylation and metabolic control systems. Trends Endocrinol. Metab. 2012, 23, 399–406. [Google Scholar] [CrossRef]
- Noguchi, N. Role of oxidative stress in adaptive responses in special reference to atherogenesis. J. Clin. Biochem. Nutr. 2008, 43, 131–138. [Google Scholar] [CrossRef]
- Miller, Y.I.; Choi, S.-H.; Wiesner, P.; Fang, L.; Harkewicz, R.; Hartvigsen, K.; Boullier, A.; Gonen, A.; Diehl, C.J.; Que, X.; et al. Oxidation-specific epitopes are danger-associated molecular patterns recognized by pattern recognition receptors of innate immunity. Circ. Res. 2011, 108, 235–248. [Google Scholar] [CrossRef]
- Inanir, A.; Sogut, E.; Ayan, M.; Inanir, S. Evaluation of pain intensity and oxidative stress levels in atients with inflammatory and non-inflammatory back pain. Eur. J. Gen. Med. 2013, 10, 185–190. [Google Scholar] [CrossRef]
- Zheng, J.; Bizzozero, O.A. Accumulation of protein carbonyls within cerebellar astrocytes in murine experimental autoimmune encephalomyelitis. J. Neurosci. Res. 2010, 88, 3376–3385. [Google Scholar] [CrossRef]
- Papadopoulos, M.C.; Koumenis, I.L.; Yuan, T.Y.; Giffard, R.G. Increasing vulnerability of astrocytes to oxidative injury with age despite constant antioxidant defenses. Neuroscience 1998, 82, 915–925. [Google Scholar] [CrossRef]
- Gegelashvili, G.; Bjerrum, O.J. Glutamate transport system as a novel therapeutic target in chronic pain: Molecular mechanisms and pharmacology. Adv. Neurobiol. 2017, 16, 225–253. [Google Scholar]
- Viggiano, A.; Monda, M.; Viggiano, A.; Viggiano, D.; Viggiano, E.; Chiefari, M.; Aurilio, C.; De Luca, B. Trigeminal pain transmission requires reactive oxygen species production. Brain Res. 2005, 1050, 72–78. [Google Scholar] [CrossRef]
- Muscoli, C.; Cuzzocrea, S.; Riley, D.P.; Zweier, J.L.; Thiemermann, C.; Wang, Z.Q.; Salvemini, D. On the selectivity of superoxide dismutase mimetics and its importance in pharmacological studies. Br. J. Pharm. 2003, 140, 445–460. [Google Scholar] [CrossRef]
- Guedes, R.P.; Dal Bosco, L.; Araújo, A.S.; Belló-Klein, A.; Ribeiro, M.F.M.; Partata, W.A. Sciatic nerve transection increases gluthatione antioxidant system activity and neuronal nitric oxide synthase expression in the spinal cord. Brain Res. Bull. 2009, 80, 422–427. [Google Scholar] [CrossRef]
- Altindag, O.; Celik, H. Total antioxidant capacity and the severity of the pain in patients with fibromyalgia. Redox Rep. 2006, 11, 131–135. [Google Scholar] [CrossRef]
- Geyik, S.; Altunısık, E.; Neyal, A.M.; Taysi, S. Oxidative stress and DNA damage in patients with migraine. J. Headache Pain 2016, 17, 10. [Google Scholar] [CrossRef]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herzberg, D.; Strobel, P.; Chihuailaf, R.; Ramirez-Reveco, A.; Müller, H.; Werner, M.; Bustamante, H. Spinal Reactive Oxygen Species and Oxidative Damage Mediate Chronic Pain in Lame Dairy Cows. Animals 2019, 9, 693. https://doi.org/10.3390/ani9090693
Herzberg D, Strobel P, Chihuailaf R, Ramirez-Reveco A, Müller H, Werner M, Bustamante H. Spinal Reactive Oxygen Species and Oxidative Damage Mediate Chronic Pain in Lame Dairy Cows. Animals. 2019; 9(9):693. https://doi.org/10.3390/ani9090693
Chicago/Turabian StyleHerzberg, Daniel, Pablo Strobel, Ricardo Chihuailaf, Alfredo Ramirez-Reveco, Heine Müller, Marianne Werner, and Hedie Bustamante. 2019. "Spinal Reactive Oxygen Species and Oxidative Damage Mediate Chronic Pain in Lame Dairy Cows" Animals 9, no. 9: 693. https://doi.org/10.3390/ani9090693
APA StyleHerzberg, D., Strobel, P., Chihuailaf, R., Ramirez-Reveco, A., Müller, H., Werner, M., & Bustamante, H. (2019). Spinal Reactive Oxygen Species and Oxidative Damage Mediate Chronic Pain in Lame Dairy Cows. Animals, 9(9), 693. https://doi.org/10.3390/ani9090693